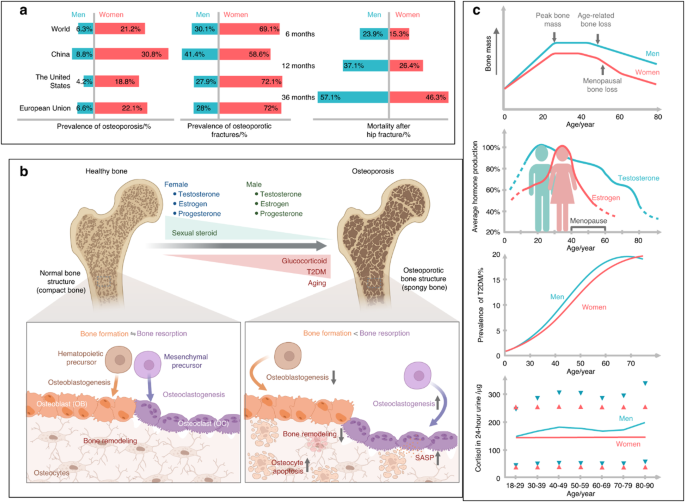
Current clinical evidence shows that pubertal suppression by treatment with GnRHa prevents the attainment of peak bone mass accrual in both trans boys and trans girls, which is only partially restored in trans girls following commencement of GAHT at a later age.3,4,7 It is unclear whether this deficit in peak bone mass in trans girls treated with GnRHa and GAHT persists until adulthood and whether it increases the risk of fracture. This is of particular importance, as an increase in fracture risk has been reported in two cohorts of trans women, compared to cis men, who did not undergo pubertal suppression but were treated with GAHT alone or in combination with gender-confirming surgery commencing in adulthood even after peak bone mass had been reached.23,24 Hence, it is possible that bone health in trans girls may be further compromised by current clinical approaches (e.g., GnRHa therapy), which impede the acquisition of peak bone mass. Importantly, these previous studies compared fracture risk in trans women compared to cis male controls but not to cis females. To address this important knowledge gap while overcoming the difficulties inherent in clinical trials in this population, we developed a mouse model of male-to-female transition in adolescence to investigate the effects of pubertal suppression and subsequent GAHT with estradiol on peak bone mass accrual, microstructure, and strength. Pubertal mice were orchidectomized at 5 weeks of age and administered estradiol at 3 weeks post-surgery for a period of 12 weeks to replicate pubertal suppression in early adolescence in trans girls with GAHT commencing sometime later. Skeletal integrity was assessed in adulthood in male-to-female mice at 20 weeks of age at a time when peak bone mass is expected to have been achieved (16 weeks in C57Bl/6 mice)22 and at an age that equates to 26-27 years in trans women.25 A strength of this study was the inclusion of multiple control groups, including both intact male and female controls, to allow comparisons between mice of the same birth sex as well as the desired gender following transitioning.
Puberty suppression by orchidectomy in pubertal male mice compared to male controls decreased peak bone mass accrual, with failure of cortical bone accumulation at both the periosteal and endosteal surfaces and trabecular bone in the metaphysis evident within the first 3 weeks post-surgery. The decrease in BV/TV in the metaphysis was associated with a reduction in the newly formed primary spongiosa arising from the growth plate characterized by a decrease in both the trabecular thickness and number, which is in accordance with trabecular bone accrual during puberty in males being dependent on androgen receptor (AR) activation in proliferating osteoblasts.26 The decrease in pubertal trabecular and cortical bone accrual in the metaphysis of Orx males was associated with bone of low mineral density, while the amount of mid-density bone accumulated prior to puberty was maintained. Oc.S/BS and N.Oc/B.Pm were decreased in Orx males at 12 weeks post-vehicle treatment compared to male controls, indicating that resorption was suppressed in trabecular bone. Consistent with this finding, the gene expression levels of Dcstamp, one of the key mediators of osteoclastogenesis, and Ctsk, a protease that degrades bone matrix, were decreased in whole bone of Orx males at 3 weeks post-surgery. In contrast, the expression of Acp5, a metalloenzyme in the acid phosphatase category, was unchanged. Similarly, mineral apposition, bone formation rates and expression of the osteoblast genes Col1a1 and Bglap were unaffected. The expression of all remodeling genes analyzed was low in all control groups at 12 weeks post-treatment (20 weeks of age), consistent with the age-related decline in bone remodeling.27 These data are consistent with previous reports of decreased cortical and trabecular bone acquisition in male rats and mice that were orchidectomized pre- or early puberty28,29 or administered high doses of GnRHa30 and assessed at 4 and 8 weeks post-surgery or 16 weeks post-GnRHa treatment, respectively. The results reported in the current and previous studies showing a decreased periosteal circumference and marked reduction in trabecular bone accrual following pubertal suppression by orchidectomy in male mice are consistent with AR signaling being the dominant pathway for periosteal and trabecular apposition of bone during puberty in male mice,20,29,31,32 while the decrease in cortical thickness is consistent with the increased endosteal erosion surface reported in prepubertal male mice Orx at 3 weeks of age.31
As expected, the failure to reach peak bone mass in Orx male-to-female mice at 20 weeks of age (12 weeks post-vehicle treatment) led to a reduction in bone strength as measured by pMOI and the maximal force the bone could withstand prior to fracture compared to intact male controls. Notably, however, although peak bone mass and bone strength were decreased in Orx male mice compared to male controls, there was no difference in any of the trabecular, cortical, mechanical or bone remodeling parameters compared to those of age-matched female controls at 20 weeks of age except for a reduction in trabecular BA/TA of the primary spongiosa. These data highlight the importance of careful consideration of control groups for clinical and preclinical studies of gender transition. While most clinical studies compare parameters between trans individuals and cis controls of the same birth sex, based on these data where the removal of endogenous sex steroids in Orx males led to female-like bones with the same strength as female controls, one might argue that comparisons between trans individuals and cis controls of the desired gender may be more appropriate, especially when assessing fracture risk.
The failure to reach peak bone mass in our early pubertal Orx mouse model compared to male but not female controls correlates well with longitudinal studies of transgender girls undergoing pubertal suppression with GnRHa therapy. In early pubertal trans girls defined by young bone age (<15 years)3 or Tanner stage 2 or 3,7 GnRHa treatment for a median of 1 or 2 years decreased BMAD Z scores of the lumbar spine, which was accompanied by a reduction in serum markers of bone remodeling. Similar observations were reported by Joseph et al. whereby GnRHa therapy in trans girls (average age of 13 years) resulted in a progressive decline in BMD and BMAD Z scores over 3 years, with the most rapid decrease occurring within the first year.33 It is important to note, however, that Z scores compare BMD to the average values of the same birth sex and age, while absolute BMD and BMAD of the femoral neck and lumber have been reported to be unchanged in each of these studies treating trans girls in early puberty with GnRHa for between 1 and 3 years, indicating an impairment to bone mass accrual rather than a loss of bone.3,7,33
To test whether GAHT in our male-to-female mouse model could restore bone accrual and strength in the pubertally suppressed male-to-female mice, we administered a high physiological dose of estradiol that was ∼2.7-fold higher than the peak estradiol levels that occur in the proestrus stage of the estrus cycle in female mice19 at 3 weeks post-orchidectomy for 12 weeks. As expected, serum concentrations of testosterone were decreased in Orx male-to-female mice, modeling serum concentrations observed in trans girls on GAHT.16 Estradiol treatment in male-to-female mice compared to all control mice increased the cortical thickness at the mid-diaphysis, consistent with the actions of high-dose estradiol to stimulate the endocortical deposition of bone previously reported in young ovariectomized female mice.34 Of interest, a bone size similar to that observed at 3 weeks post-orchidectomy was maintained with estradiol treatment of Orx male-to-female mice for 12 weeks, such that the periosteal circumference was at an intermediate level between male sham and Orx controls. These data are consistent with the hypothesis proposed by Venken et al. 29 that signaling via both the AR and estrogen receptor (ER) is needed for the peak stimulation of periosteal growth during puberty in male mice. The stimulatory effects of estradiol on periosteal expansion have also been reported in a case study of a 17-year-old boy with aromatase deficiency in whom treatment with estradiol for 3 years partially restored the bone size to levels observed in control males.35
Since disorders of overly dense bone, such as osteopetrosis, can lead to increased fragility, 3-point bending analyses were performed on the femur to determine the effect of the marked increase in cortical bone accrual at the mid-diaphysis following estradiol treatment in the male-to-female mice. Three-point bending analysis confirmed that the larger cross-sectional area and thicker cortices at the mid-diaphysis of the estradiol-treated Orx male-to-female mice made the bone stiffer and stronger and able to withstand a higher maximal force compared to all control groups. In addition, bone stiffness was higher while postyield displacement was lower in estradiol-treated Orx male-to-female mice compared to male Orx controls, which may be attributed to a decrease in the ratio of collagen to bone mineral content.36 Bending stress could be measured in future experiments to account for the increased cross-sectional area; however, this was not possible in the current study due to technical limitations. We also acknowledge that it remains theoretically possible that bone strength might be compromised at other sites in estradiol-treated Orx male-to-female mice, a possibility to be addressed by future work.
Estradiol treatment in pubertal Orx male-to-female mice markedly increased all the mineral density ranges (low, mid, upper) of trabecular and cortical bone in the distal femoral metaphysis compared to all control groups. The 18-fold increase in trabecular bone accrual was primarily driven by a concomitant 14-fold increase in trabecular number associated with an increase in the number of trabeculae arising from the growth plate in the primary spongiosa compared to Orx male controls. This was accompanied by an increase in the number of mineralizing surfaces and the bone formation rate within the secondary spongiosa of the metaphysis, while the mineral apposition rate was unaffected. The anabolic effect of estradiol treatment in male-to-female mice was remarkable given the well-established ability of estradiol to inhibit bone resorption; however, these findings are consistent with those of previous reports that estradiol increases osteoblast proliferation and inhibits apoptosis in vitro.37,38,39 Similarly, high doses of estradiol (500 μg once per week for 4 weeks) in young female ovariectomized mice compared to intact female and ovariectomized female controls have been shown to increase BV/TV by increasing the number of bone forming surfaces in vivo.34 The anabolic action of estradiol to increase bone accrual in Orx male-to-female mice was accompanied by an increase in the markers of bone resorption (Oc.S/BS, N.Oc/B.Pm and Dc-stamp, Acp5 and Ctsk gene expression). Considering the well-documented antiresorptive and proapoptotic actions of estradiol on osteoclasts, the increase in the histomorphometric and gene expression measures of bone resorption in the presence of high concentrations of estradiol in Orx male-to-female mice was intriguing. We speculate that the actions of estradiol to increase osteoblast proliferation and bone mass in these mice also resulted in a concomitant increase in the secretion of coupling factors by osteoblasts, which increased osteoclastogenesis. While this would be expected to decrease bone mass, it is plausible that the high concentration of estradiol suppressed the resorptive capacity of the osteoclasts, resulting in an uncoupling of bone remodeling with bone formation exceeding bone resorption and leading to increased bone mineral accrual. A similar phenotype is observed in patients with osteoclast-rich osteopetrosis, whereby bone formation is normal or increased in the presence of normal or increased numbers of multinucleated osteoclasts with impaired ruffled-border formation and resorption capacity, thus leading to increased bone formation in the presence of increased markers of resorption.40 Confirmation of this hypothesis would require further in-depth investigation including measures of osteoclast function (i.e., actin ring and ruffled border formation) together with an extensive analysis of the coupling process between bone resorption and formation, which is a multifaceted process involving numerous regulators.41 The mechanism of estradiol action to increase trabecular bone accrual in the current study by stimulating the formation of primary spongiosa in addition to increasing bone formation in the secondary spongiosa suggests that in trans girls, GAHT should be started as soon as possible after pubertal suppression to maximize the formation of new trabeculae from the growth plate before its fusion upon withdrawal of GnRHa treatment.
While there are very few studies investigating the effects of estradiol treatment in male Orx mice either prepubertally or in early puberty, a dose-dependent increase in the tibial bone ash weight and the levels of calcium and phosphorus as well as an increased metaphyseal and epiphyseal bone area were previously reported in male Orx mice at 3 weeks of age treated with increasing doses of estradiol 1 week post-surgery for 4 weeks.42 No effect of estradiol treatment was observed on these parameters in intact male mice. The authors suggest a possible antagonistic effect of testosterone on estradiol actions within bone such that intact males do not respond to estradiol, but in the absence of androgens, estradiol can act on growing male bones in a similar way to that of females.42 The mechanism of the anabolic action of estradiol to strikingly increase trabecular bone in the Orx male-to-female mice is unclear but is likely to be mediated, at least in part, by insulin-like growth factor-1 (IGF-1). Previously, estradiol treatment of adult (12-month-old) male mice at the time of orchidectomy prevented Orx- and age-related loss of the trabecular bone density, which was associated with increased circulating IGF-I concentrations.43 It was not possible to measure serum IGF-1 concentrations in the current study due to an insufficient volume of serum remaining following sex steroid analyses, and this warrants further investigation.
Although there was no measurable effect of sex, orchidectomy or estradiol treatment on the femoral length, male controls had a decreased growth plate thickness compared to female controls, consistent with the sexual dimorphism of bone. Orchidectomy in pubertal males increased the growth plate height, while estradiol treatment to Orx male-to-females restored the growth plate height to a level that was intermediate between male and female controls 12 weeks post-treatment. Evidence from cartilage-specific ERα knockout mice suggests that the mechanism of action for the decrease in growth plate height following estradiol treatment in Orx male-to-female mice is likely to be direct via ERα to decrease chondrocyte proliferation in adulthood.44
The marked increase in cortical and trabecular bone accrual following estradiol treatment in the Orx male-to-female mice in the current study contrasts with several reports in adolescent trans girls in whom GAHT was unable to fully restore the BMD and peak bone mass following puberty suppression. Although GAHT following GnRHa treatment in trans girls has been shown to increase the BMAD at both the lumbar spine and femoral neck and the aBMD at the lumbar spine, the BMAD Z scores did not recover to pre-GnRHa treatment scores and remained below zero,3,4,7 while levels of serum markers of bone remodeling were decreased.3,7 These differences may be ascribed to the dosing regimens of estradiol used, with a recent study suggesting insufficient serum estradiol concentrations as a possible reason for failure of the BMD Z score catch-up despite long-term GAHT.8 In contrast to our dosing regimen of GAHT in the current preclinical study, whereby delivery of estradiol via a silastic implant delivered a sustained high concentration of estradiol, often in trans girls, estradiol is administered in gradually increasing doses every 6 months for the first two years of treatment until a maximum of 2 mg/d and a sustained physiological concentration of 100–200 pg/mL serum estradiol is achieved,45,46 although exact clinical treatment regimens vary among experts. The marked increase in bone accrual and strength observed in the male-to-female mice in the setting of high serum concentrations of estradiol in the current study suggests that higher doses of estradiol may be needed to restore peak bone mass following puberty suppression in trans girls. Due to the lack of dose response experiments, we cannot directly confirm this hypothesis. Given the complexity of such experiments, we instead chose a single estradiol dose based on preliminary dose-finding experiments. However, our inference that a higher dose of estradiol might overcome bone deficits in trans girls is supported by a recent study by Boogers et al.,16 whereby trans girls on GnRHa with very tall height prediction were treated with a higher than standard dose of estradiol (6 mg/d vs. 2 mg/d). After a median of 2.8 years after treatment with estradiol, trans girls treated with the higher dose had a median serum estradiol concentration of 522 pg/mL and a greater increase in height-adjusted BMD Z scores (HAZ-scores) in both the lumbar spine and femoral neck compared to those treated with the standard dose and a median serum estradiol concentration of 126 pg/mL. The HAZ scores in the trans girls treated with the higher dose returned to the pretreatment HAZ scores after two years of GAHT, while those in trans girls receiving the regular dose of estradiol did not. Whether this increase in BMD with the higher estradiol dose is maintained once the patient has reached their expected height and begins receiving the regular dose is not known. Overall, consistent with our rodent model of adolescent male-to-female transition, the existing evidence suggests that skeletal integrity can be preserved if a dose of estradiol is administered that is sufficiently high. However, whether similar doses to those administered in our rodent models can be administered safely to trans girls or what the optimal dose of estradiol would be in trans girls is an important clinical question that requires further study. In this context, potential risks of higher-dose estradiol, e.g., thromboembolic complications, must be taken into consideration; of note, while no overt adverse events were noted with regular monitoring of the mice as per ethics approval, our study was not designed to assess risks of higher-dose estradiol, especially in humans.
The marked increase in cortical and trabecular bone accrual following estradiol treatment in the male-to-female mice suggests that the intraskeletal concentrations of estradiol were increased by the high dose of estradiol administered. In the setting of low serum testosterone concentrations and high serum estradiol concentrations, these data infer that an increase in the intraskeletal concentration of estradiol arose via diffusion into bone from the circulation rather than from the aromatization of testosterone by osteoblasts within bone. Confirming this notion requires the measurement of the intraskeletal concentrations of estradiol. Despite our efforts, we have yet to validate an LCMS method for measuring intraskeletal concentrations of sex steroids, which remains a current line of investigation.
Additional limitations of this study not previously discussed may include the use of orchidectomy rather than GnRHa therapy to reduce sex steroids to castration concentrations, although it is theoretically possible that due to their differential effects on gonadotrophins, these treatments may not have the same bone effects.47 The skeletal deficits observed in Orx male mice are broadly consistent with those reported for puberty-suppressed female adolescents and GnRHa-treated male mice.3,7,30,33 The marked increase in bone mass, which led to a visible reduction in marrow space, may have adversely affected blood cell production, resulting in anemia. The limited blood volume available for analysis prohibited these measurements. It is also possible that the increased bone mass and decreased marrow space in estradiol-treated male-to-female mice compared to control mice altered the cell populations within bone, thereby potentially confounding the gene expression data. Finally, the small number of samples for the histomorphometry analyses in some of the groups likely decreased the power to detect significant differences. Nevertheless, it is encouraging that the findings from the histomorphometric and gene expression analyses were congruent and aligned with the effect of estradiol treatment on bone structure determined by μCT in the male-to-female mice.
In conclusion, we have shown that the failure to achieve peak trabecular and cortical bone mass accrual in male-to-female mice and the deficit in bone strength compared to control males following pubertal suppression can be rectified by administration of a sustained, high dose of estradiol in late puberty. In fact, the high physiological concentrations of circulating estradiol achieved in the male-to-female mice in the current study were anabolic for bone, markedly increasing trabecular and cortical bone deposition, which translated to an increase in bone strength compared to both male and female controls. The increased bone accretion in the setting of low serum concentrations of testosterone in the male-to-female mice treated with estradiol suggests that the high serum concentrations of estradiol circumvented the requirement for local conversion of testosterone to estradiol by aromatase within bone. For these data to be informative for the preservation of skeletal integrity and strength of trans girls on GnRHa and GAHT, further study is needed comparing lower doses of estradiol to achieve circulating concentrations closer to the mid- and upper ranges of the proestrus cycle in female mice to better reflect the standard and high doses of GAHT prescribed for trans girls.16 Importantly, for clinical relevance, these future studies will need to address the possible adverse side effects of estradiol treatment (i.e., long-term cardiovascular risk and venous thromboembolism48). Nonetheless, these data are consistent with our hypothesis and imply that it is possible to preserve bone health and prevent long-term fractures in trans girls treated with GnRHa and GAHT if a dose of estradiol that protects the bone without adverse side effects can be identified. Given the lack of information assessing either bone microstructure by high-resolution modalities such as HR-pQCT or fracture prevalence following pubertal suppression and GAHT in trans girls, future studies in this area should be considered a research priority.


留言 (0)