
記住我
Atherosclerotic cardiovascular diseases (ASCVD) have emerged as the most common burden of disease as a result of the aging and expanding global population (Mensah et al., 2019). As the pathology of ASCVD, atherosclerosis (AS) generates a continuous buildup of vessel-occluding plaques in the subendothelial intimal layer of coronary arteries, eventually leading to considerable blood flow restriction and essential tissue hypoxia (Libby, 2002; Gallino et al., 2014). Most cardiovascular events are caused by the rupture of atherosclerotic plaques in the arterial artery wall and the subsequent formation of an occluding thrombus.
In addition to the deposition and retention of modified lipoproteins and the buildup of immune cells in the walls of major arteries, AS is characterized by a low-grade, persistent, chronic inflammation of the arterial wall (Edgar et al., 2021). All phases of AS are mostly attributed to monocytes and monocyte-derived macrophages, which are also thought to be responsible for persistent chronic inflammation (Moore et al., 2013). The traditional view is that innate immune cells, such as macrophages, can only eliminate pathogens non-specifically through biological processes such as phagocytes (Bonilla and Oettgen, 2010). However, a growing body of research suggests that monocytes/macrophages may also develop memory capabilities similar to those of the adaptive immune system after exposure to pathogens (Arts et al., 2018). Myeloid cells of the innate immune system become more sensitive after activation with the same or different stimuli to produce a persistent inflammatory monocyte/macrophage phenotype, a phenomenon known as “trained immunity” or “innate immunological memory” (Netea et al., 2020). This persistent overactivation of the innate immune system could contribute to the incessant vascular wall inflammation that is characteristic of AS (Moore et al., 2013).
For thousands of years, herbal medicines have been widely utilized alone or as a supplementary strategy to treat various disorders in East Asia because of their reduced toxicity, fewer side effects, and cheaper cost (Wang et al., 2018). Along with the development of these natural therapies, herbal medicine is becoming more widely accepted as a supplement and alternative therapy in many countries (Liang et al., 2021). According to the most recent statistics on US-FDA (United States Food and Drug Administration) authorized drugs, herbal remedies have been a vital source of novel medications (Newman and Cragg, 2020). A growing body of scientific evidence has revealed that natural medicines and phytochemicals from natural herbal medicines exhibit promising anti-AS properties (Zhang et al., 2021a). Based on the regulation of targeting trained immunity in monocyte and macrophage, natural compounds generated from herbal remedies are surely excellent resources for selecting potential therapeutics to treat AS.
This review aims to provide more information on the role of trained immunity in the pathophysiology of AS, which might be a potential pharmacological target of natural products.
2 Trained immunity in ASConventional wisdom generally considers the adaptive immune system as a specific protective mechanism that can form more specialized lines of defense against re-infection with the same pathogens (Domínguez-Andrés et al., 2019). However, innate immune cells (e.g., macrophages/monocytes) have been reported to display similar immune memory, referred to as trained immunity (Conrath et al., 2015; Milutinović and Kurtz, 2016; Gourbal et al., 2018). Studies of gene-specific chromatin changes brought about by lipopolysaccharide (LPS) have first shown trained immunity characteristics of monocytes/macrophages (Foster et al., 2007). Subsequently, infectious stimuli, such as β-glucan and Bacille Calmette–Guérin (BCG), improved their reactivity to stimulation with unrelated infections or molecular patterns linked with those pathogens (Quintin et al., 2012; Saeed et al., 2014). Factors that contribute to the development of AS, such as uric acid and oxidized low-density lipoprotein (oxLDL), and other endogenous ligands can activate trained immunity (Bekkering et al., 2014; Crişan et al., 2017). Freshly isolated monocytes from patients who had symptoms of coronary artery disease (CAD) had a higher capacity to produce cytokines than those from healthy controls, and this capacity was maintained following ex vivo conversion to macrophages for 5 days (Shirai et al., 2016). The atherogenic factors are characterized by increased production of proatherogenic cytokines and chemokines like tumor necrosis factor-α (TNF-α), IL-6, monocyte chemoattractant protein-1 (MCP-1), matrix metalloproteinases 2 (MMP-2), and MMP-9 and increased foam cell formation is indeed demonstrated by large-scale phenotyping of trained macrophages in vitro (Bekkering et al., 2014).
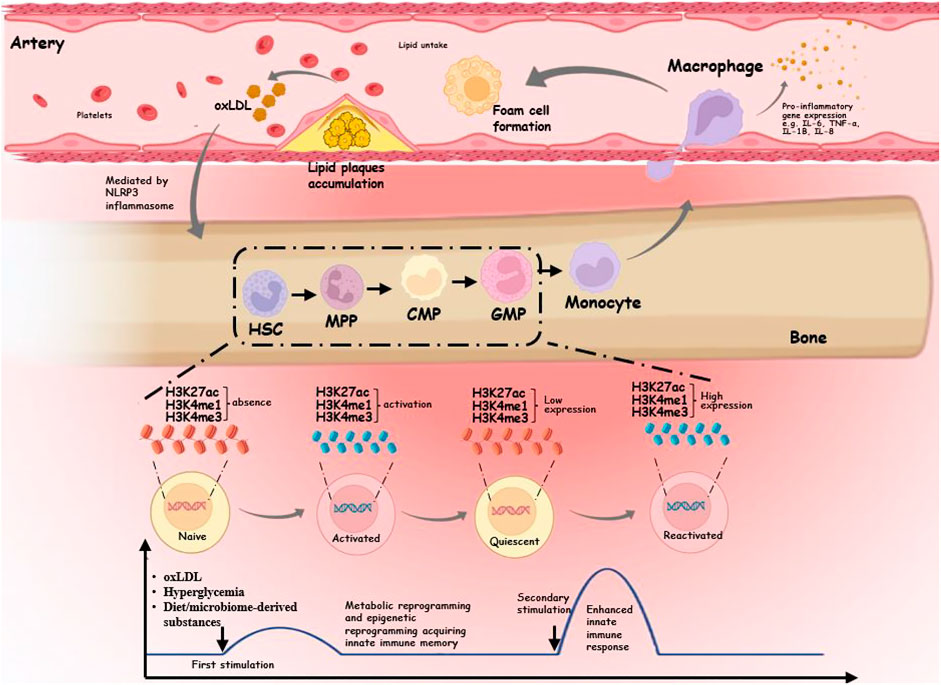
After short activation with endogenous ligands, a persistent proinflammatory phenotype can emerge in AS monocytes/macrophages (Leentjens et al., 2018). The three key components of trained immunity are metabolic reprogramming, epigenetic reprogramming, and the promotion of myelopoiesis progenitors (Fanucchi et al., 2021) (Figure 1). First, metabolic reprogramming is responsible for the induction, maintenance, and regulation of trained immunity. Different metabolic pathways supply the required substrates for altering the structure of the respective sections of the chromatin and genome, in addition to acting as a source of energy and building components for the dynamic remodeling of the epigenetic landscape. Second, epigenetic reprogramming ultimately links metabolic changes to a cell’s gene expression and inflammatory phenotype. Finally, trained immunity in bone marrow by hematopoietic stem cells (HSCs) maintains long-term effects on circulating monocytes through differentiation into progenitor and mature cells (Mitroulis et al., 2018). In addition to atherosclerotic triggers, such as lipoproteins, glucose, diet, and microbiota-derived substances, proinflammatory cytokines secreted by monocytes/macrophages with an inflammatory phenotype may alter the tissue microenvironment by altering macrophages, the functional state of cells, leading to a vicious circle (Groh et al., 2018).
FIGURE 1. Schematic diagram of the trained immunity mechanism in atherosclerotic cardiovascular disease. In the hematopoietic system, myeloid cells exposed to endogenous triggers undergo epigenetic and metabolic reprogramming, resulting in acquiring innate immune memory. The initial gene activation is accompanied by the accumulation of H3K4me3 on the gene promoter, and the persistence of H3K4me1 or H3K27ac in secondary stimulation leads to an enhanced innate immune response. These trained myeloid cells differentiate into monocytes, which travel further into the intima to become macrophages. Trained macrophages produce high levels of proinflammatory cytokines such as TNF-α, IL-6, IL-8, and IL-18 and uptake of lipids to form foam cells. When plaques form, endogenous stimuli may be further released to form trained immunity mediated by the NLRP3 inflammasome.
2.1 Metabolic reprogramming of trained immunity in ASThe trained immune activation has to quickly access a supply of substrates to initiate the numerous metabolic processes associated with the immune response. Intracellular metabolic pathways of glucose, amino acids, lipids, and nucleic acids are altered in response to trained immune activation (Fanucchi et al., 2021). When normal cells are at rest, they obtain enough energy via metabolic processes that are extremely effective but rather slow, such as oxidative phosphorylation (OXPHOS) and fatty acid oxidation (FAO) (Augert et al., 2020). In contrast, trained immune cells continue to opt for “aerobic glycolysis,” which uses glycolysis instead of OXPHOS to generate energy under normoxic conditions, similar to the “Warburg effect” in cancer (Mills et al., 2016; Renner et al., 2017). In addition to glucose metabolism, trained immune cells exhibit altered lipid and amino acid metabolic patterns. For instance, the Krebs cycle’s anabolic redefinition to synthesize cholesterol and phospholipids from citrate and acetyl coenzyme A (CoA) is a crucial metabolic event in trained monocytes (Arts et al., 2016a). When exposed to β-glucan, cholesterol synthesis is increased, but fluvastatin, an inhibitor of the enzyme 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase, inhibits trained immunity by downregulating histone H3 lysine 4 trimethylation (H3K4me3) and limiting the production of proinflammatory cytokines (Bekkering et al., 2018). For the progression of AS, the control of cholesterol import and efflux is critical. Similar to glutamine, arginine, and glycine, several particular amino acids are overexpressed in atherosclerotic plaques and have AS-promoting effects (Mallat et al., 1999; Sheehan et al., 2011). The intermediate metabolites from many metabolic pathways, such as aerobic glycolysis, glutaminolysis, cholesterol metabolism, and fatty acid synthesis, not only are a source of energy for the cell but also play several significant biological roles (Groh et al., 2018). Additionally, some chemo drugs made from natural herbal products, such as resveratrol and epigallocatechin gallate, can prevent cells from reprogramming their metabolism in response to AS.
2.1.1 Glucose metabolism and ASAlthough OXPHOS produces ATP more efficiently than other cellular processes (approximately 30 ATP molecules can be produced per glucose molecule during OXPHOS, whereas glycolysis can only produce two ATP molecules per glucose molecule) (Tabas and Bornfeldt, 2020). However, glycolysis produces ATP faster than OXPHOS, allowing immune cells to respond quickly to stimuli (Tabas and Bornfeldt, 2020). A clinical trial found an enhanced capacity for cytokine production in circulating monocytes obtained from ASCVD patients, which was associated with the upregulation of glycolytic enzymes (Bekkering et al., 2016; Shirai et al., 2016). This phenotype continued even after in vitro macrophage differentiation, displaying a greater glycolytic flux and a higher oxygen consumption rate (Shirai et al., 2016). Furthermore, the inhibition of specific tricarboxylic acid (TCA) cycle steps that support inflammatory processes is associated with increased glycolysis in inflammatory macrophages that primarily produce and release proinflammatory mediators, such as interleukin-1 (IL-1), TNF-α, chemokine C–C motif ligand 2 (CCL2), IL-12, and nitric oxide (NO), through inducible nitric oxide synthase (iNOS). The key proteins involved in glycolysis are introduced in the following sections.
2.1.1.1 GLUT1Glucose transporter 1 (GLUT1; gene name SLC2A1) on the cell membrane initiates glucose uptake by monocytes/macrophages (Fukuzumi et al., 1996). LPS and oxLDL, which cause inflammation, can boost GLUT1 expression and thereby increase glucose influx. Hexokinase phosphorylates glucose inside the cell to produce glucose-6-phosphate, which is then utilized in the pentose phosphate pathway (PPP), fatty acid synthase (FAS), or glycolysis. When glucose is processed in the cytosol by glycolysis, two ATPs and pyruvates are produced. Pyruvate produced during glycolysis either enters the mitochondrial TCA cycle or is transformed to lactate by lactate dehydrogenase (Christofk et al., 2008). In plaque macrophages, GLUT1 can promote antiatherosclerotic activities. The efferocytosis procedure increases the expression of GLUT1, which promotes an increase in glucose absorption and a transition from OXPHOS to improved aerobic glycolysis, both of which are required for the effective clearance of apoptotic cells (Morioka et al., 2018). When myeloid-targeted LysM-Cre Slc2a1fl/fl animals were transplanted into Ldlr−/− mice on a Western-style diet (WTD), the amount of necrotic core in the aorta increased (Morioka et al., 2018). Another study revealed that GLUT1 deletion in hematopoietic cells inhibited myelopoiesis, monocyte recruitment to lesions, and the progression of AS in ApoE−/− mice, indicating that the main role of GLUT1 in this model was to encourage the proliferation of bone marrow HSCs and multi-potential progenitors, as well as the commitment of these cells to the bone marrow (Sarrazy et al., 2016).
2.1.1.2 HIF-1αHyperoxia-inducible substance 1α (HIF-1α) is activated in hypoxic circumstances, allowing cells to switch to glycolysis and create ATP when oxygen is limited. Low oxygen levels trigger the HIF-1α transcription factor to initiate glycolytic metabolism, which decreases the need for OXPHOS and increases the expression of the key glycolysis proteins GLUT1, hexokinase II (HK-II), and 6-phosphofructo-2-kinase/fructo-2, 6-bisphosphatase (PFKFB3), which increases glycolytic flux (Tawakol et al., 2015). The activated macrophages will emit a lot of cytokines and absorb a lot of glucose. Indeed, hypoxia, HIF-1α expression, and FDG (fluorodeoxyglucose) uptake in macrophages are associated with atherosclerotic plaques in animal models of AS (Folco et al., 2011; Tawakol et al., 2015; Aarup et al., 2016).
2.1.2 Lipid metabolism and ASIn homeostasis, lipoproteins taken up by macrophages are transported to lysosomes for the hydrolysis of cholesteryl esters. Free cholesterol is transported to the cytoplasm, where it is transported to the cell membrane for export or transported to the ER for re-esterification and storage in lipid droplets (LDs). Macrophages in advanced plaques provide signs of huge accumulations of free cholesterol, which suggests a breakdown in the mechanisms that keep cholesterol levels in balance. Membrane damage and metabolic dysregulation in the ER and mitochondria are required to maintain macrophage cholesterol homeostasis and reduce inflammation, which results from excessive accumulation of free cholesterol. Furthermore, high levels of modified cholesterol, oxLDL, are taken up by macrophages to form foam cells and promote plaque by secreting numerous proinflammatory cytokines and chemokines and producing MMPs that degrade plaque extracellular matrix pathogenesis (Khokha et al., 2013; Tall and Yvan-Charvet, 2015). It is reported that the induction of trained immunity in monocytes required stimulation of the cholesterol biosynthesis pathway but not cholesterol synthesis itself. oxLDL is an endogenous ligand that triggers trained immunity to activate monocytes/macrophages (Bekkering et al., 2014; Crişan et al., 2017). Another crucial characteristic of monocytes trained on β-glucan is increased cholesterol production (Netea et al., 2020). In primary human monocytes, fluvastatin, an inhibitor of HMG-CoA reductase, inhibits trained immunity (Arts et al., 2016a). Notably, β-glucan-induced training of mature myeloid cells and their progenitors requires enhanced cholesterol production. The accumulation of cholesterol esters and lipids with more saturated acyl chains is associated with the long-term myelopoiesis bias that β-glucan induced training imparts to HSCs (Mitroulis et al., 2018). The HSC population increase and myelopoiesis caused by β-glucan are reduced by HMG-CoA reductase inhibitor (Mitroulis et al., 2018).
2.1.3 Amino acid metabolism and ASUnder AS pathological conditions, amino acid metabolites play an important role in supporting the induction, maintenance, and regulation mechanisms of trained immunity (Napoli et al., 2006). Therefore, it is necessary to decipher the role of specific amino acid metabolites in the induction of trained immunity.
2.1.3.1 GlutamineGlutamate is one of the amino acids that has been well-studied for its role in controlling inflammation (Wallace and Keast, 1992). By directly converting into glutamate, α-ketoglutarate, and succinate semialdehyde, glutamine contributes to the TCA cycle (Jha et al., 2015). Additionally, glutamate can be employed as a source of citrate for the FAS-catalyzed production of fatty acids. Recent research has demonstrated that glutaminolysis is increased in trained macrophages and is essential for the establishment of a trained macrophage phenotype in response to β-glucan (Arts et al., 2016a). In a different research study, oxidized phospholipids made of 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine (oxPAPC) were exposed to macrophages, which led to the development of AS, glutaminolysis, and IL-1 48. This study demonstrated that in contrast to macrophages activated with LPS alone, those exposed to oxPAPC and LPS together had a metabolic change (Di Gioia et al., 2020). This metabolic shift was characterized by increased mitochondrial respiration, glutaminolysis, and accumulation of oxaloacetate, which stabilized HIF-1α and increased IL-1β production. IL-1β immunoreactivity in CD68+ lesional cells decreased in mice with systemic suppression of this pathway, which also reduced early AS (Di Gioia et al., 2020). Furthermore, the TCA cycle’s glutamine replenishment causes fumarate to accumulate, which integrates immunological and metabolic circuits to cause monocyte epigenetic reprogramming by inhibiting KDM5 activity and boosting the methylation of histone lysine 4 residues (Arts et al., 2016a). An epigenetic program identical to trained immunity-mediated by β-glucan was induced by fumarate. To support this, glutaminolysis inhibition and cholesterol production suppression in mice decreased the induction of trained immunity by β-glucan (Arts et al., 2016a).
2.1.3.2 ArginineIn the context of AS pathology, arginine metabolism and its by-product NO are critical for the early stages of the disease (Lv et al., 2021). iNOS is ubiquitously expressed in activated and developing macrophages. NO creation by arginine is probably a factor in the metabolic transition. NO has been reported to inhibit OXPHOS in activated dendritic cells and inflammatory macrophages downstream of iNOS (Everts et al., 2012; Van den Bossche et al., 2016). Conversely, in alternatively activated macrophages and in the macrophages of atherosclerotic lesions that are regressing, Arg1 converts arginine to putrescine (Willecke et al., 2015). Under specific conditions, these two arginine metabolic routes can inhibit one another. As a result, NO inhibits ornithine decarboxylase’s ability to catalyze the conversion of ornithine to putrescine by S-nitrosylation a cysteine that is essential for the enzyme’s ability to function (Bauer et al., 2001). Conversely, ornithine decarboxylase prevents macrophages from becoming activated in an inflammatory response (Hardbower et al., 2017). Arginine metabolism has been reprogrammed, which promotes proinflammatory and healing processes.
2.1.3.3 SerineRecent research has demonstrated that LPS-activated macrophages promote serine synthesis, PPP, and one-carbon metabolism, which synergistically drive epigenetic reprogramming of IL-1β expression. The production of S-adenosylmethionine (SAM) during LPS-induced inflammation is fueled by the synergistic integration of glucose-derived ribose and one-carbon units supplied by glucose and serine metabolism into the methionine cycle through de novo ATP synthesis. Impairment of these metabolic pathways that feed SAM generation leads to anti-inflammatory outcomes (Yu et al., 2019). According to a different research study, serine is necessary for the synthesis of glutathione and IL-1β through the action of glycine (Rodriguez et al., 2019).
2.2 Epigenetic reprogramming of trained immunity in ASRegulating gene expression without changing the DNA sequence itself is referred to as epigenetic reprogramming. Epigenetic reprogramming enables innate immune cells to react to future stimuli with a stronger, quicker, or qualitatively different transcriptional response (Zarzour et al., 2019). Epigenetic regulatory mechanisms encompass diverse molecular processes, including histone post-translational modifications, DNA methylation, and long non-coding RNAs (lncRNAs).
2.2.1 Histone modificationsEpigenetic reprogramming occurs primarily through histone changes at the level of the chromatin structure to promote a sustained enhanced functional state of trained innate immune cells (Saeed et al., 2014). Neutralization of the positive charge of lysine residues in histones by histone acetylation increases the binding of transcription factors activating gene transcription (Bannister and Kouzarides, 2011). The specific lysine residue implicated and the sum of the additional methyl groups determine the effect of histone methylation on gene transcription. Two important epigenetic marks for trained immunity are as follows: the H3K4me3 accumulation at the gene promoter and the histone 3 lysine 27 acetylation (H3K27ac) acquisition at the gene’s distal enhancer (generated by histone 3 lysine acid 4 methylation (H3K4me1)) (Netea et al., 2020).
2.2.2 DNA methylationDNA methylation is involved in the regulation of patterns of gene expression. DNA methyltransferases (DNMTs) use CpG-rich regions as recognition cues to methylate cytosines (m5C), which suppresses transcription. Proteins with histone-binding domains that can “read,” “write,” or “erase” histone marks may detect the tails that protrude from histone octamers (Fanucchi et al., 2021). In addition to methylation, acetylation, phosphorylation, and ubiquitination, these enzymes can catalyze the addition or removal of a broad and diverse range of other histone modifications. Different DNA methylation patterns discriminate between “responders” (those who can experience taught immunity) and “non-responders” to stimuli, such as BCG, that produce trained immunity (Verma et al., 2017). Forty-three genes had distinct methylation patterns in BCG-naive responders as opposed to non-responders in a follow-up investigation, which may be utilized to predict sensitivity to triggers of trained immunity (Das et al., 2019). Numerous studies have shown that various DNA and histone modification combinations affect whether DNA is kept in an accessible or “open” state or an inaccessible or “closed” one (Fanucchi et al., 2021). To enable quick and effective transcriptional activation, highly accessible DNA is quickly bound by the transcriptional machinery and transcription factors. This establishes a clear connection between the transcriptional state of protein-coding genes and the “openness” of DNA.
2.2.3 lncRNAsDuring trained immunity, lncRNA-dependent regulation has a significant impact on the epigenetic reprogramming of immune genes (Fanucchi et al., 2021). Several lncRNAs known as immune-gene priming lncRNAs (IPLs) were found by the application of a bioinformatic pipeline that comprised 3D nuclear architecture, lncRNA and enhancer expression data, and the epigenetic status of immune genes at the genome scale (Fanucchi et al., 2019). The WD repeat-containing protein 5 (WDR5)-mixed lineage leukemia protein 1 complex is directed across the chemokine promoters by UMLILO (upstream master lncRNA of the inflammatory chemokine locus), allowing H3K4me3 epigenetic priming, according to careful analysis of a prototypical IPL known as UMLILO (Fanucchi et al., 2019). Several trained immune genes share this mechanism. Training mediated by β-glucan upregulates IPLs in a way that depends on the nuclear factor of activated T cells, which epigenetically reprograms immune genes. The Cxcl genes are not trained, and the murine chemokine topologically associating domain is devoid of an IPL. Cxcl genes are trained as a result of the insertion of UMLILO into the chemokine topologically associating domain in murine macrophages (Fanucchi et al., 2019). This offers compelling evidence that the development of trained immunity depends on lncRNA-mediated control. Further research is required to examine these pathways in various experimental contexts, as recent studies have only examined the function of IPLs in β-glucan-induced trained immune characteristics.
2.3 Modulation of myelopoiesis progenitorsThe observation of trained circulating monocytes months after BCG vaccination suggests that adaptive processes induced by trained immunity involve alterations in hematopoietic progenitors at the bone marrow level (Kleinnijenhuis et al., 2012). Evidence shows that trained immunity plays a role at the bone marrow level in the context of AS. In mice, the administration of β-glucan leads to long-term transcriptional and metabolic alterations in hematopoietic stem and progenitor cells, resulting in their expansion and bias toward myelopoiesis. This enhances their ability to respond to secondary LPS stimulation and protects them from chemotherapy-induced myelosuppression (Mitroulis et al., 2018). The shared β-subunit of the IL-3/GM-CSF receptor, CD131, is linked with enhanced surface expression in this long-term reprogramming. In a mouse model of predisposition to AS, a similar process occurs when hypercholesterolemia induces enhanced myeloid proliferation and inflammation, suggesting a possible role for trained immunity in the context of traditional cardiovascular risk factors (Wang et al., 2014). Existing evidence also supports a link between enhanced glycolysis in myeloid cells and AS. In hypercholesterolemic ApoE−/− mice, leucocytes and HSPCs show enhanced GLUT1-dependent glucose absorption, which is linked to an elevated mitochondrial potential, providing evidence for a role for myeloid cell glycolysis in myelopoiesis and atherogenesis. This suggests that the mitochondria in these cells are fed by the inflow of glycolytic metabolites for OXPHOS and ATP synthesis (Sarrazy et al., 2016).
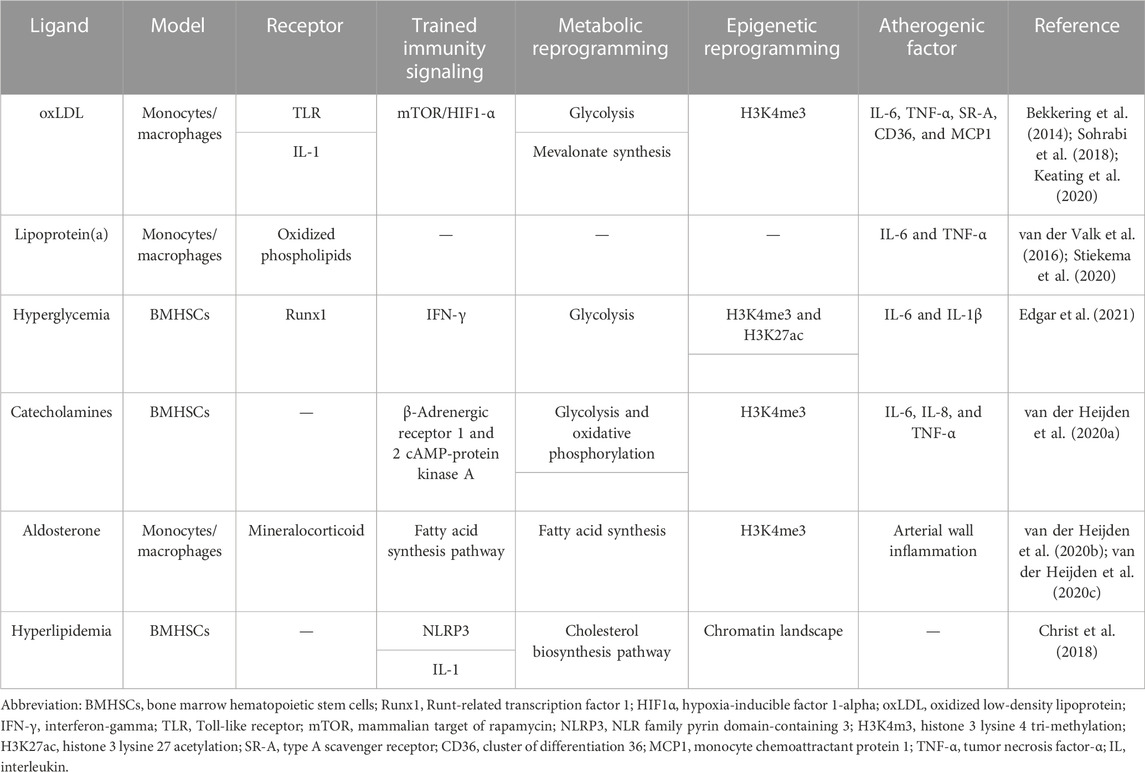
3 Endogenous triggers of trained immunityIn addition to microbial sources, endogenous molecules, such as cellular metabolites oxLDL, lipoprotein(a), and hyperglycemia, can induce trained immunity (Bekkering et al., 2014; van der Valk et al., 2016; Braza et al., 2018; Edgar et al., 2021). These endogenous triggers play a role in the development of ASCVD (Flores-Gomez et al., 2021). We will discuss the link between these endogenous triggers of trained immunity and atherosclerotic plaque formation in activated monocyte–macrophages (Table 1).
TABLE 1. Endogenous triggers of trained immunity.
3.1 oxLDLoxLDL is a modified lipoprotein and is one of the key atherogenic molecules within plaques that activates immune cells (Moore and Tabas, 2011). oxLDL-trained macrophages exhibit significant metabolic and epigenetic rewiring, similar to BCG and β-glucan. The mammalian target of the rapamycin (mTOR)/HIF1-α signaling pathway is necessary for the upregulation of glycolysis and OXPHOS in oxLDL-induced cells (Keating et al., 2020). The increase in glycolysis and the proinflammatory phenotype in macrophages were avoided by pharmacological suppression of the mTOR pathway and the signaling molecules involved and by inhibiting glycolysis with 2-deoxyglucose (Sohrabi et al., 2018). Epigenetic reprogramming is another characteristic of oxLDL-trained macrophages. OxLDL interacts with the myeloid cell surface receptor cluster of differentiation 36 (CD36) as a damage-associated molecular pattern (DAMP) (Moore et al., 2013). The internalization and release of oxLDL into the cytoplasm may create cholesterol crystals, which activates the NOD-, LRR-, and pyrin domain-containing protein 3 (NLRP3) inflammasome and releases IL-1β and other proinflammatory cytokines, as well as a protracted inflammatory response (Sheedy et al., 2013). Promoters of genes encoding proinflammatory and proatherogenic cytokines and chemokines, such as IL-6, TNF-α, type A scavenger receptor (SR-A), and CD36, are more likely to have the activating histone modification H3K4me3 20. OxLDL training was fully blocked by pharmacologically inhibiting histone methyltransferases, demonstrating that epigenetic alterations are what actually trained immunity by oxLDL (Bekkering et al., 2014).
3.2 Lipoprotein(a)Lipoprotein(a) is the main circulating carrier of oxidized phospholipids, which plays an important role in atherogenesis (Boffa and Koschinsky, 2019). Monocytes from healthy donors exposed for 24 h to high lipoprotein(a) extracted from hyperlipidemic patients produced more proinflammatory cytokines during the subsequent 6 days compared to controls. Anti-oxidized phospholipid antibodies reduced the training of monocyte-derived macrophages, demonstrating that oxidized phospholipids are the mediating factor in this process (van der Valk et al., 2016). After Pam3Cys and LPS ex vivo stimulation, monocytes showed an increased ability to generate proinflammatory cytokines, including IL-6 and TNF-α (van der Valk et al., 2016). A recent study has shown that individuals with cardiovascular diseases (CVD) may have their proinflammatory monocyte activation reversed by significantly reducing their lipoprotein(a) levels, demonstrating that at least some of this proinflammatory impact is reversible (Stiekema et al., 2020).
3.3 HyperglycemiaHyperglycemia, a cardinal feature of diabetes, exacerbates AS progression, delays plaque regression (Parathath et al., 2011), and increases proinflammatory gene expression and resistance to induction of M2-related gene expression (American Diabetes Association, 2015). Evidence suggests that hyperglycemia induces trained immunity in HSCs and macrophages, significantly exacerbating AS (Edgar et al., 2021). High extracellular glucose stimulated the production of proinflammatory genes and the functional properties that are proatherogenic in macrophages through pathways that depend on glycolysis. These traits were sustained by diabetic mouse bone marrow-derived macrophages even when they were cultivated in physiological glucose, showing hyperglycemia-induced trained immunity. A disease-relevant and enduring kind of trained innate immunity was demonstrated by an increase in aortic root AS following bone marrow transplantation from diabetic mice into (normoglycemic) Ldlr−/− mice. HSCs and macrophages generated from the bone marrow showed a proinflammatory priming effect in diabetes, according to integrated tests for transposase-accessible chromatin, chromatin immunoprecipitation, and RNA sequencing analysis (Edgar et al., 2021). Transcription factors, notably runt-related transcription factor 1 (Runx1), are implicated as mediators of trained immunity (Himes et al., 2005). These in vitro signs of trained immunity brought on by hyperglycemia were eliminated by pharmacological suppression of Runx1.
3.4 CatecholaminesIncreased sympathetic nervous system activity leads to proinflammatory leukocytosis in models of chronic psychological stress, stroke, and myocardial infarction (Dutta et al., 2012; Heidt et al., 2014; Courties et al., 2015). The pathways causing inflammatory alterations in disorders with high catecholamine levels can be explained by the fact that catecholamines cause long-lasting proinflammatory modifications in monocytes in vitro and in vivo, indicating well-trained immunity (van der Heijden et al., 2020a). After being restimulated with LPS 6 days later, monocyte-derived macrophages exposed to a relevant quantity of epinephrine/norepinephrine had higher levels of TNF-α and IL-6. Similar to oxLDL, this trained immune phenotype is connected to a higher glycolytic capability and OXPHOS. Studies using pharmacological inhibition demonstrated that the cAMP-protein kinase A pathway and the β-adrenergic receptors 1 and 2 are crucial for catecholamine-induced training (van der Heijden et al., 2020a). Patients who have pheochromocytoma and are regularly exposed to brief bursts of catecholamine production have this proinflammatory monocyte characteristic (Neumann and Young, 2019). Systemic inflammatory symptoms and an increased ex vivo cytokine response in activated monocytes were present in these individuals (Neumann and Young, 2019).
3.5 AldosteroneHuman macrophages deriving from monocytes have a long-lasting proinflammatory phenotype in vitro in response to transiently elevated aldosterone concentrations, which may be a factor in the atherosclerotic condition AS chronic inflammation of the artery wall (van der Heijden et al., 2020d). Aldosterone affects intracellular metabolism by increasing fatty acid synthesis, but it does not influence glycolysis and OXPHOS, as found in oxLDL training (van der Heijden et al., 2020c). Additionally, training by aldosterone is linked to the enrichment of H3K4me3 at the promoters of proinflammatory cytokines, including TNF-α and IL-6, demonstrating that aldosterone trains monocyte-derived macrophages in vitro. However, circulating monocytes are not more capable of producing cytokines in individuals with primary hyperaldosteronism. The macrophages of individuals with primary hyperaldosteronism only express more TNF-α following ex vivo differentiation into macrophages in autologous serum (van der Heijden et al., 2020b). These findings imply that aldosterone differs from the trained immune systems that have been well-established and elaborated by other stimuli.
3.6 HyperlipidemiaRecent research examined the possibility that a WTD, which is high in fats, sweets, and salt and lacks fiber, might lead to trained immunity (Christ et al., 2018). Circulating monocytes and their myeloid progenitors in AS-prone Ldlr−/− mice were significantly affected by proinflammatory transcriptional and epigenetic reprogramming from this WTD over 4 weeks. Increased inflammatory responses to subsequent innate immunological stimulation were brought on by the food intervention. Even when the mice were shifted to a typical chow diet for an additional 4 weeks, this trained immune phenotype was maintained despite circulating cholesterol levels and systemic inflammatory indicators reverting to normal (Christ et al., 2018).
4 Inhibitors of targeting trained immunityPharmacological inhibitors of histone methyltransferases and inhibitors of glycolysis, glutaminolysis, and the mevalonate pathway could restrain trained immunity. Following intraperitoneal injection of β-glucan, the effects of pharmacologically suppressing glutaminolysis and the pathway that produces mevalonate on the formation of trained immunity have been established in mouse models in vivo (Arts et al., 2016a). As a result, it would allow the development of innovative pharmaceutical methods to lower the risk of ASCVD and maybe lessen its negative consequences. In this section, we systematically summarize all reported drugs that suppress trained immunity based on multiple publications.
4.1 Agents that modulate metabolic reprogramming4.1.1 WortmanninThe fungus metabolite wortmannin was demonstrated to function as a selective inhibitor of AKT/phosphoinositide 3-kinase (PI3K) (Ui et al., 1995). The intermediate stimulation of the Akt/PI3K pathway is what causes mTOR to become active (Kelley et al., 1999). As stimulation with β-glucan caused a high phosphorylation of Akt, β-glucan was responsible for inducing this signal pathway in monocytes. Additionally, mTOR activation was inhibited as a result of Akt phosphorylation inhibition. Monocyte-trained immunity by β-glucan was suppressed by the Akt inhibitor wortmannin in a dose-dependent manner (Cheng et al., 2014).
4.1.2 RapamycinAccumulated mevalonate enhances the AKT-mTOR pathway during the establishment of trained immunity, which then triggers HIF1-α activation and a switch from OXPHOS to glycolysis. This response results in circulating monocytes with a trained immunity phenotype (Bekkering et al., 2018). The inhibition of mTOR with rapamycin prevents mevalonate-induced trained immunity. Additionally, BCG-induced trained immunity and β-glucan-induced trained immunity depend on the development of the histone marks H3K4me3 and H3K9me3, which are inhibited by the pharmacological regulation of rate-limiting glycolysis enzymes with rapamycin 30 88. Although rapamycins potently suppress trained immunity in vitro and T-cell proliferation in vivo, they exert little effect on innate immune cells (Braza et al., 2018).
4.1.3 AICArOne of the most widely utilized pharmacological AMP-activated protein kinase (AMPK) activity modulators is the nucleoside 5-aminoimidazole-4-carboxamide (AICAr). Early research on AMPK’s function in the physiological control of metabolism and the etiology of cancer was mostly centered on the use of AICAr as an AMPK activator (Višnjić et al., 2021). AICAr produces dose-dependent inhibition of β-glucan-induced trained immunity by indirectly inhibiting mTOR (Cheng et al., 2014).
4.1.4 MetforminMetformin is extensively used as a first-line therapy for type 2 diabetes and has a high safety profile (McCreight et al., 2016). Metformin acts through AMPK activation and subsequent mTOR inhibition. Metformin completely inhibits the protective effects of mice receiving metformin during and after primary infection with low-inoculum C. albicans, which increases survival during disseminated candidiasis brought on by a primary C. albicans injection. In vitro, metformin suppresses trained immunity induced by β-glucan (Cheng et al., 2014). Metformin also inhibits trained immunity by inhibiting the formation of histone marks, H3K4me3 and H3K9me3, by regulating the rate-limiting enzymes of glycolysis 30 88.
4.1.5 AscorbateAscorbate (vitamin C) is an essential micronutrient in primates and serves as an antioxidant and a cofactor for various enzymatic activities represented by prolyl hydroxylases (Fujii et al., 2022). Because the induction of glycolysis by mTOR is mediated by the activation of HIF1-α and stimulation of glycolytic enzymes and ascorbate inhibits HIF-1α expression, it inhibits training immune in a dose-dependent manner (Cheng et al., 2014).
4.1.6 ZVAD-fmkWestern diet feeding of Ldlr−/− mice induces systemic inflammation, which induces long-lasting trained immunity in myeloid cells (Christ et al., 2018). NLRP3 is a key pathway mediating Western diet-induced trained immunity, and the use of small-molecule inhibitors that block NLRP3 signaling can mitigate its potentially deleterious effects in inflammatory diseases (Christ et al., 2018). ZVAD-fmk (benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone), a pan-caspase inhibitor, inhibits NLRP3 inflammasome activation in atherosclerotic mice, reducing the accumulation of serum IL-1β and plaque cholesterol crystals (Sheedy et al., 2013).
4.1.7 2-DGA d-glucose mimic, 2-deoxy-d-glucose (2-DG), inhibits glycolysis by producing and accumulating intracellularly 2-deoxy-d-glucose-6-phosphate (2-DG6P), which then inhibits the activity of hexokinase and glucose-6-phosphate isomerase and results in cell death (Pajak et al., 2019). BCG immunization causes immunometabolic activation and epigenetic reprogramming, whereas 2-DG’s restriction of glycolysis during BCG-induced training cancels out the enhanced cytokine production (Arts et al., 2016b). In addition, the inhibitory effect of 2-DG on glycolysis also inhibits histone methylation and prevents mevalonate-induced trained immunity (Arts et al., 2016b).
4.1.8 3PO3-(3-Pyridinyl)-1-(4-pyridinyl)-2-propen-1-one (3PO), a small-molecule inhibitor of PFKFB3, inhibits glycolytic flow and is cytostatic to malignant cells (Clem et al., 2008). In cells trained with oxLDL, PFKFB3, a critical rate-limiting enzyme in glycolysis, is increased. The in vitro training protocol’s dose-dependent attenuation of the oxLDL-augmented production of TNF-α and IL-6 upon subsequent stimulation with LPS was achieved by co-incubating 3PO with oxLDL for the first 24 h (Clem et al., 2008).
4.1.9 FluvastatinFluvastatin, the first fully synthetic HMG-CoA reductase (HMGCR) inhibitor, is reported to prevent the growth and spread of certain malignancies (Cai and Zhao, 2021). Fluvastatin prevents trained immunity by downregulating H3K4me3 and blocking the production of proinflammatory cytokines (Arts et al., 2016a). Fluvastatin also prevents the enhanced foam cell production brought on by training with oxLDL and stops the epigenetic reprogramming of BCG, β-glucan, and oxLDL-induced trained immunity (Arts et al., 2016a). Additionally, following OxLDL-induced trained immunity, scavenger receptor CD36 and SR-A mRNA expression increase, whereas cholesterol efflux transporter ATP binding cassette transporter A1 (ABCA1) and ATP binding cassette transporter G1 (ABCG1) decrease. These effects may be reversed by adding fluvastatin (Bekkering et al., 2018).
4.1.10 CeruleninCerulenin is a potent and specific inhibitor of type II FAS found in various bacteria and mammalian tissues (Tomoda et al., 1984). It is an antifungal antibiotic discovered in a culture filtrate of Cephalosporium caerulens (Porrini et al., 2014). Aldosterone levels above normal are linked to a higher risk of CVD in people and the induction of trained immunity in primary human monocytes (van der Heijden et al., 2019). Aldosterone’s trained immunity was reduced when cells were pre-incubated with the fatty acid synthesis inhibitor cerulenin for 1 h before re-stimulating with P3C (van der Heijden et al., 2019).
4.2 Agents that modulate epigenetic reprogramming4.2.1 Ro5-3335Extracellular glucose promotes macrophage-trained immunity and induces a pro-atherogenic phenotype through a glycolysis-dependent pathway. Runx1, which mediates trained immunity produced by hyperglycemia, is implicated by the pattern of open chromatin (Edgar et al., 2021). A benzodiazepine identified from the screen, Ro5-3335, has a direct interaction with Runx1 (Cunningham et al., 2012). In vitro hyperglycemia-induced trained immunity was reversed by pharmacological suppression of Runx1 with Ro5-3335 4.
4.2.2 MTAThe histone methyltransferase inhibitor 5′-deoxy-5′-methylthioadenosine (MTA) is a non-selective methyltransferase inhibitor. OxLDL causes monocytes to develop a proinflammatory phenotype that persists over time and speeds up AS. MTA completely reverses the methylation of histones, which is required for the change in chromatin architecture that results in increased gene transcription, and thus completely reverses the trained immunity phenotype induced by oxLDL (Bekkering et al., 2014).
4.2.3 ResveratrolSirtuin 1 is a nicotinamide adenine dinucleotide (NAD+)-dependent protein deacetylase and master metabolic regulator (Deng et al., 2019). Phytochemical resveratrol, which is abundant in the skin of red grapes and wine, has been studied extensively for its ability to stimulate Sirtuin 1 activity (Lee et al., 2019). Given that histone acetylation is necessary for β-glucan-induced trained immunity, trained immunity in the presence of the histone deacetylase activator resveratrol prevented trained SHIP-1-deficient macrophages from producing more TNF-α (Saz-Leal et al., 2018).
4.2.4 EGCGThe compound epigallocatechin-3-gallate (EGCG) has been discovered to be a new histone acetyltransferase inhibitor (HATi) with broad specificity for the majority of HAT enzymes (Choi et al., 2009). EGCG can also inhibit trained immunity that relies on β-glucan-induced epigenetic reprogramming (Ifrim et al., 2014).
5 Antiatherosclerotic herbal medicine potentially targeting trained immunityThe notion that trained monocytes/macrophages exhibit a broad range of pro-atherogenic phenotypes, including increased production of cytokines/chemokines and foam cells, has recently been extensively supported experimentally (Leentjens et al., 2018). Trained immunity occurs not only in circulating monocytes but also in myeloid progenitors, ensuring a long-term state of hyperactivation of innate immune cells. Trained immunity is mediated by metabolic and epigenetic reprogramming at the level of histone methylation. Theoretically, these processes are amenable to pharmacological intervention. In the past few decades, more studies have shown that various naturally occurring anti-atherogenic natural products, such as flavonoids, phenols, terpenoids, carotenoids, phenylpropanoids, and alkaloids, may be involved in the regulation of pharmacological targets of trained immunity (Supplementary Table S2). We systematically summarize all relevant literature to investigate all potential natural products against trained immunity in ASCVD.
5.1 FlavonoidsFlavonoids are a group of secondary plant metabolites often employed by vegetables for growth and microbial defense (Izzo et al., 2020). Flavonoids can be further classified as flavones, flavonols, flavanones, isoflavonoids, anthocyanins, flavanols, or catechins based on structural distinctions (Kumar and Pandey, 2013). Due to their antioxidant, anti-inflammatory, anti-mutagenic, anti-aging, cardioprotective, antiviral/bacterial, and anticarcinogenic qualities and their ability to influence enzyme performance, they are linked to a variety of positive health impacts. Flavonoids are thought to mediate epigenetic changes, including DNA methylation, histone modifications, and non-coding RNAs (Fatima et al., 2021). We examine some significant natural products that may target monocyte/macrophage and trained immunity in AS in this section.
5.1.1 AlpinetinAlpinetin (7-hydroxy-5-methoxyflavanone), a flavonoid, is the main active component of Alpinia katsumadai Hayata, a traditional medicinal plant. It engages in various biological processes that affect the NF-κB, MAPK, and PI3K signaling pathways, such as antibacterial, anti-ROS, anticancer, and anti-inflammatory actions (Huo et al., 2012; Wu et al., 2020a; Zhang et al., 2020). The inhibition of the NLRP3 inflammasome may be one way of suppressing trained immunity (Christ et al., 2018). Mechanistically, alpinetin inhibits NLRP3-mediated anti-inflammatory activity and reduces mitochondrial ROS production and HIF-1α transcription, thereby inhibiting HIF-1α signaling (Zhang et al., 2020; Zhu et al., 2021). The expression of the toll-like receptor 4 (TLR4) stimulated by LPS may be dramatically downregulated by alpinetin; alpinetin was reported to have had an anti-inflammatory impact by preventing the production of TNF-α, IL-6, and IL-1β in LPS-stimulated human macrophages (Hu et al., 2013).
5.1.2 AnthocyaninsAnthocyanins are water-soluble glycosides of polyhydroxyl and polymethoxyl derivatives of 2-phenylbenzopyrylium or flavylium salts and are partially responsible for the pigmentation of berries (Azzini et al., 2017). The major anthocyanins in plant foods are glycoside forms of anthocyanidins, including pelargonidin, cyanidin, delphinidin, peonidin, petunidin, and malvidin (Khoo et al., 2017). The bioavailability of anthocyanins is higher than previously thought because the parent compounds are immediately absorbed and converted to bioactive metabolites that remain in circulation (Scalbert et al., 2005; Czank et al., 2013). Anthocyanins increase total antioxidant capacity, antioxidant defense enzymes, and high-density lipoprotein (HDL) antioxidant properties in preclinical and clinical populations through multiple measures, thereby reducing CVD risk factors and mortality in patients with coronary heart disease (Garcia and Blesso, 2021). An essential mediator of trained immunity, the NLRP3-caspase-1 inflammasome, is directly activated upstream by ROS, which is an important mediator of trained immunity (Sun et al., 2020). Preclinical research suggests that anthocyanidins regulate cellular cholesterol efflux from macrophages, hepatic paraoxonase 1 expression, and activity to affect reverse cholesterol transport (RCT) and HDL function beyond simple HDL cholesterol content (Millar et al., 2017). In human populations (such as those who are hyperlipidemic, hypertensive, or diabetic), dietary anthocyanin intake is linked to positive changes in serum biomarkers related to HDL function. These changes include an increase in HDL cholesterol concentration and HDL antioxidant and cholesterol efflux capacities (Millar et al., 2017).
The powdered wild blueberry (Vaccinium angustifolium) component high in anthocyanins also reduced lipid buildup in macrophages generated from THP-1 (Del Bo et al., 2016). In accordance with additional studies, the black rice anthocyanin-rich extract blocked the generation of oxLDL and decreased total cholesterol (TC) and LDL-cholesterol (LDL-C) while boosting the amount of HDL-cholesterol (HDL-C) in serum from rats and ApoE−/− mice. In order to lower the risk of an embolism, it also decreased the area of atherosclerotic plaque and improved the stability of the plaque (Xia et al., 2006). In hypercholesterolemic rabbits, fatty streak development and lipid metabolism were slightly influenced by pomegranate peel extract containing anthocyanins (Sharifiyan et al., 2016). This evidence suggests that the potential of anthocyanins to regulate inflammation, lipid buildup, and macrophage may play a role in how an anthocyanin-rich diet lowers the risk of developing CVD.
5.1.2.1 Cyanidin-3-O-β-glucosideCyanidin-3-O-glucoside (C3G) is the anthocyanin with the greatest abundance. C3G is abundantly found in fresh fruits, including grapes, berries, blood oranges, peaches, and apples, and in beverages and colored cereals, such as purple rice and maize (Fang, 2015). One investigation found that methylated proteins, particularly H3K4, lose mono- or dimethyl groups when exposed to C3G or its metabolites, which block the enzyme lysine-specific demethylase 1, which controls histone methylation (Abdulla et al., 2013), thereby directly affecting histone-modifying enzymes (Persico et al., 2021). The results reported here show how dietary C3G intake may effectively control H3K4me3 in the mouse liver, especially in promoter areas (Persico et al., 2021). Recent research in a rat model of high-fat diet (HFD)-induced AS examined the antiatherosclerotic potential of C3G. The findings demonstrated that adding 150 mg/kg of C3G to the diet significantly reduced body weight, visceral adiposity, TG, TC, free fatty acids, and AS index (Um et al., 2013). C3G protected ApoE−/− mice against endothelial dysfunction and AS brought on by hypercholesterolemia by preventing the buildup of cholesterol and 7-oxysterol in the aorta (Wang et al., 2012).
5.1.3 BaicalinBaicalin is a flavonoid active ingredient extracted from the roots of Scutellaria baicalensis Georgi, a plant used for many years in Chinese traditional medicine to treat various inflammatory illnesses (Liu and Liu, 2017; Riham et al., 2019). One study showed that baicalin could control metabolic diseases in vivo. The therapy with baicalin in HFD rats markedly improved fasting blood glucose levels (Guo et al., 2009). Furthermore, baicalin is reported to inactivate succinate dehydrogenase (SDH) to inhibit ROS production and protect glutamine synthetase (GS) protein stability from oxidative stress to improve glutamate handling and reduce excitotoxicity (Song et al., 2020a).
5.1.4 ChrysinChrysin (5,7-dihydroxyflavone) is a flavonoid that naturally occurs in food and is frequently found in honey and propolis, among other plant extracts (Song et al., 2020b). Chrysin possesses various biological qualities, including anti-inflammatory, anti-bacterial, antidiabetic, anticancer, antioxidant, and anti-allergenic actions (Kasala et al., 2015; Mani and Natesan, 2018). One study showed that chrysin has a good expansion effect on human HSCs due to its antioxidant properties by delaying HSC differentiation, inhibiting ROS-activated apoptosis, and regulating cyclin-dependent kinase inhibitors, which can maintain the self-renewal and multilineage differentiation potential of human HSCs (Litviňuková et al., 2020). Chrysin is an emerging histone deacetylase inhibitor for epigenetic regulation in cancer studies (Ganai et al., 2021).
Chrysin may reduce inflammation by modulating M1/M2 status. It promotes the anti-inflammatory M2 phenotype and suppresses the M1 phenotype in peritoneal and cultured macrophages in vitro by activating PPAR-γ (Feng et al., 2014). One study showed that chrysin inhibited NLRP3 inflammasome activation and increased IL-1β levels to reduce synovitis (Liao et al., 2020). Another study showed that chrysin inhibits ROS-mediated Akt/mTOR signaling in cells and induces autophagy (He et al., 2021). The study showed that the overexpression of PPARγ, liver X receptor (LXR)α, ABCA1, and ABCG1 expression led to a considerable increase in HDL-mediated RAW264.7 macrophage cholesterol efflux following chrysin treatment (Lin et al., 2015b).
5.1.5 DaidzeinDaidzein, a substance mostly present in soy foods and plants such as red clover, is one of the most studied and potent phytoestrogens (Yang et al., 2012). Studies in non-human primates have shown that dietary intake of soy protein can interfere with related epigenetic changes that may influence the etiology of complex diseases (Howard et al., 2011). By stimulating the PPARγ-LXRα-ABCA1 pathway, daidzein protected low-density lipoprotein (LDL) from oxidation and increased paraoxonase-1 (PON-1) activity in Huh7 cells, which may control cholesterol efflux (Schrader et al., 2012; Ikhlef et al., 2016). Furthermore, daidzein therapy decreased blood cholesterol and increased TG levels in middle-aged male rats given HFD designed to induce AS (Sosić-Jurjević et al., 2007). These suggest that daidzein has antiatherosclerotic potential.
5.1.6 Ellagic acidEllagic acid (EA) is a dilactone of hexahydroxydiphenic acid that may be found in various nuts, fruits, and vegetables, such as pomegranates, walnuts, black raspberries, raspberries, almonds, and strawberries (Galano et al., 2014). According to in vitro, in vivo, and clinical investigations, it has a wide range of physiological actions, including anti-inflammatory, antioxidant, antibacterial, anticarcinogenic, antiplasmodial, antiviral, hepatoprotective, antifibrotic, immunomodulatory, and neuroprotective activities (Gupta et al., 2021). A study showed that EA promotes hematopoietic progenitor cell proliferation and megakaryocyte differentiation (Gao et al., 2014). Other studies have shown that EA interrupts the sequential histone remodeling steps of adipocyte differentiation by reducing the coactivator-associated arginine methyltransferase 1 (CARM1) activity, including histone acetylation and dissociation of HDAC9 from chromatin (Kang et al., 2014). Pomegranate peel polyphenols, in particular pomegranate ellagic acid (PEA), also boosted ApoA1-mediated macrophage cholesterol efflux by upregulating ABCA1 and LXRα and inhibited macrophage lipid buildup by lowering the expression of CD36 (Zhao et al., 2016).
5.1.7 EGCGEGCG, a typical polyphenol flavonoid molecule with eight free hydroxyl groups, is the most common (Chakrawarti et al., 2016). Research has revealed that EGCG has antibacterial, antiviral, antioxidant, anti-arteriosclerosis, anti-thrombosis, anti-vascular proliferation, anti-inflammatory, and anti-tumor activities (Liu and Yan, 2019). As a histone acetyltransferase inhibitor, EGCG can significantly inhibit the training of monocytes (Ifrim et al., 2014). In different research, EGCG-loaded nanoparticles targeted macrophages via their CD36 receptor, reduced the release of inflammatory factors by mouse peritoneal macrophages, and reduced the lesion surface area of arterial plaques in LDLr−/− mice (Zhang et al., 2019). EGCG also inhibited the oxLDL-induced overexpression of SR-A in the same cell line, reducing oxLDL absorption and the formation of foam cells (Chen et al., 2017). EGCG regulated macrophage polarization toward the M2 state. EGCG decreased the expression of proinflammatory M1 mediators, iNOS, TNF-α, IL-1β, and IL-6, in the LPS-administered lung microenvironment and increased the expression of KLF4, Arg1, and ym1, which enhanced the M2 phenotype of macrophages (Almatroodi et al., 2020).
5.1.8 HesperidinHesperidin (3′,5,7,-trihydroxy-4′-methoxyflavanone), a flavanone family of flavonoids, is a derivative of hesperetin, which is present in citrus fruits, such as oranges and grapefruit (Muhammad et al., 2019). Hesperidin has several pharmacological effects, with the main ones being the stimulation of antioxidation, the inhibition of the generation of proinflammatory cytokines, and the inhibition of the proliferation of cancer cells (Li and Schluesener, 2017). A study using metabolic tracing studies showed that TLR signaling in mouse and human macrophages redirects metabolic flux to increase acetyl-CoA for glucose production, thereby enhancing histone acetylation (Christ and Latz, 2019). According to a preclinical study, hesperetin provided neuroprotection by controlling the TLR4/NF-κB signaling pathway in response to the harmful effects of LPS (Muhammad et al., 2019). Hesperetin decreased the generation of foam cells produced from THP-1 by promoting ABCA1 expression by boosting the activities of the ABCA1 promoter and LXR enhancer, upregulating the ApoA1-mediated cholesterol efflux (Iio et al., 2012).
5.1.9 IcariinIcariin, one of the primary ingredients in epimedium, is an 8-isopentane flavonoside that has several pharmacological benefits, including enhancement of cardiovascular function, promotion of hematological function, prevention of neuronal damage, and anti-osteoporosis properties (El-Shitany and Eid, 2019). Icariin decreased RAW264.7 macrophage infiltration at atherosclerotic lesions by reducing the CX3CR1–CX3CL1 interaction, which is directly related to monocyte adhesion and migration (Wang et al., 2016a).
5.1.10 PratenseinPratensein is a compound extracted from Radix Polygala roots. It has anti-inflammatory, anti-apoptotic, and antioxidant effects (Liu et al., 2016b). Pratensein increases the expression of the ABCA1 protein and HDL levels in HepG2 cells (Gao et al., 2008). AS is brought on by passive LDL transport across damaged endothelial cells. Recent research has revealed a novel therapeutic target in the fight against AS: scavenger receptor class B type I (SR-BI)-mediated endothelial LDL transcytosis. This process increases LDL entry into the arterial wall and the development of AS (Huang et al., 2019). Further investigat
留言 (0)