
記住我



Alterations in DNA damage response (DDR) and related genes constitute one of the molecular hallmarks of prostate cancer (PCa).1–3 Up to 25% of metastatic PCa (mPCa) harbor DDR alterations, most frequently involving the homologous recombination repair (HRR), the Fanconi anemia (FA), and the mismatch repair (MMR) pathways1,2,4–6 (Fig. 1). The most frequently altered genes in mPCa are BRCA2 (9% to 13%), ATM (5% to 7%), CDK12 (≈5%), Fanconi Anemia Complementation Group A (FANCA) (1% to 5%), MSH2 (≈ 2%), BRCA1 (1% to 2%), and checkpoint kinase 2 (CHEK2) (1% to 2%),7–12 and ≈8% of these alterations are germline.8 The HRR pathway plays an essential role in the repair of DNA double-strand breaks (DSBs), along with the error-prone nonhomologous end-joining (NHEJ) pathway. While the HRR pathway is mainly active in the S and G2 cell cycle phases, the NHEJ is active during all phases of the cell cycle.4,13,14 DNA interstrand crosslinks (ICLs) and replication fork stalling are resolved by the FA pathway in cooperation with nucleotide excision repair, HRR, and translesion synthesis, while nucleotide mismatches are resolved by the MMR pathway.15,16 Decomposition by mutational signatures of PCa tumor samples showed the relevant contribution of several DDR-related single-base substitutions (SBSs) and indel (ID) mutational signatures.3,12,17 In previous work, we analyzed the prevalence of alterations in DDR genes in a large PCa brain metastases cohort and could correlate the presence of specific DDR-related SBS signatures (SBS44) with underlying genomic alterations (MSH2 defects).12 In the same cohort, a high representation (>10% of mutations) of the HRR defective SBS3 signature was identified.12 From a therapeutic perspective, relevant efforts have been made to target DDR alterations in mPCa. However, the most relevant clinical benefit from poly (ADP-ribose) polymerase inhibitors (PARPis) has been demonstrated for BRCA1- and BRCA2-alterations.18–22 PARPis are synthetically lethal with BRCA1/2 deficiencies, leading to the accumulation of DNA single-strand breaks in cells with pre-existing deficient DSB repair. For many other DDR alterations, including ATM, responses to PARPi are less prominent and much more heterogeneous, underlining their distinct impact on the DNA repair phenotype. Moreover, for BRCA2- and PALB2-deficiencies, higher efficacy of PARPis has been observed for PCa tumors harboring biallelic alterations.19 Further, PCa tumors deficient for mismatch proteins [mismatch repair–deficient (dMMR)] or with microsatellite instability (MSI-high), and a subset of tumors with CDK12 deficiency, are vulnerable to treatment with immune checkpoint inhibitors (ICIs). We aimed to review the most common non-BRCA DDR genomic alterations in mPCa and analyze their current and emerging clinical implications.
 FIGURE 1:
FIGURE 1: DNA repair pathways most frequently altered in prostate cancer. Most frequent DDR genomic alterations in prostate cancer concern genes involved in the homologous recombination repair pathway (BRCA2, BRCA1, ATM, PALB2, CHEK2), in the Fanconi Anemia pathway (FANCA), and the mismatch repair pathway (MSH2, MSH6). Schematic representation of the (A) HRR and FA pathways and (B) NHEJ and MMR pathways. DDR indicates DNA damage response; FA, Fanconi anemia; FANCA, Fanconi Anemia Complementation Group A; HRR, homologous recombination repair; MMR, mismatch repair; NHEJ, nonhomologous end joining.
GERMLINE DNA DAMAGE REPAIR ALTERATIONS IN PROSTATE CANCERMost frequent germline alterations in PCa concern DDR genes, including MMR genes. Previous studies identified the presence of germline alterations in around 8% of PCa patients, most frequently occurring in BRCA2 (≅5%), ATM (≅1%), and BRCA1 (<1%).23 Further studies showed even higher frequencies of genomic alterations, identifying up to 17.2% of included patients (BRCA2, 4.7%; CHEK2, 2.9%; MUTYH, 2.4%; and ATM, 2.0%).24 On the other hand, germline variants are most frequently found for PCa harboring genomic alterations in PALB2, CHEK2, BRCA1, BRCA2, and ATM.25 For instance, a germline ATM alteration is related to a 4-fold increase in PCa risk.26 Moreover, higher variant allele frequencies in next-generation sequencing results increase the probability of an underlying germline alteration.25HOXB13 is another gene in which germline alterations lead to an increased risk of PCa and are related to familial PCa.27 In the Cancer Genome Atlas cohort, which included samples from localized PCa, the frequency of germline alterations was lower (4.6%).10
Following current NCCN and ESMO guidelines, patients with tumors with pathogenic or likely pathogenic mutations in BRCA1, BRCA2, ATM, PALB2, and CHEK2, as well as Lynch syndrome-associated genes, should undergo germline counseling and/or testing.28,29
MOLECULAR LANDSCAPE OF NON-BRCA DNA DAMAGE RESPONSE ALTERATIONS IN PROSTATE CANCER Homologous Recombination Repair and Nonhomologous End-joining Pathways Ataxia Telangiectasia MutatedAtaxia telangiectasia mutated (ATM) is a protein kinase crucial in DSB signaling, whose activation leads to the amplification of the DNA damage signal resulting in stimulation of the HRR and the NHEJ pathways.30 ATM activation is induced by DNA DSBs and the resulting recruitment of the MRN complex (MRE11, RAD50, and NBS1) to the site of the damage promotes HRR. However, other DNA lesions, such as single-strand break, topoisomerase I cleavage complexes,31 and complex cellular events, such as oxidative stress32 eventually lead to ATM activation. When activated, the ATM homodimer undergoes monomerization, promoting the initial stages of DNA resection through the CtBP-interacting protein (CtIP) in collaboration with the MRN complex. Along the HRR pathway, ATM interacts with multiple other key components, including EXO1 and BRCA1, and PALB2. Besides HRR, ATM is also activated in the context of the NHEJ pathway. Upon recognition of the DNA DSBs by the Ku heterodimer (Ku70-Ku80), DNA pyruvate kinase catalytic subunits (DNA PKcs) are recruited to the DNA damage foci. These DNA PKcs are phosphorylated by ATM, stimulating the DNA repair process. On the other side, DNA PKcs also phosphorylate ATM, repressing its interaction with the MRN complex.33 ATM also regulates the TP53 binding protein 1 (53BP1), which governs the DNA end resection and promotes NHEJ in favor of the HRR pathway.33,34 Preclinical studies have shown that NHEJ-dependent DSB repair is impaired in ATM-deficient cells.35 Moreover, further preclinical studies have demonstrated that RAD51 foci formation is not impaired in irradiated ATM knock-out (ATMKO) PCa cell lines, indicating a lower impact of ATM deficiency on the HRR function.36 This finding was also confirmed when HRR function was assessed by DR-GFP assay.36 ATM is altered in ≈5% to 7% of PCa and is enriched in high Gleason tumors.37 Drug sensitivity studies in ATMKO PCa cell lines showed increased sensitivity to ATR inhibitors compared to PARP inhibitors.36 ATR inhibitors block phosphorylation of CHK1, leading to cell cycle arrest in S and G2/M.38 This synthetic lethality with ATR inhibitors has also been observed in other neoplastic cell lines with ATM deficiency, such as chronic lymphocytic leukemia cell lines.39 In the clinical scenario, several recent trials showed a somewhat limited benefit for PARP inhibitors in patients with ATM-deficient metastatic castration-resistant PCa (mCRPC) compared to patients with BRCA-deficient tumors.20,40,41
Partner and Localized of BRCA2PALB2 (Partner and Localized of BRCA2) is an essential component of the HRR pathway.42 Following direct interaction with BRCA1, PALB2 recruits BRCA2 and RAD51 monomers to the sites of DNA DSBs and supports strand invasion within the HRR pathway.42 Moreover, PALB2 plays a relevant role in maintaining genomic stability under exposure to DNA-damaging agents.43 In the biomarker analysis performed within the TOPARP-B trial patient population, patients with biallelic loss of PALB2 received benefit from treatment with olaparib, along with patients harboring homozygous BRCA2 alterations.19 In contrast, monoallelic alterations did not derive major benefits.19 A recent study in a Polish PCa population showed that PALB2 alterations were associated with an age-adjusted hazard ratio for mortality of 2.52 (P=0.0023), pointing to a derived more aggressive tumor phenotype.44 Moreover, biallelic alterations in PALB2 (also termed FANCN) have been reported to cause a severe subtype of FA disease, leading to increased cancer predisposition (eg, acute myeloid leukemia or neuroblastoma) in childhood.45 In melanoma, the presence of mutations in PALB2 has been correlated with higher tumor mutational burden.46
Checkpoint Kinase 2CHEK2 encodes the serine/threonine kinase CHK2 and is altered in about 1% to 2% of PCa.11,47 The occurrence of DNA DSB promotes CHK2 phosphorylation through ATM, which leads to CHK2 dimerization and autophosphorylation. Following activation, CHK2 phosphorylates multiple nuclear proteins involved in DNA repair, such as BRCA1 and BRCA2, promoting the HRR pathway.48 CHK2 also phosphorylates p53 and other proteins involved in the cell cycle and apoptosis regulation.47 Following DNA DSB damage, CHK2 promotes cell cycle arrest in G1/S and G2/M. Also, phosphorylation of p53 promotes cell cycle arrest in the G1/S phases.49,50 Specific alterations in CHEK2 (1100delC and I157T mutations) have been correlated with an increased risk of PCa. However, no association with familiar PCa has been shown.51
Fanconi Anemia Pathway Fanconi Anemia Complementation Group AThe FA pathway is essential for repairing DNA ICLs, which leads to DNA replication fork stalling.15 Among the multiple DNA repair proteins involved in the FA pathway, Fanconi Anemia Complementation Group A (FANCA) is the most commonly altered in PCa (2.5% to 3%), including deep deletions in ≈2.5% tumors and mutations in ≈ 0.5%.1,52 Besides being a key component of the FA core complex, FANCA is also involved in other DDR pathways, such as the single-strand annealing pathway, contributing to the DNA DSB repair.15,53 Once established, ICLs are identified by the FANCM-FAAP24-MHF1-MHF2 complex, which then recruits the rest of the components of the FA core complex.15 In this pathway step, an interaction with ATR and BRCA1 is required to recruit the FA core complex successfully. The activation of the FA core complex enables the monoubiquitylation of the paralogue FANCD2 and FANCI heterodimers, which promotes the nucleotide excision by ERCC4-ERCC1, required to release one of the DNA strands at the ICL (unhooking).15,54 The missing nucleotides are inserted and extended by a DNA polymerase within translesion synthesis. The insertion step generates many mutations at the initial ICL location. Once this translesion synthesis is completed, incisions leading to DNA DSBs are generated at the initial ICL location. The BRCA2-PALB2 complex plays, in combination with RAD51, an essential role in the repair of these DSB breaks through the HRR pathway. Moreover, an interaction of FANCJ or BRIP1 with the MMR proteins MLH1 and PMS2, which constitute the hMutLa heterodimer, is also required for the correct ICL repair.55 Interestingly, the FA pathway downregulates the activity of the NHEJ pathway but promotes the alternative end-joining pathway.15 In recent work, molecular characterization of squamous cell carcinomas from patients with FA showed that deficiencies in the FA function lead to a high prevalence of structural variants and complex genomic rearrangements.56
Cyclin-dependent Kinase 12Cyclin-dependent kinase 12 (CDK12) alterations are found in ~5% of PCa.57–59 Loss-of-function alterations of CDK12 impairs DNA repair through modulating expression levels of several DNA repair genes. By suppressing polyadenylation, CDK12 supports the production of full-length transcripts, a process relevant to many DNA repair genes. Therefore, the loss of CDK12 leads to impaired expression of several DDR genes.60,61 This effect is gene-length dependent, leading to transcription termination at 3′ and polyadenylation, which leads to earlier cleavage of long genes (>45 kb). Moreover, DDR genes, such as BRCA1, BARD1, or RAD51, are enriched in polyadenylation sites, which makes them especially vulnerable to the absence of CDK12. In PCa, biallelic CDK12 deficiency constitutes a distinct and unique molecular subtype of PCa, with mutual exclusivity with SPOP mutations and ETS fusions.57CDK12-deficient PCa is characterized on the genomic level by focal tandem duplications and high neoantigen burden, making these tumors vulnerable to ICIs. This increased neoantigen burden is associated with higher immune infiltration, with enrichment in CD4+ FOXP3- T regulatory cells.57,62 Moreover, CDK12-deficient tumors lack genomic signatures characteristic of HRD tumors. These tumors are typically characterized by poor prognosis, show poor responses to androgen receptor (AR) signaling inhibition (ARSI), PARPi, and taxane-based chemotherapy, and exhibit variable responses to ICIs.62,63
Mismatch Repair Pathway3% to 4% of PCa harbor alterations in the MMR genes. Mismatch DNA lesions are recognized by the hMutSa heterodimer (MSH2 and MSH6), which preferentially identifies single-base mismatches, or by the hMutSa (MSH2 and MSH3) complex, which recognizes mismatches originating through small insertions or deletions.16 Moreover, the hMutLa heterodimer (MLH1 and PMS2) is recruited to the DNA lesion. This heterodimer has endonuclease activity and is required to support the nucleotide excision by EXOI 3′→5′, which can independently resect in the direction 5′→3′). Proliferating cellular nuclear antigen (PCNA) interacts with both heterodimers supporting the initiation of DNA synthesis, which is performed by the DNA polymerase δ. In mCRPC, the most common MMR alterations are found in MSH2 and MSH6.8,10,11 On the IHC level, the loss of MSH2 usually co-occurs with MSH6, either due to germline or biallelic somatic mutations in MSH2, MSH6, or EPCAM. However, the loss of MSH6 may present independently of conserved MSH2 IHC. We summarized the main available methods and assays able to assess DDR and MMR defects in Table 1.
TABLE 1 - Summary of Testing Platforms for DDR and MMR Alterations Methodology Platform/assay Scope and thresholds DDR tumor testing NGS(targeted assays) Myriad Genetics MyChoice CDx Mutations and large rearrangements in 15 DDR genes: ATM, BARD1, BRCA1, BRCA2, BRIP1, CDK12, CHEK1, CHEK2, FANCL, PALB2, PPP2R2A, RAD51B, RAD51C, RAD51D, RAD54LCNV indicates copy number variations; IHC, immunohistochemistry; LOH, loss of heterogeneity; LST, large-scale transitions, MMR, mismatch repair; MSI, microsatellite instability; NGS, next-generation sequencing; PCR, polymerase chain reaction; SNV, single nucleotide variants; TAI, telomeric allelic imbalance; TMB, tumor mutational burden.
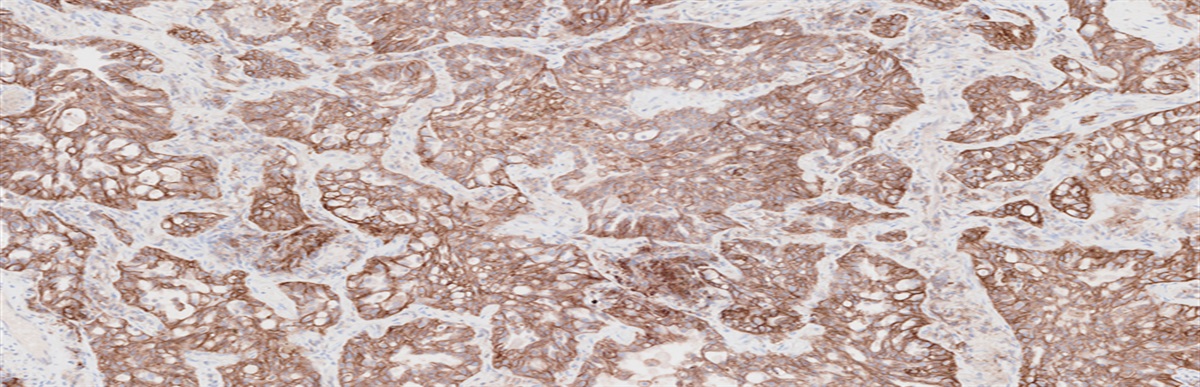
PCa tumors harboring BRCA2 defects, especially biallelic alterations, have been correlated with higher Gleason scores and more aggressive histologic subtypes, such as cribriform histology, as well as with the presence of intraductal carcinoma.64–68 However, an association with germline BRCA2 alterations could not be demonstrated.68 On the other hand, an especially high (49%) prevalence of DNA repair alterations has been reported in a cohort of ductal PCas (14% of patients had an alteration within the MMR genes and 31% within the HRR pathway).69 Relevantly, 20% of the patients had an underlying DDR germline autosomal dominant mutation. Moreover, intraductal histology has been correlated with higher genomic instability scores.70 A retrospective analysis of a PCa patient cohort (n=60) with at least one monoallelic alteration in CDK12 showed a very high prevalence of high Gleason scores (93.3%) and the presence of intraductal histology in 15.4% of the patients.62,63 Similarly, dMMR/MSI-H PCas usually present as undifferentiated tumors (grade group 5) and are also frequently associated with intraductal histology.71 However, the recommendation to perform germline testing based on the presence of intraductal or cribriform histology variant is highly controversial, and only recommended as optional by the current guidelines.28
PRECISION ONCOLOGY TARGETING OF NON-BRCA DNA DAMAGE REPAIR ALTERATIONS IN PROSTATE CANCER Established Therapeutic Strategies Chemotherapy Agents, Androgen Signaling Inhibition, and Radiotherapy“Classical” DNA-targeting drugs, such as topoisomerase II inhibitors (mitoxantrone) or DNA cross-linking agents (carboplatin), have been commonly used in the treatment of mCRPC. For platinum-based chemotherapy, studies including PCa molecular characterization have confirmed the enhanced activity of platinum-derivates in tumors with DDR alterations.72–75 Several studies have shown that AR signaling upregulates the expression of DDR proteins so that a combination of radiotherapy with androgen deprivation therapy (ADT) is considered synergistic.76 In fact, radiotherapy treatment for localized PCa is usually combined with ADT±ARSI.77,78 However, PCa tumors with BRCA1/2 alterations classically show shorter responses to ARSI.79
Moreover, several studies have analyzed how the presence of specific DDR proteins correlates with the efficacy of radiotherapy treatment in PCa. For instance, high ATM expression in PCa tumor tissue was correlated with worse clinical outcomes in patients with localized PCa treated with radiotherapy.80 In addition, 2 metastases-directed radiotherapy studies for oligometastatic disease (ORIOLE and STOMP) pointed to a larger benefit from radiotherapy treatment in patients with alteration in ATM, BRCA1/2, Rb1, or TP53.81
Poly-ADP-ribose Polymerase InhibitorsEstablished targeted treatment options for mPCa with DDR alterations include PARPi and immunotherapy with ICIs. Across distinct clinical trials, treatment with PARPi has shown major clinical efficacy in BRCA1/2-mutated PCa.19–22,40,41 The PROfound phase 3 trial showed overall survival (OS) benefit for the cohort of patients with BRCA1, BRCA2, and ATM alterations. However, BRCA1/2-altered patients had the greatest benefit.40 Following these results, olaparib received FDA approval for patients with HRR-altered mCRPC who progressed after ARSI.40 Rucaparib also received approval for patients with mCRPC harboring somatic or germline BRCA1/BRCA2 mutations who previously received chemotherapy with a taxane.41 The recently published TRITON3 phase 3 trial showed progression-free survival (PFS) benefit and preliminary OS benefit for rucaparib after ARSI in patients with mCRPC with BRCA1/2-mutated tumors.20 However, there was no benefit regarding tumor responses or PFS for patients with defects in ATM.20 Moreover, the first-line mCRPC phase 3 trials PROPEL and MAGNITUDE trials showed PFS benefit for the combination olaparib or rucaparib, respectively, and abiraterone-prednisone in HRR-altered tumors, which was more relevant for BRCA1/2-mutated tumors.21,22 Similarly, biomarker analysis from the phase 2 TOPARP-B trial illustrated that the activity of olaparib in mCRPC was highly dependent on the underlying DDR defect.19 This study showed that the alterations conferred the greatest sensitivity were homozygous BRCA2 deletions, biallelic defects in PALB2, and ATM deficiency with protein loss.19 In conclusion, effective targeted treatment strategies are currently lacking for most non-BRCA DDR alterations, which constitute at least 50% of all DDR alterations (Fig. 2).
 FIGURE 2:
FIGURE 2: Molecularly targeted treatment for DNA repair altered metastatic prostate cancer. Established molecularly targeted treatment options for DDR-deficient mCRPC are PARPi and immunotherapy with ICIs. PARPi, in monotherapy or combined with ARSI, has demonstrated major clinical efficacy in BRCA2-altered and BRCA1-altered PCa, and tumors with biallelic loss of PALB2. dMMR/MSI-high PCa and a subset of CDK12-altered PCa are sensitive to immunotherapy with ICIs. The preclinical data point to the potential efficacy of ATR and ATM inhibitors in ATM-deficient and FANCA-deficient tumors, respectively. The efficacy of these drugs is currently being assessed in early-phase clinical trials in relapsed/refractory solid tumors. Defects in BRCA1, BRCA2, and, possibly, FANCA confer sensitivity to platinum-based chemotherapy. ARSI indicates androgen receptor signaling inhibition; DDR, DNA damage response; ICI, immune checkpoint inhibitor; mCRPC, metastatic castration-resistant PCa; PARPi, poly-ADP-ribose polymerase inhibitor; PCa, prostate cancer.
Immune Checkpoint InhibitorsImmunotherapy with ICIs is highly active in dMMR/MSI-high PCa tumors and has shown variable activity in CDK12-deficient tumors. On the contrary, ICIs have shown a very limited efficacy in biomarker unselected mCRPC, and PCa are globally considered immunologically cold tumors.82–86 The efficacy of the anti-PD1 ICI pembrolizumab was assessed in the KEYNOTE-158 phase 2 study in relapsed/refractory MSI-H/dMMR solid tumors. This study included 8 patients with mPCa and showed a response rate of 30.8% and a median OS of 20.1 months (95% CI: 14.1–27.1) for the global pan-tumor patient population.87 Based on these data, pembrolizumab is approved for the treatment of relapsed/refractory MSI-H/dMMR mPCa (https://www.accessdata.fda.gov/drugsatfda_docs/label/2021/125514s096lbl.pdf). Further work showed that alterations in MMR correlate with PD-L1 expression levels in PCa.88
In a cohort of 60 CDK12-altered PCa, around half of the patients had biallelic loss.59 These tumors are characterized by genomic instability, with frequent tandem duplications and gene fusions.59CDK12-deficient PCa poorly responds to treatment with ARSI, PARPi, or taxane-based chemotherapy and have a variable vulnerability to ICIs (Fig. 2).59
Emerging Therapeutic StrategiesData from preclinical studies have suggested that ATR inhibitors are active in ATM-deficient tumors and may act synergistically with PARPi and platinum-based chemotherapy.36,89–91 Combined ATR and PARP inhibition was synergistic in ATMKO clones of PCa cell lines (22RV1).89 These ATM-deficient cells showed variable sensitivities to rucaparib and platin but homogeneous vulnerability to irradiation. Preclinical data demonstrate that opposite to BRCA loss, KO of ATM alone does not lead to a classical HRD phenotype with a lack of Rad51 foci formation. However, a combined ATM and ATR inhibition does confer an HRD phenotype.89 The same results have also been observed when using DR-GFP reporters.89 Early-phase clinical trials with ATR inhibitors (eg, ATG-018, RP-3500, AZD6738) are ongoing for refractory DDR-altered solid tumors, including PCa. The phase 1 TRESR trial (NCT04497116), assessing the safety and preliminary efficacy of the ATRi camonsertib in DDR-deficient solid tumors showed a modest response rate (12%) in ATM-deficient mCRPC.92 A CHK1/2 inhibitor (AZD7762), which acts downstream of ATR, has shown sustained clinical activity in a RAD50-mutant, functionally ATM-deficient small-cell carcinoma.93 In FANCA-deficient cell lines, ATM inhibitors have shown preclinical activity.94,95 Several ATM inhibitors (eg, M4076, AZD0156) are being assessed in early-phase trials for advanced refractory solid tumors. To our knowledge, no targeted therapeutic strategies distinct from PARPi have been successfully developed for CHEK2-altered tumors. CRISPR screen performed in PCa cell lines showed that CHK2 loss confers resistance to PARPi, and that combined PARP and ATR inhibition can overcome this resistance (Fig. 2).96
Several commonly altered genes in PCa, such as AR, PTEN loss, or the TMPRSS2-ERG fusion, have been related to impaired DNA repair function.97 While conserved AR signaling enhances DDR response, ADT sensitizes PCa cells to DNA-damaging agents, such as radiotherapy, a synergy routinely used in the biochemical relapse setting.97,98 Loss of PTEN has been reported to be synthetically lethal with ATM inhibition in preclinical models.97,99TMPRSS2-ERG has been shown to downregulate the NHEJ pathway.97,100 Other common PCa genomic alterations related to impaired DDR function are SPOP mutations and loss of CHD1, both downregulating the HRR pathway.97 Moreover, a recent work uncovered the presence of lower levels of XRCC1 in formalin-fixed paraffin-embedded PCa tumor tissue of African American patients, as well as increased uracil and pyrimidine lesions and increased uracil DNA glycosylase levels, pointing to an impaired base excision repair pathway function.101 XRCC1 prevents the trapping of PARP1 during base excision repair, a DNA repair pathway that removes damaged or incorrect bases.102,103
DISCUSSIONDDR alterations are prevalent in advanced PCa and constitute a highly heterogenous group of molecular alterations leading to distinct DNA repair–deficient phenotypes. A yet unresolved question is how to optimally assess the presence of a DDR-deficient phenotype that could predict vulnerability to targeted treatment with PARPi. Previous studies have shown that ATM and CHEK2-altered PCa exhibit lower genomic instability scores, assessed as a combination of loss-of-heterozygosity, large-scale transitions, and telomeric allelic imbalance, as compared to tumors with BRCA2 alterations.104 Another study showed that PCa with alterations in BRCA1, BRCA2, FANCA, and ATR had higher loss-of-heterozygosity scores, assessed by targeted NGS with FoundationOne CDx, as compared, for instance, with CDK12-altred PCa.11 In a recent study, Ritch et al105 proposed using a machine-learning tool, DARC Sign, to identify DDR defects based on whole-exome sequencing of plasma circulating cell-free DNA. This model outperformed previous classifiers, such as CHORD106 or HRDdetect,107 based on analysis of whole-genome sequencing data.105 These results should be, however, validated in further independent cohorts.
From a therapeutic perspective, several clinical trials within the mCRPC setting have shown that BRCA2, BRCA1, and PALB2 alterations, especially in biallelic loss, are most vulnerable to targeted treatment with PARPi.19,20,40 These alterations lead to a classical HRD phenotype. On the other hand, PCa tumors with alterations in MMR genes, which lead to a dMMR/MSI-high phenotype, and alterations in CDK12, lead to a genomic instability phenotype with increased neoantigen burden. These tumors, especially dMMR/MSI-high, show high response rates to treatment with ICIs, which otherwise lack meaningful activity in PCa.87 However, other frequent alterations, such as ATM, are less vulnerable to PARP inhibition.20,40ATM-deficient PCa account for ≅5% to 7% of all PCa, concerning a numerically relevant cohort of patients. For ATM deficiency, preclinical studies point to increased sensitivity to ATR inhibitors. Currently, ATR inhibitors are being assessed in early clinical trials for advanced solid tumors, in monotherapy, and in combination with PARPi. For instance, in the refractory mCRPC setting, the phase 2 TRAP trial assesses the combined activity of the ATR inhibitor ceralasertib and the PARPi olaparib in DDR-deficient and proficient tumors.108 However, preliminary results point to a modest activity in ATM-deficient tumors. A recent preclinical study suggested that ATR inhibition might boost the efficacy of anti-PD-L1 tumors in PCa.109 Further, preclinical as well as clinical studies showed increased sensitivity to ionizing radiation in ATM-deficient tumors.110–112 Moreover, efficacy data from a cohort of patients with PCa treated radioligand therapy (177Lu-617-PSMA) point to increased PSA responses in patients carrying ATM pathogenic mutations.113 Preliminary results from the phase 1 LuPARP trial, which assessed the safety and preliminary efficacy of the combination of 177 Lu-617-PSMA radioligand therapy combined with olaparib, showed so far good safety profile but a similar rate of PSA50 responses as compared to treatment with177 Lu-617-PSMA alone.114
CONCLUSIONSDDR alterations are found in approximately a quarter of all PCa and constitute the most frequently altered genes in the germline. Distinct genomic alterations lead to highly heterogeneous DNA repair–deficient phenotype. To date, successful targeted treatment options have been established for a subset of DDR-deficient PCa, such as tumors with BRCA1 and BRCA2-alterations, which exhibit synthetic lethality with PARPi, and dMMR/MSI high tumors, which are vulnerable to ICIs. The discovery of novel DDR alterations that might sensitize PCa tumors to DNA-damaging therapies is of great relevance for further improving personalized cancer therapy.
ACKNOWLEDGMENTSThe authors thank Dr Mariana Ricca at the University of Bern for her editorial support. Figure 1 was generated with Biorender.com.
REFERENCES 1. Akhoundova D, Feng FY, Pritchard CC, et al. Molecular genetics of prostate cancer and role of genomic testing. Surg Pathol Clin. 2022;15:617–628. 2. Lozano R, Castro E, Aragón IM, et al. Genetic aberrations in DNA repair pathways: a cornerstone of precision oncology in prostate cancer. Br J Cancer. 2021;124:552–563. 3. van Dessel LF, van Riet J, Smits M, et al. The genomic landscape of metastatic castration-resistant prostate cancers reveals multiple distinct genotypes with potential clinical impact. Nat Commun. 2019;10:5251. 4. Li X, Heyer W-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008;18:99–113. 5. Gulhan DC, Lee JJ-K, Melloni GEM, et al. Detecting the mutational signature of homologous recombination deficiency in clinical samples. Nat Genet. 2019;51:912–919. 6. Rosenthal R, McGranahan N, Herrero J, et al. deconstructSigs: delineating mutational processes in single tumors distinguishes DNA repair deficiencies and patterns of carcinoma evolution. Genome Biol. 2016;17:31. 7. Pritchard CC, Mateo J, Walsh MF, et al. Inherited DNA-repair gene mutations in men with metastatic prostate cancer. N Engl J Med. 2016;375:443–453. 8. Robinson D, Van Allen EM, Wu YM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015;162:454. 9. Beltran H, Yelensky R, Frampton GM, et al. Targeted next-generation sequencing of advanced prostate cancer identifies potential therapeutic targets and disease heterogeneity. Eur Urol. 2013;63:920–926. 10. Abeshouse A, Ahn J, Akbani R, et al. The molecular taxonomy of primary prostate cancer. Cell. 2015;163:1011–1025. 11. Chung JH, Dewal N, Sokol E, et al. Prospective comprehensive genomic profiling of primary and metastatic prostate tumors. JCO Precis Oncol. 2019;3:1–23. 12. Rodriguez-Calero A, Gallon J, Akhoundova D, et al. Alterations in homologous recombination repair genes in prostate cancer brain metastases. Nat Commun. 2022;13:2400. 13. Chang HHY, Pannunzio NR, Adachi N, et al. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat Rev Mol Cell Biol. 2017;18:495–506. 14. Mao Z, Bozzella M, Seluanov A, et al. DNA repair by nonhomologous end joining and homologous recombination during cell cycle in human cells. Cell Cycle. 2008;7:2902–2906. 15. Ceccaldi R, Sarangi P, D’Andrea AD. The Fanconi anaemia pathway: new players and new functions. Nat Rev Mol Cell Biol. 2016;17:337–349. 16. Li G-M. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008;18:85–98. 17. Alexandrov LB, Kim J, Haradhvala NJ, et al. The repertoire of mutational signatures in human cancer. Nature. 2020;578:94–101. 18. Mateo J, Porta N, Bianchini D, et al. Olaparib in patients with metastatic castration-resistant prostate cancer with DNA repair gene aberrations (TOPARP-B): a multicentre, open-label, randomised, phase 2 trial. Lancet Oncol. 2020;21:162–174. 19. Carreira S, Porta N, Arce-Gallego S, et al. Biomarkers associating with PARP inhibitor benefit in prostate cancer in the TOPARP-B trial. Cancer Discov. 2021;11:2812–2827. 20. Fizazi K, Piulats JM, Reaume MN, et al. Rucaparib or physician’s choice in metastatic prostate cancer. N Engl J Med. 2023;388:719–732. 21. Saad F, Armstrong AJ, Thiery-Vuillemin A, et al. PROpel: phase III trial of olaparib (ola) and abiraterone (abi) versus placebo (pbo) and abi as first-line (1L) therapy for patients (pts) with metastatic castration-resistant prostate cancer (mCRPC). J Clin Oncol. 2022;40(suppl_6):11. 22. Chi KN, Rathkopf DE, Smith MR, et al. Phase 3 MAGNITUDE study: first results of niraparib (NIRA) with abiraterone acetate and prednisone (AAP) as first-line therapy in patients (pts) with metastatic castration-resistant prostate cancer (mCRPC) with and without homologous recombination repair (HRR) gene alterations. J Clin Oncol. 2022;40(suppl_6):12. 23. Robinson D, Van Allen EM, Wu YM, et al. Integrative clinical genomics of advanced prostate cancer. Cell. 2015;161:1215–1228.
留言 (0)