

記住我
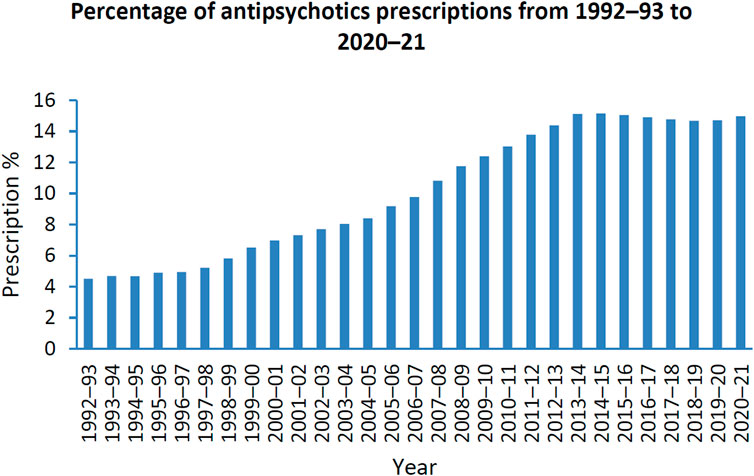
Antipsychotics are the first line treatment in schizophrenia (Lally and MacCabe, 2015). Schizophrenia is a state whereby patients lose connection with reality. This can be in the form of hallucinations, delusions and disorder of thoughts with far-reaching effects on individuals, their families and society (Howes et al., 2009; Patel et al., 2014). Global statistics has shown that schizophrenia is the eighth leading cause of disability in people aged 15–44 years (Haddad and Correll, 2018). Antipsychotics are also used off-label in the treatment of anxiety, borderline personality disorder, substance use disorders, eating disorders, post-traumatic stress disorder, obsessive-compulsive disorder, mood disorders, insomnia, agitation and attention deficit hyperactivity disorder (Wang et al., 2021a; Azimi Manavi et al., 2023). Between 2005–2014, the prescription rate of antipsychotic use overall has increased in 10 out of 16 countries, with the atypical antipsychotics, quetiapine, olanzapine and risperidone being most prescribed (Hálfdánarson et al., 2017). The Australian Institute of Health and Welfare has reported that 17.7% of Australians received mental health-related medications between 2020 and 2021 where the highest prescription rates were 8.9% and 8.7% for antidepressants and antipsychotics, respectively (Australian Institute of Health and Welfare, 2022). Moreover, the prescription of antipsychotics has increased by 10.5% from 1992-93 to 2020-21 (Australian Institute of Health and Welfare, 2022) (Figure 1).
FIGURE 1. Percentage of patients prescribed antipsychotics from 1992–1993 to 2020–2021; Data adopted from mental health services in Australia: Mental health related prescriptions Australian Institute of Health and Welfare (Australian Institute of Health and Welfare, 2022).
The use of antipsychotics is associated with a number of side effects including tardive dyskinesia (repetitive, involuntary movements commonly in areas of the face, eyes, and mouth), dystonia (involuntary muscle movements that can affect the head, face, and neck), akathisia (inability to remain still), weight gain, hyperglycaemia, type 2 diabetes and adverse cardiovascular impacts including sudden cardiac death (De Hert et al., 2011). There is a growing body of evidence suggesting that antipsychotics are also associated with negative, off-target side effects that include reductions in bone mineral density (BMD) and increased fracture risk (Kishimoto et al., 2008; Crews and Howes, 2012; Fraser et al., 2015; Bolton et al., 2017; May et al., 2019; Paschou et al., 2019; Azimi Manavi et al., 2023). The mechanism(s) by which antipsychotics induce bone loss are yet to be fully explored and understood. This has led to an emerging area of research to understand the underlying mechanisms by which antipsychotics negatively impact bone.
Bone is a multicellular organ consisting of different cell types, namely,: osteoclasts, osteoblasts, osteocytes, chondrocytes, bone lining cells, and bone-resident macrophages known as osteal macrophages or osteomacs (Florencio-Silva et al., 2015; Miron and Bosshardt, 2016; Kenkre and Bassett, 2018; Mohamad et al., 2021). Old and damaged bone is resorbed by osteoclasts while new bone is formed by osteoblasts, which, when entombed in newly formed bone become osteocytes, the most prevalent cells in bone tissue (∼90–95%) that act as a master regulator of osteoclast and osteoblast function (Metzger and Narayanan, 2019). Chondrocytes form the cartilage matrix which is later replaced by osteoblasts to undergo endochondral ossification (Chen et al., 2021), while resting osteoblasts are known as bone lining cells (Lee et al., 2021). Osteomacs also regulate bone modelling and remodelling by acting as potential osteoclast precursors and secreting oncostatin M to promote bone formation by osteoblasts (Walker et al., 2010; Mohamad et al., 2021; Hioki et al., 2022). The fine balance between osteoclast and osteoblast activity is central to maintaining normal bone homeostasis and resultant healthy bone (Florencio-Silva et al., 2015).
Antipsychotics target different receptors including dopamine, serotonin and adrenergic receptors, which are predominantly located in the brain, but have recently been shown to be present in osteoclasts and osteoblasts (Aringhieri et al., 2018). Therefore, this review summarises commonly prescribed antipsychotics and details their mechanisms of action, in addition to how they may influence various dopamine, serotonin and adrenergic receptors shown to be expressed in osteoclasts and osteoblasts that may ultimately lead to bone loss.
2 AntipsychoticsInitially, antipsychotics categorised as either “typical” or “atypical.” This has changed first-generation antipsychotics (FGA), second-generation antipsychotics (SGA) and third-generation antipsychotics (TGA). These categories are primarily based on their affinity to dopamine and serotonin receptors, where they act as either agonist, partial agonist, antagonist or an inverse agonist (Strange, 2008). First generation antipsychotics are dopamine D2 (D2) antagonists, which have been effective in the treatment of positive symptoms while less effective in the treatment of negative symptoms (Mailman and Murthy, 2010; Li et al., 2016). Positive symptoms include hallucinations, delusions and disorder of thoughts while the negative symptoms cause dysfunction in personal, social and occupational functioning through means such as anhedonia, avolition and social withdrawal, as well as, cognitive dysfunction (Winship et al., 2019). The first generation antipsychotic “chlorpromazine” was the first to be recognised as a psychotropic drug in 1952 by Henri Laborit (Laborit et al., 1952; Ban, 2007). Following chlorpromazine, other FGAs including haloperidol, trifluperazine, thioridazine and fluphenazine were introduced (Ramachandraiah et al., 2009). First generation antipsychotics are associated with a number of undesirable side effects including hyperprolactinemia, sedation, weight gain, autonomic and cardiovascular effects, akathisia, parkinsonism, dystonia and tardive dyskinesia (Mailman and Murthy, 2010), thus, spurring on the development of second generation antipsychotics. Second generation antipsychotics are D2 and serotonin (5HT) antagonists with affinity ratio greater than 1.12 for D2/5HT2A which have been more effective in reducing negative and cognitive symptoms compared to FGAs, but associated with greater weight gain and metabolic syndromes such as hypertriglyceridemia, elevated glucose, insulin and low-density lipoprotein cholesterol levels (Lieberman, 2004; Mailman and Murthy, 2010; Pillinger et al., 2020; Kiss et al., 2022). A third generation of antipsychotics such as aripiprazole, cariprazine and brexpiprazole were introduced most recently and have been successful in treating positive, negative and cognitive symptoms (Vasiliu, 2022). These predominantly act as pre-synaptic and post-synaptic D2 partial agonists rather than receptor antagonists, as well as D3 and 5HT1A partial agonists, while simultaneously acting as a serotonin 5HT2A and 5HT2B antagonist (Li et al., 2016; Kiss et al., 2022). Current research is focusing on D3 targeting antipsychotics due to their higher affinity for dopamine than D2 receptors (Maramai et al., 2016). Moreover, D3 are predominantly expressed in the limbic system, which is involved in schizophrenia pathology (Kiss et al., 2022) and the blockage of the D3 is associated with pro-cognitive effects in rats (Watson et al., 2012). As shown in Table 1, antipsychotics possessaffinity affinity to other receptors including adrenergic, histamine and muscarinic receptors (Table 1).
TABLE 1. Receptor binding profiles for commonly prescribed antipsychotics.
3 Schizophrenia and bone health: epidemiological findingsThere is a growing body of evidence to show that schizophrenia is associated with osteoporosis, a degenerative bone disease characterised by low BMD and increased fracture risk (Jung et al., 2011; Lally, 2020; Forns et al., 2021; Azimi Manavi et al., 2023). The onset of schizophrenia occurs primarily in adolescents and young adults, where peak bone mass is yet to be achieved (Forns et al., 2021; Zhu and Zheng, 2021). Patients with schizophrenia are possibly vulnerable to bone defects due to lifestyle factors such as a reduced physical activity, smoking, alcohol consumption, vitamin D deficiency and poor dietary habits in addition to genetic and biological factors (Coentre et al., 2009). A study has shown that smoking and alcohol consumption also leads to further reduced BMD in patients with schizophrenia compared to those who do not smoke or drink alcohol (Jung et al., 2011). Moreover, studies show that the use of antipsychotics further exacerbates bone loss and fracture risk in patients with schizophrenia (Oderda et al., 2012; Rigler et al., 2013; Fraser et al., 2015; Bolton et al., 2017; May et al., 2019; Paschou et al., 2019; Shen et al., 2019). Similarly, studies have shown that increased bone fractures are linked with schizophrenia compared to controls (Tsai et al., 2014; Stubbs et al., 2015; Tseng et al., 2015).
4 Antipsychotic use and bone health: epidemiological, in vitro and in vivo findingsEpidemiological evidence is growing to suggest antipsychotic use (independent of schizophrenia) is associated with lower BMD, increased fracture risk (Oderda et al., 2012; Rigler et al., 2013; Fraser et al., 2015; Bolton et al., 2017; May et al., 2019; Paschou et al., 2019; Shen et al., 2019; Azimi Manavi et al., 2023) and occurrence of re-fracture (Shen et al., 2019). Some studies suggest typical antipsychotics have a higher fracture risk compared to atypical antipsychotics (Ching-Min et al., 2020), while others suggest there is no difference with regards to the type of antipsychotics and associated fracture risk (May et al., 2019; Shen et al., 2019). Risperidone has been shown to reduce BMD in children and young adults (Calarge et al., 2010; Rosenbloom, 2010) with a significant decrease in BMD compared to olanzapine or other SGAs (Becker et al., 2003; Clapham et al., 2020). It has been reported that patients receiving either FGAs or SGAs are at higher fracture risk as antipsychotics in these categories such as chlorpromazine, risperidone and zotepine increased incident fracture, as well as, re-fracture (Ching-Min et al., 2020). A meta study concluded that FGA users show a 77% increase in hip fractures compared to non-users, while SGA users show a 41% increase. This study also noted that associated fracture risk is 29%, 46%, 49% and 94% for risperidone, olanzapine, quetiapine and haloperidol, respectively (Papola et al., 2018; Shen et al., 2019). Interestingly, research has shown that antipsychotics can accumulate up to 15-fold higher concentration in bone marrow compared to plasma (Houseknecht et al., 2017; May et al., 2019).
Evidence from in-vitro (Lahdelma et al., 2010; Kawamura et al., 2011; Motyl et al., 2012; Li et al., 2019) and in-vivo bone models (Oh-ie et al., 2002; Kunimatsu et al., 2010; Kawamura et al., 2011; Motyl et al., 2012) support these findings. In experiments conducted in rat primary cells, clozapine inhibited osteoblast mitogenesis and differentiation, as well as, osteoclast formation, whereas haloperidol had no effect (Costa et al., 2011). In mouse bone marrow mesenchymal stem cells (BMSCs), chlorpromazine inhibited osteoclastogenesis via inhibiting the intracellular calmodulin pathway (Kawamura et al., 2011). Risperidone significantly stimulated osteoclast differentiation without affecting osteoblast differentiation (Motyl et al., 2012). In vitro studies on antipsychotic-related bone defects in primary human cells are scant. A study showed clozapine has cytotoxic effects on primary cultures of human BMSCs (Lahdelma et al., 2010). A study using the human osteoblast cell line hFob1.19 showed that risperidone, amisulpride, olanzapine and aripiprazole inhibited osteoblasts cell viability and caused apoptosis in a dose dependent manner (Li et al., 2019).
In vivo evidence has similarly shown negative effects of antipsychotics on bone. Risperidone reduced trabecular and cortical bone mass in mice with increased osteoclast differentiation and resorption using bone marrow derived cells (Motyl et al., 2012). Chlorpromazine and haloperidol have also been shown to decrease BMD in trabecular bone of mice via increased resorption (Oh-ie et al., 2002; Kunimatsu et al., 2010). However, this induced bone loss through dopamine blockers is considered to be rodent specific (Kunimatsu et al., 2010). Clozapine, but not haloperidol, decreased BMD in-vivo in mice and reduced osteoclast and osteoblast activity in-vitro (Kawamura et al., 2011). Chlorpromazine also reduce osteoclast formation and osteoclastic gene expression in mouse BMSCs (Lahdelma et al., 2010).
5 Antipsychotics and bone cell function: importance of dopamine receptors/signalling in osteoclasts and osteoblastsDopamine is a neurotransmitter that controls sleep, motivation, reward, attention, voluntary movements, vision, hormonal regulation and motor functions (Hornykiewicz, 2017; Yang and Tsai, 2017; Wang et al., 2020). Abnormalities in the dopamine system are associated with schizophrenia and psychosis, where positive symptoms are linked with increased dopamine levels in the mesolimbic pathway associated with dopamine D2 hyper-activity (McCutcheon et al., 2018; Stahl, 2018; McCutcheon et al., 2019). Accordingly, most antipsychotics have been developed to target dopamine D2. Decreased levels of dopamine in cortical pathway has also been observed using PET scanning in patients with schizophrenia, which needs further study as it is believed that hypo-activity of the mesocortical pathway is associated with negative and cognitive symptoms in schizophrenia (Li et al., 2016; Weinstein et al., 2017). Even though this theory has been accepted for decades, recent data indicate the involvement of dorsal striatum in pathophysiology of schizophrenia (McCutcheon et al., 2019).
Dopamine receptors are G-protein-coupled receptors (GPCR), which belongs to GPCR alpha subunits Gs (stimulatory receptors) and Gi (inhibitory receptors) (Zhang and Shi, 2016; Martel and Gatti McArthur, 2020). There are five dopamine receptors, which are sub-categorised as D1-and D2-like, depending on pharmacology and ability to regulate cyclic adenosine monophosphate (cAMP) (Martel and Gatti McArthur, 2020). Dopamine D1 and D5 belong to D1-like receptors that modulate signalling pathways via Gs and increase cAMP production while D2, D3 and D4 belong to D2-like receptors that modulate via Gi and inhibit cAMP production (Figure 2) (Neve et al., 2004; Martel and Gatti McArthur, 2020). Moreover, D2-like receptors have 10- to 100-fold higher affinity for dopamine than D1-like receptors (Martel and Gatti McArthur, 2020). This leads to the assumption that extracellular dopamine levels or changes to dopamine levels from antipsychotic use may have different impacts on D2-like receptors compared to D1-like receptor signalling (Martel and Gatti McArthur, 2020). Dopamine has been found in bone marrow in higher concentrations (∼10 µM) than typically found in circulation (Matt and Gaskill, 2020). Moreover, hypothalamic neurons produce dopamine to regulate bone metabolism via hypothalamic-pituitary-gonadal axis while sympathetic nerves also contain dopamine which can penetrate to bone marrow (Wang et al., 2021b).
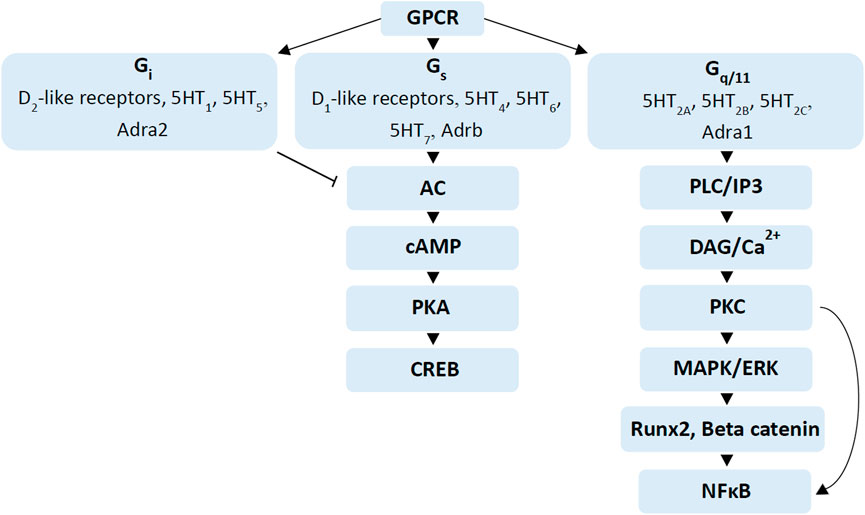
FIGURE 2. Signalling cascades regulated by dopamine, serotonin and adrenergic receptors. G-protein coupled receptor stimulation by different neurotransmitters such as dopamine, serotonin and adrenergic receptors regulate different signalling cascades. Activation of Gi receptors such as D2-like receptors, 5HT1, 5HT5 and Adra2 inhibit adenylate cyclase (AC), while activation of Gs receptors such as D1-like receptors, 5HT4, 5HT6, 5HT7 and Adrb stimulate AC to increase cAMP levels, which in turn activate the PKA/CREB pathway (Zhao et al., 2016; Ramaswamy et al., 2017; Wang et al., 2018; Martel and Gatti McArthur, 2020). Gq/11 receptors activate PLC-IP3 and DAG-PKC signalling pathways, which activate the mitogen-activated protein kinase (MAPK)/ERK signalling cascade (Gustafsson et al., 2006a; Dai et al., 2014; McCorvy and Roth, 2015). Activation of MAPK/ERK also stimulates Runx2 and beta-catenin in osteogenesis while activation of Protein kinase C (PKC) stimulates nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB), important in osteoclast differentiation (Abu-Amer, 2013; Zhao et al., 2016; Kim et al., 2019).
5.1 Dopamine signalling in osteoclastsDopamine has been demonstrated to inhibit osteoclastogenesis in a dose-dependent manner using osteoclast precursors derived from: i) human CD14+ cells isolated from peripheral blood mononuclear cells (PBMCs) (Hanami et al., 2013); ii) murine BMSCs (Yang et al., 2016); and iii) RAW 264.7 cells (Wang et al., 2021b). There are limited studies showing links between dopamine and bone cell function. Studies have reported that dopamine can interact with receptors in osteoclasts derived from human BMSCs to inhibit resorption via nuclear factor of activated T-cells cytoplasmic 1 (NFATc-1) and c-Fos signalling pathways (Hanami et al., 2013; Wang et al., 2020). Hanami et al. (2013) showed that dopamine D2-like signalling inhibits osteoclastogenesis in-vitro, which could be a protective mechanism against bone resorption while Wang et al. (2021) showed that dopamine inhibits osteoclastogenesis via D2R/cAMP/protein kinase A (PKA)/cAMP response element-binding protein (CREB) pathway in primary mice BMSCs (Wang et al., 2021b). In contrast, it has been shown that D1-like receptor antagonist SCH23390 inhibits osteoclastogenesis in mice bone marrow-derived macrophages (Nakashioya et al., 2011).
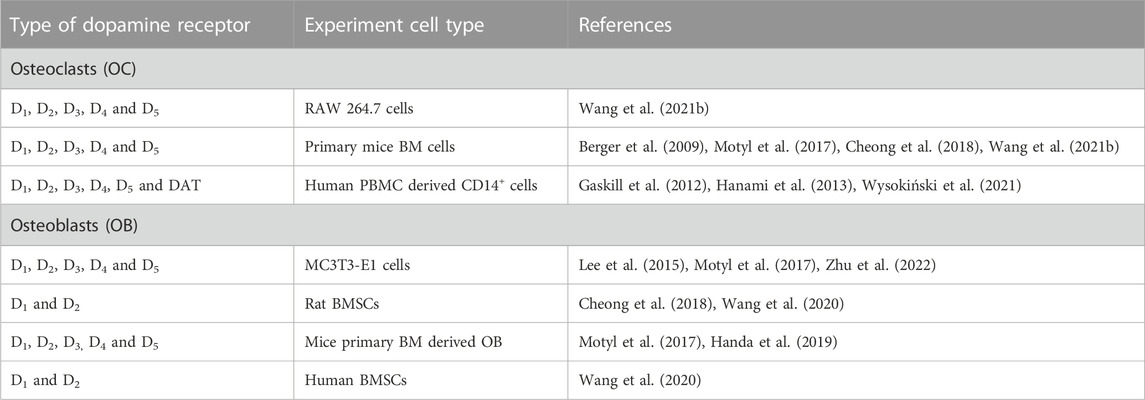
Expression of dopamine receptors in osteoclast has been reported in a number of animal and human models (Table 2). The presence of all five dopamine receptors and the dopamine transporter (DAT) have been reported in human CD14+ monocytes at both transcriptional and protein levels (Gaskill et al., 2012; Hanami et al., 2013). Maximum expression was observed in the early stages of osteoclastogenesis, decreasing after day 7 (Hanami et al., 2013). Similarly, all five dopamine receptors were identified in RAW 264.7 cells and in primary mice bone marrow derived macrophage-lineage cells from C57BL/6J mice (Wang et al., 2021b), in addition to mouse BMSCs, where increased expression of D1, D2 and D3 was observed during osteoclastogenesis (Cheong et al., 2018).
TABLE 2. Dopamine receptor expression in osteoclasts and osteoblasts.
5.2 Dopamine signalling in osteoblastsThe effects of dopamine on osteoblasts is more contentious. A study by Lee et al. (2015) reported that dopamine enhances osteoblast proliferation and mineralisation in-vitro in a MC3T3-E1 pre-osteoblastic murine cell line (Lee et al., 2015), while another study by Motyl et al. (2017) directly refuted these findings, reporting suppression of osteoblast mineralisation and osteoblast markers of gene expression (Motyl et al., 2017). The expression of all five dopamine receptors in osteoblasts has been shown at both mRNA and protein levels in MC3T3-E1 cells (Lee et al., 2015; Zhu et al., 2022). Dopamine (50 µM) did not affect osteoblast cell proliferation but increased mineralisation, compared to control, whereas concentrations above 100 µM induced cell death (Lee et al., 2015). Bone marrow mesenchymal stem cells can be differentiated into cartilage, tendon, muscle, fat and osteoblasts (Caplan, 1991; Pittenger et al., 1999; Sacchetti et al., 2007; Méndez-Ferrer et al., 2010; Wang et al., 2020). Primary murine BMSCs-derived osteoblasts express dopamine receptors, which regulate proliferation and differentiation (Motyl et al., 2017; Cheong et al., 2018; Handa et al., 2019; Wang et al., 2020). The expression of D1 and D2 was significantly increased from day 3 to day 7 in rat BMSCs (Wang et al., 2020). Wang et al. (2020) showed the presence of dopamine receptors D1 and D2 in human BMSCs where the expression of D1 increased until day 5 and the expression of D2 decreased from day three to day 5 (Wang et al., 2020). Moreover, alkaline phosphatase activity (ALP), mineralisation and expression of bone sialoprotein, runt-related transcription factor 2 (Runx2), and osteocalcin were stimulated with dopamine (5 nM). This stimulation was lost at 50 µM, whereas 500 μM was toxic (Wang et al., 2020). This study also demonstrated that dopamine-induced osteogenic differentiation of human BMSCs was mediated through D1 receptor activation via extracellular signal-regulated kinase (ERK) 1 and 2 signalling pathways and activation of Runx2 transcriptional activity, while the suppression of cAMP/PKA pathway may inhibit dopamine-induced human BMSCs differentiation (Wang et al., 2020).
Antipsychotics targeting D3 receptors have gained more attention in treating impaired cognitive, social, and motor functions associated with schizophrenia. To our knowledge, there are no studies on the effects of D3 on bone remodelling. Hence, it is important to investigate the effects of D3 in bone cells function, due to its similarity to D2, which can also activate ERK/MAPK signalling pathway (Kiss et al., 2022), which is important in bone homeostasis and skeleton development (Kim et al., 2019). Moreover, both D2 and D3 activate their signalling cascades in GPCR independent manner via beta arresting mediated pathways (van Gastel et al., 2018). Studies show that beta arrestin 2 is involved in regulating MAPK/AKT/glycogen synthase kinase 3 (GSK3) pathways downstream of the dopamine receptors (Del'Guidice et al., 2011). Glycogen synthase kinase 3 is expressed in the brain and important in psychiatric diseases such as schizophrenia (van Gastel et al., 2018). Interestingly, both D2 and D3 are positively coupled to beta arrestin/AKT/GSK3 pathway (van Gastel et al., 2018). Beta arrestin 2 stimulates mice trabecular bone formation but not bone resorption indicating the importance of this pathway in bone homeostasis (Gesty-Palmer et al., 2009).
In summary, the presence of all dopamine receptors in osteoclasts has been shown using RAW 264.7, primary mice BMSCs and human CD14+ cells where expression increased during osteoclastogenesis (Table 1). The presence of dopamine receptors in osteoblasts has also been shown in MC3T3-E1 (with no difference in expression level for 14 days), rat BMSCs (with increased D1 and D2 from day 3 to 7) and human BMSCs (with increased D1 expression until day 5 while D2 expression decreased after day 3) (Lee et al., 2015; Wang et al., 2020). It has also been demonstrated that low concentrations of dopamine (5 nM) can increase osteoblast marker expression (Wang et al., 2020). Moreover, dopamine regulates osteoclastogenesis via nuclear factor of activated T Cells 1 (NFATc1), cFOS, cAMP/PKA/CREB via D2 receptor while dopamine regulates osteoblastogenesis via ERK 1 and 2, Runx2 and cAMP/PKA via the D1 receptor (Wang et al., 2020; Wang et al., 2021b). However, there is limited data available on the effect of dopamine on human models. As all antipsychotics primarily target dopamine receptors as antagonists or partial agonists it is important to understand the dopamine receptor expression profile in bone cells to understand the complex mechanism of antipsychotics-induced bone loss.
6 Antipsychotics and bone cell function: importance of serotonin receptors/signalling in osteoclasts and osteoblastsSerotonin is a product of tryptophan metabolism and is produced both in the central nervous system and peripherally in gastrointestinal tract and in platelets, where it does not cross the blood brain barrier (Brenner et al., 2007; Kanova and Kohout, 2021). Peripheral serotonin accounts for the majority of serotonin produced in the body (∼95%), while the brain is responsible for only a minor fraction (Ducy and Karsenty, 2010; El-Merahbi et al., 2015). Centrally, serotonin acts as a neurotransmitter, while peripherally its role is as a hormone (Kanova and Kohout, 2021). As a neurotransmitter, serotonin regulates mood, reward, anger, perception, appetite, aggression, attention, memory and sex drive. Peripherally, serotonin regulates major organ functions including glucose homeostasis and lipid metabolism (Berger et al., 2009; Kanova and Kohout, 2021). It has been reported that serotonin acts centrally to inhibit bone resorption and increase bone formation, whilst peripherally it acts to directly inhibit bone formation (Ducy and Karsenty, 2010; El-Merahbi et al., 2015). It has been reported that osteoclasts can synthesise serotonin in the presence of receptor activator of nuclear factor kappa-Β ligand (RANKL). Osteoblast-derived RANKL stimulates the expression of tryptophan hydroxylase 1 (TPH1) by osteoclasts, which results in serotonin synthesis that may then play a role as a paracrine/autocrine factor to further regulate osteoclast formation and resorption (Chabbi-Achengli et al., 2012).
There are seven types of serotonin receptors, 5HT1 to 5HT7, which are all GPCRs, except 5HT3 (Nichols and Nichols, 2008; McCorvy and Roth, 2015). These receptors modulate different signalling pathways via Gi, guanine nucleotide binding protein alpha 11 (Gq/11), and Gs (Figure 2) (McCorvy and Roth, 2015). The receptors 5HT1 and 5HT5 belong to the Gi protein-coupled receptors, while the 5HT2A, 5HT2B and 5HT2C receptors belong to the Gq/11 protein-coupled receptors and 5HT4, 5HT6 and 5HT7 belong to Gs protein-coupled receptors (Dai et al., 2014; McCorvy and Roth, 2015). The Gi protein-coupled receptors 5HT1 and 5HT5 suppress the cAMP/PKA pathways, inhibiting bone formation (Yadav et al., 2008; Dai et al., 2014; McCorvy and Roth, 2015). It has also been shown that activation of the cAMP/PKA pathway is associated with increased osteogenesis in-vitro and increased bone formation in-vivo (Siddappa et al., 2009). In contrast, in OC, the inhibition of PKA leads to the phosphorylation of activating transcription factor 4 (ATF4), which stimulates osteoclast differentiation (Kode et al., 2012). Gq/11 protein-coupled receptors, 5HT2A, 5HT2B and 5HT2C activate phospholipase C-inositol phosphate 3 (PLC-IP3) and diacylglycerol-protein kinase C (DAG-PKC) signalling pathways, which promote osteoblast differentiation and bone formation while Gs protein-coupled receptors 5HT4, 5HT6 and 5HT7 increase cAMP levels (Gustafsson et al., 2006a; Dai et al., 2014; McCorvy and Roth, 2015).
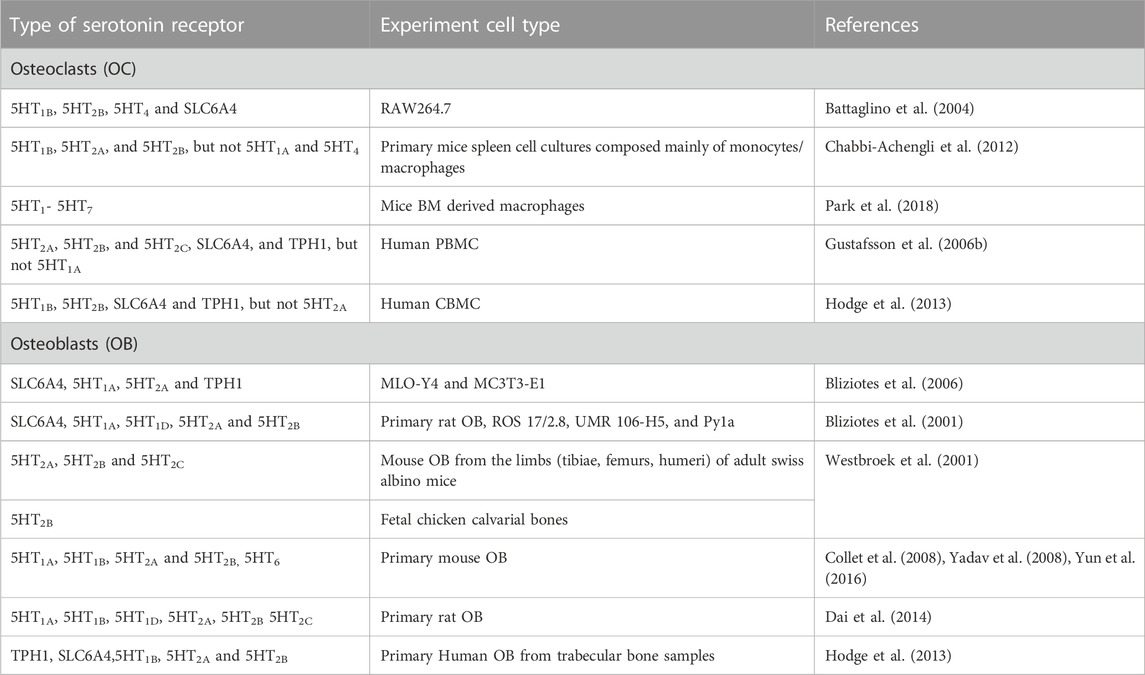
6.1 Serotonin signalling in osteoclastsThe localisation of serotonin receptors has been extensively reported in animal and human in-vitro and in-vivo bone cell models, with varying expression profiles of serotonin receptors in osteoclasts (Table 3). Expression of 5HT1B, 5HT2A, 5HT4 and serotonin transporter (SLC6A4) has been reported in differentiated RAW264.7-derived osteoclasts (Battaglino et al., 2004), while the presence of all serotonin receptors (5HT1- 5HT7) have been demonstrated in mouse BMSCs, suggesting a role for these receptors in osteoclasts differentiation (Park et al., 2018). The presence of 5HT1B, 5HT2A, and 5HT2B was shown in osteoclast precursors from mice spleen cells and not 5HT1A and 5HT4 (Chabbi-Achengli et al., 2012). The expression level changed with RANKL exposure where 5HT1B receptor expression increased and 5HT2B expression decreased with no change in the 5HT2A receptor expression (Chabbi-Achengli et al., 2012). This could suggest that 5HT1B may be important in osteoclast differentiation from precursors to mature osteoclasts while 5HT2A may act before osteoclasts lineage begins (Chabbi-Achengli et al., 2012). Gustafsson et al. (2006) showed the presence of 5HT2A, 5HT2B, 5HT2C, SLC6A4 and TPH1 but not 5HT1A in human PBMC-derived osteoclasts (Gustafsson et al., 2006b). The presence of 5HT1B, 5HT2B, SLC6A4 and TPH1 in human umbilical cord blood-derived osteoclast precursors have been shown, while the level of 5HT2B expression increased in mature osteoclasts (Hodge et al., 2013).
TABLE 3. Serotonin receptor expression in osteoclasts and osteoblasts.
Serotonin has been shown to increase osteoclastogenesis by stimulating both osteoclast formation and resorption in human PBMC derived osteoclasts (Gustafsson et al., 2006b). It has been reported that RANKL induces serotonin synthesis in mice osteoclast precursors whereas serotonin synthesis decreased as osteoclasts matured (Chabbi-Achengli et al., 2012). The study by El-Merahbi, Löffler et al. (2015) also found that 5HT1B and 5HT2A are involved in mediating serotonin in osteoclast differentiation. Moreover, selective blockage of 5HT1B, 5HT2B and 5HT4 did not affect osteoclastogenesis in RAW264.7 cells (Battaglino et al., 2004). In contrast, antagonism at 5HT1B and 5HT4 in mouse BMSCs-derived osteoclasts inhibit osteoclast marker expression (Battaglino et al., 2004). This may be because RAW264.7 cells are more differentiated than osteoclast precursors used in bone marrow cell preparation, also suggesting the importance of 5HT1B and 5HT4 in mediating osteoclast differentiation (Battaglino et al., 2004). The osteoclasts activity was inhibited by serotonin or serotonin agonists for 5HT1, 5HT4, and 5HT7, with no effect on osteoclasts activity with 5HT2 and 5HT5 agonists in mice BMSCs (Park et al., 2018). Serotonin has been shown to increase osteoclastogenesis at concentrations of 10 μM or below in mice BMSCs derived osteoclasts, with 5HT6 being more significant (Park et al., 2018). Moreover, it has been reported that mast cells release serotonin locally at high concentration (Battaglino et al., 2004). Mast cells in bone marrow are reported to increase in ovariectomised rats that suffered rapid bone loss (Lesclous et al., 2001). These data suggest that serotonin may modulate osteoclastogenesis in-vivo.
6.2 Serotonin signalling in osteoblastsThe expression of SLC6A4, 5HT1A, 5HT1B, 5HT1D, 5HT2A, 5HT2B, 5HT2C and TPH1 have been reported in primary cultures of rat osteoblasts, in addition to the osteoblastic cell lines; ROS 17/2.8, UMR 106-H5, Py1a, MLO-Y4 and MC3T3-E1, with the highest expression of 5HT1B and 5HT2A observed in both the early and late stages of differentiation (Bliziotes et al., 2001; Dai et al., 2014). The presence of TPH1 indicates that these cell lines may have the capacity to synthesise serotonin that may subsequently act in a paracrine/autocrine manner on bone cells in the bone remodelling space. Receptor 5HT2B has been demonstrated in fetal chicken bone, where this receptor had the highest mRNA expression in skeleton tissue compared to the other tested tissues (Westbroek et al., 2001). The presence of 5HT2A, 5HT2B and 5HT2C also observed in murine osteoblasts, where 5HT2A and 5HT2B had higher mRNA expression compared to 5HT2C (Westbroek et al., 2001). Serotonin decreased osteoblast proliferation and mineralisation from 1 nM to 10 μM, where there is an increased trend of proliferation and mineralisation after 1 µM (Dai et al., 2014). Collet et al. (2008) reported 5HT2B receptor expression increased during osteoblast differentiation in bone marrow derived mouse osteoblasts while 5HT2B−/- leads to osteopenic phenotype (Collet et al., 2008). Yadav et al. (2008) also reported the presence of 5HT1B, 5HT2A and 5HT2B receptors in primary mouse osteoblasts and that serotonin inhibited osteoblast proliferation via 5HT1B by inhibiting cAMP/PKA/CREB while there was no effect of 5HT2A and 5HT2B on bone mass (Yadav et al., 2008). In contrast, it has been shown that 5HT2B receptor activation was responsible for increased ALP activity and mineralisation in mice (Baudry et al., 2010). Hodge et al. (2013) reported the presence of TPH1, SLC6A4, 5HT1B, 5HT2A and 5HT2B in mature primary human osteoblasts with the highest expression of TPH1. Yun et al. (2016) showed that 5HT1-7 expression is higher in bone compared to brain, where the 5HT6 expression was the highest followed by 5HT2 in primary mice calvarial osteoblasts where 5HT6 inhibited osteoblast differentiation via Jab1 in BMP2 signalling but not PKA and ERK1/2 (Yun et al., 2016). This data suggest the possible role of 5HT6 in regulating abnormal osteoblasts differentiation.
In summary, expression of serotonin receptors has been clearly demonstrated in both animal and human models of osteoclast- and osteoblastogenesis. Receptors 5HT1B, 5HT2A, 5HT2B and 5HT4 are found to be implicated in regulating osteoclast and osteoblast activity and more specifically the loss of 5HT2B leads to decreased bone formation (Battaglino et al., 2004; Collet et al., 2008; Baudry et al., 2010; Chabbi-Achengli et al., 2012; Hodge et al., 2013; Park et al., 2018) while 5HT6 may important in regulating osteoblasts differentiation (Yun et al., 2016). There are relatively few reports available, however, that detail associated signalling pathways downstream of serotonin receptor binding in bone cell function, especially in human models. Therefore, it is important to further investigate serotonin receptor expression profile in human bone cells to better understand antipsychotic-induced bone loss as SGA and TGA have varying affinities to these receptors.
7 Antipsychotics and bone cell function: importance of adrenergic receptors/signalling in osteoclasts and osteoblastsAdrenergic receptors (adrenoceptors) are GPCRs that are involved in activating neurotransmitters; noradrenaline (norepinephrine) and adrenaline (epinephrine) (Graham, 1990). These receptors are expressed in the peripheral and central nervous system as hetero receptors inhibiting the release of neurotransmitters such as serotonin, gamma-aminobutyric acid (GABA) and dopamine (Cruz Grecco Teixeira et al., 2016). There are two main types of adrenergic receptors; alpha (α) and beta (β), with multiple subtypes. Alpha receptors have subtypes α1 (Adra1) and α2 (Adra2) with further sub-categories α1A (Adra1a), α1B (Adra1b), α1D (Adra1d), and α2A (Adra2a), α2B (Adra2b), α2C (Adra2c) (Perez, 2020) while beta receptors have subtypes β1 (Adrb1), β2 (Adrb2) and β3 (Adrb3). The Adra1, Adra2 and Adrb modulate signalling pathways through Gq/11, Gi and Gs, respectively (Figure 2) (Bylund et al., 2013).
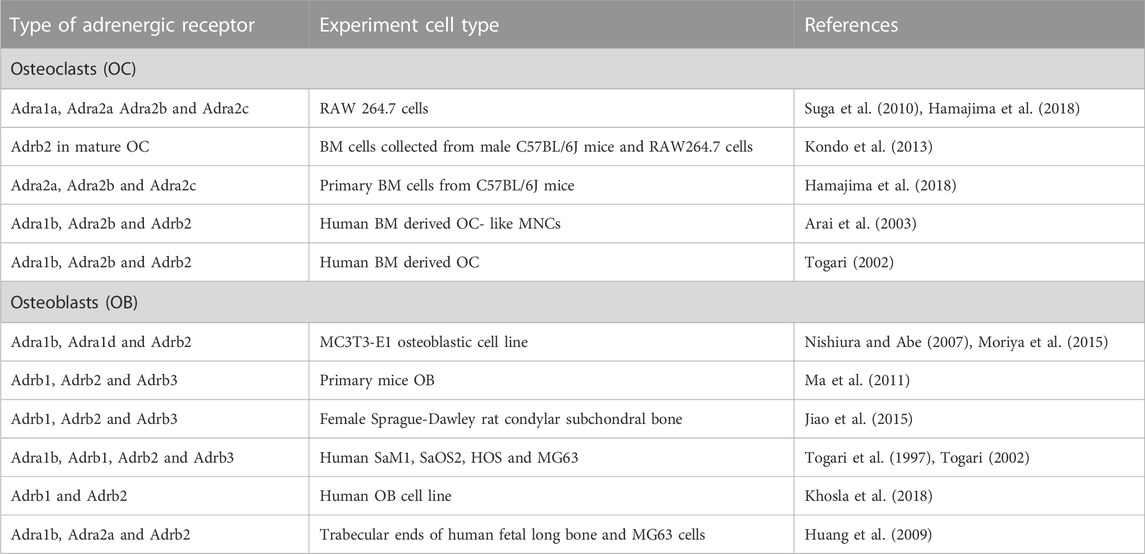
7.1 Adrenergic signalling in osteoclastsMammalian bone is innervated by sympathetic nerves (Hill and Elde, 1991; Khosla et al., 2018). In normal bone remodelling, noradrenaline is released by sympathetic nerves, which are known to inhibit bone formation and stimulate bone resorption (Khosla et al., 2018). Adrenergic receptor expression has been shown in human osteoclasts and osteoblasts (Table 4). The expression of β-adrenergic receptors have been demonstrated in both osteoclasts and osteoblasts (Kondo et al., 2013). Activation of β-adrenergic receptors regulate the secretion of cytokines by osteoblasts, which indirectly stimulated osteoclast activity in human bone marrow-derived osteoclast-like mononuclear cells (Arai et al., 2003; Zhong and Xia, 2021). Activation of Adrb2 inhibited CREB phosphorylation and subsequent osteoblast proliferation, which result in increased ATF4 phosphorylation and RANKL expression leading to increased osteoclast activity and bone resorption (Elenkov et al., 2001; Kajimura et al., 2011; Zhong and Xia, 2021). It has been suggested that β–adrenergic receptor stimulation may also increase RANKL production by T cells, which also may drive osteoclastogenesis (Elenkov et al., 2001; Bonnet et al., 2008).
TABLE 4. Adrenergic receptor expression in osteoclasts and osteoblasts.
Animal studies have shown that β-receptor agonists increase the levels of prostaglandin 2 and interleukin-6 (IL-6), which promotes osteoclast proliferation and resorption in mice skulls, thereby decreasing bone mass (Zhong and Xia, 2021). The mouse osteoclast precursor cell line RAW264.7 expresses Adrb2 and these receptors stimulate osteoclastogenesis via reactive oxygen species production (Zhong and Xia, 2021). The presence of all three Adra2 receptors in RAW264.7 cells have been reported on days 0, 2 and 4 after administration of RANKL while there was no significant difference in receptor expression between days 0, 2 and 4 (Hamajima et al., 2018). In contrast, in mice primary bone marrow cells, Adra2b showed to be upregulated after RANKL administration while Adra2a and Adra2c expression was significantly downregulated (Hamajima et al., 2018). Similarly, expression of Adrb2, but not Adrb1 and Adrb3, is detected in mature osteoclasts derived from mice BMSCs (Zhong and Xia, 2021). It has also been reported that Adra2 agonists suppress osteoclastic gene expression in RAW264.7 cells and in primary bone marrow cells, namely; NFATc1, tartrate-resistant acid phosphatase (TRAP) and cathepsin K, with a corresponding decrease in TRAP positive multi-nucleated osteoclast number in mouse BMSCs (Hamajima et al., 2018).
7.2 Adrenergic signalling in osteoblastsReceptor Adrb2 is thought to be the main receptor expressed in sympathetic nerves responsible for the regulation of bone mass via osteoblasts (Zhong and Xia, 2021). Adrb2 negatively impacts bone formation by inhibiting the cAMP/CREB pathway, which is important in osteoblast activity (Dimitri and Rosen, 2017; Zhong and Xia, 2021). It has also been shown that parathyroid hormone (PTH) suppresses Adrb2 expression in the MC3T3-E1 osteoblastic cell line mediating phosphorylation of CREB (Moriya et al., 2015). In bone cells, the major pathway involved in PTH is via cAMP/PKA/CREB where the phosphorylation of CREB upregulates osteoblasts differentiation and bone formation (Zhang et al., 2011). Moreover, in animal studies, Adrb2-deficient mice have shown higher osteoblast activity and high bone mass with decreased resorption (Takeda et al., 2002; Elefteriou et al., 2005; Zhong and Xia, 2021). Other studies have also shown that blocking Adrb2 increases bone mass (Pasco et al., 2004; Sato et al., 2010).
Receptors Adra1b and Adra1d was strongly expressed in MC3T3-E1 cells (Nishiura and Abe, 2007). Likewise, Adra1b and Adrb2 were detected in human osteoblasts and MG63 cells, whereas expression of Adra2a was relatively weak, with low levels of expression of Adra2c only in demonstrated in MG63 cells (Huang et al., 2009). This study also showed the expression of Adra1b to be three-fold higher than Adrb2 expression in human osteoblasts. It has been reported the expression of Adrb1, Adrb2, Adrb3 and Adra1b receptors in human periosteum-derived osteoblastic cells (SaM1) and human osteosarcoma-derived cells: SaOS2, HOS and MG63 (Togari et al., 1997; Togari, 2002).
In human osteoblasts, both Adra1 and Adrb2 agonists stimulate RANKL and soluble decoy RANK receptor osteoprotegerin (OPG) mRNA expression, where Adra1 agonists stimulated proliferation and OPG mRNA expression at lower concentration than required for RANKL stimulation. The Adrb2 agonists predominantly upregulated resorption signals by increasing RANKL expression at lower concentration than required for OPG stimulation. Moreover, beta blockers inhibited Adrb2 induced RANKL mRNA expression, but not OPG mRNA expression (Huang et al., 2009), which could result in higher BMD and protection against fracture, which has also been shown by Pasco et al. (2004) (Pasco et al., 2004). This suggests the possibility that Adra1 and Adrb2 may be intimately involved in regulating bone turnover. It has also suggested that the PKC and ERK signalling pathways may mediate RANKL expression via Adra1 receptors (Huang et al., 2009).
There is a lack of data on adrenergic receptor expression profile in bone cells, particularly in primary human cells, where adrenergic receptors, especially Adrb2, are associated with bone mass regulation. In summary, both osteoclasts and osteoblasts express adrenergic receptors including Adra1b, Adra2a, Adra2b, Adra2c, Adra1d, Adrb1, Adrb2 and Adrb3 (Table 1). Among these receptors, Adrb2 is highly implicated in regulating bone mass, where Adrb2-signaling increases resorption in osteoclasts via prostaglandin 2 and IL-6, while decreasing bone formation via cAMP/PKA/CREB/AP1 signalling leading to overall decrease in bone mass (Kellenberger et al., 1998; Arai et al., 2003; Zhong and Xia, 2021). Moreover, Adra1 also regulates osteoclast resorption via PKC/ERK pathway (Huang et al., 2009).
8 ConclusionIn light of the emerging clinical evidence that schizophrenia and antipsychotics both independently decrease BMD and increase fracture risk, there remains a need to build on the existing evidence identifying the presence of dopamine, serotonin and adrenergic receptors in bone. The evidence to date indicate that dopamine directly inhibits osteoclastogenesis via the D2/cAMP/PKA/CREB pathway (Wang et al., 2021b). In contrast, dopamine stimulates osteoblastogenesis via the D1/ERK 1 and 2/Runx2 pathway while inhibition of cAMP/PKA can inhibit dopamine-induced osteogenesis (Wang et al., 2020). Centrally-produced serotonin decreases bone resorption and increases bone formation. However, in the periphery, serotonin directly acts on osteoblasts to inhibit bone formation (Ducy and Karsenty, 2010). Data indicate that 5HT2B and 5HT6 play an important role in maintaining bone mass. It has been suggested that bone turnover is regulated by Adrb2 and Adra1 by regulating RANKL and OPG levels (Huang et al., 2009). Further, activation of Adrb2 inhibits CREB phosphorylation leading to decreased osteoblast proliferation and Adrb2 can activate cAMP/PKA/AP1 pathways to regulate bone formation (Kellenberger et al., 1998; Arai et al., 2003; Zhong and Xia, 2021).
Therefore, we assume that antipsychotics may cause imbalance in bone formation and resorption via dopamine, serotonin and adrenergic receptors through cAMP/PKA/CREB, ERK 1/2, AP1 and Jab1/Smad1, 5, 8/BMP2 pathways. Further studies are required in animal and human models of bone formation and resorption to help characterise the role of associated signalling pathways and the direct impact of antipsychotics on different receptors present in bone cells. Given antipsychotics largely target dopamine and serotonin receptors, as well as, albeit to a lesser extent adrenergic receptors, it is important to explore the mechanism(s) of action by which antipsychotics impact bone loss.
Author contributionsDKW drafted this manuscript. All authors contributed to the article and approved the submitted version.
FundingThis work was supported by the National Health and Medical Research Council (NHMRC), Australia (Grant Nos 1162867). DKW and BAM were supported by the Deakin University Postgraduate Research Scholarship (DUPRS) and LW by the NHMRC Emerging Leadership Fellowship (Grant No. 1174060).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
ReferencesAbbas, A. I., Hedlund, P. B., Huang, X. P., Tran, T. B., Meltzer, H. Y., and Roth, B. L. (2009). Amisulpride is a potent 5-HT7 antagonist: Relevance for antidepressant actions in vivo. Psychopharmacol. Berl. 205 (1), 119–128. doi:10.1007/s00213-009-1521-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Al Shirawi, M. I., Edgar, N. E., and Kennedy, S. H. (2017). Brexpiprazole in the treatment of major depressive disorder. Clin. Med. Insights Ther. 9, 1179559X1773180. doi:10.1177/1179559x17731801
CrossRef Full Text | Google Scholar
Arai, M., Nagasawa, T., Koshihara, Y., Yamamoto, S., and Togari, A. (2003). Effects of β-adrenergic agonists on bone-resorbing activity in human osteoclast-like cells. Biochimica Biophysica Acta (BBA) - Mol. Cell Res. 1640 (2), 137–142. doi:10.1016/s0167-4889(03)00042-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Aringhieri, S., Carli, M., Kolachalam, S., Verdesca, V., Cini, E., Rossi, M., et al. (2018). Molecular targets of atypical antipsychotics: From mechanism of action to clinical differences. Pharmacol. Ther. 192, 20–41. doi:10.1016/j.pharmthera.2018.06.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Arnaiz, J. A., Rodrigues-Silva, C., Mezquida, G., Amoretti, S., Cuesta, M. J., Fraguas, D., et al. (2021). The usefulness of Olanzapine plasma concentrations in monitoring treatment efficacy and metabolic disturbances in first-episode psychosis. Psychopharmacol. Berl. 238 (3), 665–676. doi:10.1007/s00213-020-05715-5
CrossRef Full Text | Google Scholar
Asano, T., Tanaka, K.-I., Tada, A., Shimamura, H., Tanaka, R., Maruoka, H., et al. (2017). Ameliorative effect of chlorpromazine hydrochloride on visceral hypersensitivity in rats: Possible involvement of 5-ht2a receptor. Br. J. Pharmacol. 174 (19), 3370–3381. doi:10.1111/bph.13960
PubMed Abstract | CrossRef Full Text | Google Scholar
Azimi Manavi, B., Stuart, A. L., Pasco, J. A., Hodge, J. M., Samarasinghe, R. M., Weerasinghe, D. K., et al. (2023). Use of antipsychotic medication and its relationship with bone mineral density: A population-based study of men and women. Front. Psychiatry 13, 13. doi:10.3389/fpsyt.2022.1004366
CrossRef Full Text | Google Scholar
Ban, T. A. (2007). Fifty years chlorpromazine: A historical perspective. Neuropsychiatr. Dis. Treat. 3 (4), 495–500.
PubMed Abstract | Google Scholar
Battaglino, R., Fu, J., Späte, U., Ersoy, U., Joe, M., Sedaghat, L., et al. (2004). Serotonin regulates osteoclast differentiation through its transporter. J. Bone Mineral Res. 19 (9), 1420–1431. doi:10.1359/JBMR.040606
PubMed Abstract | CrossRef Full Text | Google Scholar
Baudry, A., Bitard, J., Mouillet-Richard, S., Locker, M., Poliard, A., Launay, J.-M., et al. (2010). Serotonergic 5-HT2B receptor controls tissue-nonspecific alkaline phosphatase activity in osteoblasts via eicosanoids and phosphatidylinositol-specific phospholipase C. J. Biol. Chem. 285 (34), 26066–26073. doi:10.1074/jbc.M109.073791
PubMed Abstract | CrossRef Full Text | Google Scholar
Becker, D., Liver, O., Mester, R., Rapoport, M., Weizman, A., and Weiss, M. (2003). Risperidone, but not olanzapine, decreases bone mineral density in female premenopausal schizophrenia patients. J. Clin. Psychiatry 64 (7), 761–766. doi:10.4088/jcp.v64n0704
PubMed Abstract | CrossRef Full Text | Google Scholar
Bliziotes, M., Eshleman, A., Burt-Pichat, B., Zhang, X. W., Hashimoto, J., Wiren, K., et al. (2006). Serotonin transporter and receptor expression in osteocytic MLO-Y4 cells. Bone 39 (6), 1313–1321. doi:10.1016/j.bone.2006.06.009
PubMed Abstract | CrossRef Full Text | Google Scholar
Bliziotes, M. M., Eshleman, A. J., Zhang, X. W., and Wiren, K. M. (2001). Neurotransmitter action in osteoblasts: Expression of a functional system for serotonin receptor activation and reuptake. Bone 29 (5), 477–486. doi:10.1016/s8756-3282(01)00593-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Boer, M., Castelein, S., Wiersma, D., Schoevers, R., and Knegtering, R. (2015). The facts about sexual (Dys)function in schizophrenia: An overview of clinically relevant findings. Schizophr. Bull. 41, 674–686. doi:10.1093/schbul/sbv001
PubMed Abstract | CrossRef Full Text | Google Scholar
Bolton, J. M., Morin, S. N., Majumdar, S. R., Sareen, J., Lix, L. M., Johansson, H., et al. (2017). Association of mental disorders and related medication use with risk for major osteoporotic fractures. JAMA Psychiatry 74 (6), 641–648. doi:10.1001/jamapsychiatry.2017.0449
留言 (0)