
記住我
Extraintestinal pathogenic Escherichia coli (ExPEC) is responsible for 80%–90% of community acquired urinary tract infections (Ejrnaes, 2011) as well as 30% of bacteremia (Diekema et al., 2002), thus is one of the major agents of human diseases. In addition, ExPEC infects a broad spectrum of birds and mammals, such as causing sepsis and sudden death in swine and avian species (Dho-Moulin and Fairbrother, 1999; Ding et al., 2012), thereby is an important zoonotic pathogen to threaten public health. To cause sepsis, most virulent ExPEC strains have the ability to survive and proliferate within host bloodstream (Kaper, 2005). Although numerous virulence factors have been identified in diverse ExPEC pathotypes, the underlying mechanisms of ExPEC breaking through the barriers and evading immune clearance during bloodstream infection are incompletely understood.
Numerous serine protease autotransporters of enterobacteria (SPATEs) from class-2 have been identified in pathogenic E. coli to degrade a variety of mucins, thus play critical roles during the bacterial infection (Harrington et al., 2009; Ruiz-Perez and Nataro, 2014; Gibold et al., 2016), while related mechanisms involving in bloodstream infection have never been clarified in a sepsis isolate. Mucin is the major component of mucus layer widely presenting on the surface of epithelial tissues in the respiratory, urinary and genital tracts (Harrington et al., 2009; Navarro-Garcia et al., 2010; Gibold et al., 2016). In adherent-invasive E. coli (AIEC) strain LF82, the class-2 SPATE “Vat” promotes crossing of the intestinal mucus layer to cause Crohn’s disease (Gibold et al., 2016). Besides contributing to intestinal colonization, SPATEs “Pic” and “Tsh” are expressed during urinary tract infection of uropathogenic E. coli (UPEC), and significantly associated with the acute pyelonephritis (Heimer et al., 2004). Class-2 SPATEs also display a lectin-like activity with affinity to degrade diverse O-glycosylated mucin-like substrates, including the leukocyte surface O-glycoproteins (Henderson et al., 1999; Leyton et al., 2003; Ruiz-Perez et al., 2011). The cleavages of this type O-glycoproteins usually damage their vital roles in numerous cellular functions in leukocytes, thus result in immunomodulation (Szabady et al., 2009; Ruiz-Perez et al., 2011). These observations suggested that the SPATEs may mediate the immune evasion of bacterial pathogens from leukocytes, while there were no further studies providing solid evidences to verify that these functions contribute to the bacterial survival during bloodstream infection.
In this study, two class-2 SPATE encoding genes (vatPU-1 and tsh PU-1) of porcine ExPEC strain PU-1 were identified to be significantly upregulated in host blood but not in serum in vitro. Following works identified that these two SPATEs significantly interact with hemoglobin of red blood cell (RBC) for adhesion and spreading within bloodstream, and impair polymorphonuclear leukocytes’ (PMNs) functions via cleaving the mucin-like O-glycoproteins for immune evasion, which help to better understand how ExPEC colonizing within host bloodstream and cause severe sepsis.
Materials and methodsBacterial strains and genetic manipulationsBacterial strains and plasmids used in this study were listed in Table S1. PU-1 is an O2:K1 ExPEC strain (isolated from the blood of a piglet) causing acute sepsis in mouse infection model (Ma et al., 2020; Ma et al., 2021). All strains were grown on Luria-Bertani (LB) broth medium at 37°C with 180 rpm, supplemented with corresponding antibiotics, or isopropyl–D-thiogalactopyranoside (IPTG) when necessary. DNA amplification, ligation and electroporation were performed as previously described (Ma et al., 2018) unless otherwise indicated. Deletion mutants were constructed using the λ red mutagenesis method (Datsenko and Wanner, 2000), and the details of primers, restriction enzymes and fragments’ deletion have been listed in Table S2. All restriction and DNA-modifying enzymes were purchased from Thermo Fisher Scientific (Waltham, MA, USA) and performed according to the supplier instruction.
Ethical approval statementPorcine blood and mucus of the respiratory tract were collected from the healthy pigs of a slaughterhouse to perform the following studies. Blood from healthy donators was obtained from Jiangsu province Blood Center, and related experiments were approved by the Medicine Human Subjects committee of Jiangsu Province. Five-week-old female specific pathogen free (SPF) BALB/c mice were purchased from Yangzhou University (Comparative Medicine Center). All animal experiments were performed in strict accordance with the animal welfare standards of the Animal Research Committee Guidelines of Jiangsu Province (License Number: SYXK (SU) 2017-0007), and approved by the Ethics Committee for Animal Experimentation of Nanjing Agricultural University.
Mouse infection assayTen mice in each group were challenged by intraperitoneal injection with the indicated strain at the designed doses and monitored for symptoms until 7 days post-infection. The negative-control group was challenged with an equal volume of sterile PBS. To evaluate bacterial proliferation in vivo, the bacterial load assay was conducted. Five mice in each group were inoculated with 1 × 106 CFU/mouse of the indicated strain, the infected blood was harvested at the designed time of post-infection, and then serially diluted in PBS and plated on LB agar to enumerate the CFU.
RNA isolation and RT-qPCR analysisTotal RNA was extracted with the E.Z.N.A. bacteria RNA isolation kit (Omega), and residual genomic DNA was then removed by digestion with DNase I (TaKaRa). The PrimeScript RT reagent kit (TaKaRa) was used for cDNA synthesis. The RT-qPCR was performed using SYBR premix Ex Taq (TaKaRa) with the gene-specific primers. The relative amount of target gene mRNA was normalized to the transcript of housekeeping gene tus (Ma et al., 2018), and the relative fold change was calculated by the threshold cycle (2-ΔΔCT) method. The reported values represented the mean ± SD of three independent RNA extractions.
ELISA with hemoglobinThe recombinant His6-VatPU-1 and His6-TshPU-1 proteins were purified by Ni-NTA Spin Columns (QIAGEN) from BL21 (DE3) carrying the recombinant pET-21a plasmid after IPTG induction. Human hemoglobin (Sigma-Aldrich) prepared at a concentration of 50 μg/mL in PBS were coated onto separate wells of a 96-well plate. The purified His6-VatPU-1 and His6-TshPU-1 were added to each well for 1 h at 37°C, and then washed three times and incubated with the anti-His antibody (Abcam, 1:2000) at 37°C for 2 h. After thrice wash, the processed membranes were stained with the HRP conjugated secondary antibodies (Thermo Fisher, 1:2000) at 37°C for 1 h, and detected using the 3,3’-diaminobenzidine. The reaction was stopped after 30 min by addition of 1 M H2SO4, and absorbance was measured at 450 nm. Three independent experiments were performed, with four wells for every reaction in each experiment, and the values obtained were averaged.
Mucin gel penetration assay and cleavage assayThe mucin gel penetration assay were used as previously described (Henderson et al., 1999; Silva et al., 2003). A solution containing 10% mucin of porcine respiratory tract and 0.3% agarose in HBSS was loaded into a 1-mL injection syringe, creating a mucous column. A 0.1 mL prepared bacterial cells (10 × 109 cells/mL) were layered onto the mucin. The columns were incubated for 3 h at 37°C in a vertical position. Afterwards, five fractions (each one contains 0.2 mL) were collected from the button by applying gentle pressure. Each fraction was serially diluted and plated to the LB agar media for CFU enumerating. Western blot assays were performed to analyses the cleavage activity of SPATEs to mucin. The mucins were extracted from the mucus of porcine respiratory tract as previously described (Gibold et al., 2016). Degradation reactions were separated by 12% SDS-PAGE, and then transferred to PVDF membranes (Bio-Rad) for subsequent blocking, washing, incubating with the specific and secondary antibodies (using as the instructions of manufacturer), and detecting using the 3,3’-diaminobenzidine.
Flow cytometryBefore incubating with the conjugated mAb of APC-CD43 (Invitrogen), PMNs were incubated with the human IgG to block Fc receptors. The prepared cells were incubated with SPATEs for the indicated times, and then stained by incubation with the dye-conjugated antibodies specific to the extracellular domain of host glycoproteins. The positive staining with antiCD16 and CD16b mAbs (R&D Systems) were selected as the low forward-scatter and high side-scatter characteristics, respectively, and then used to gate the neutrophils. The samples were analyzed in an Accuri C6 fow cytometer (BD Accuri)/fluorescence-activated cell sorter (FACS), and analyzed using the CFlow plus software (BD Accuri).
PMNs chemotaxis and transendothelial migration assaysChemotaxis and transmigration assays were performed according to previously described protocols (Yamamoto et al., 2000; Ruiz-Perez et al., 2011). For the chemotaxis assay, 3 × 105 calcein-stained human PMNs were incubated with the indicated protein in the upper chamber of Transwell, and 100 mM of IL-8 (MedChemExpress) or 100 nM of fMLP (N-formyl-methionineleucine-phenylalanine, MedChemExpress) was added to the lower chamber as chemoattractant. After 4 h incubation, the PMNs that had migrated toward the chemoattractant were collected for counting with a Fluoroskan fluorometer. For transendothelial migration assays, human brain microvascular endothelial cells (HBMECs) were seeded on the inserts of Transwells at a concentration of 2 × 104 cells/well, and cultured until the monolayers were confluent (~ 4 days). Afterwards, similar operating steps were performed as the chemotaxis assays.
Detection of blood indicatorsThe blood samples were analyzed by a Blood RT (Routine Test) machine in the Animal Hospital of Nanjing Agriculture University. The whole bloods of mice infected with the indicated bacterial strains were collected, and centrifuged to get the sera. The mouse interleukin 6 and 8 ELISA test kits (HUYU biological technology Co Ltd, shanghai) were used as the instructions of manufacturer to detect the levels of interleukin 6 and 8 release in the sera.
Statistical analysisStatistical analyses were performed using Prism 8.0 (GraphPad), and the full details were described below. Two-way ANOVA was used for the qRT-PCR assay, One-way ANOVA was performed for the results of bacterial survival assay, blood routine test, IL-6 and IL-8 release, PMNs chemotaxis and transmigration. For infection experiments, survival data were analyzed with the log rank test. For all tests, a P value < 0.05 were considered statistically significant, and all data were shown as mean ± SD.
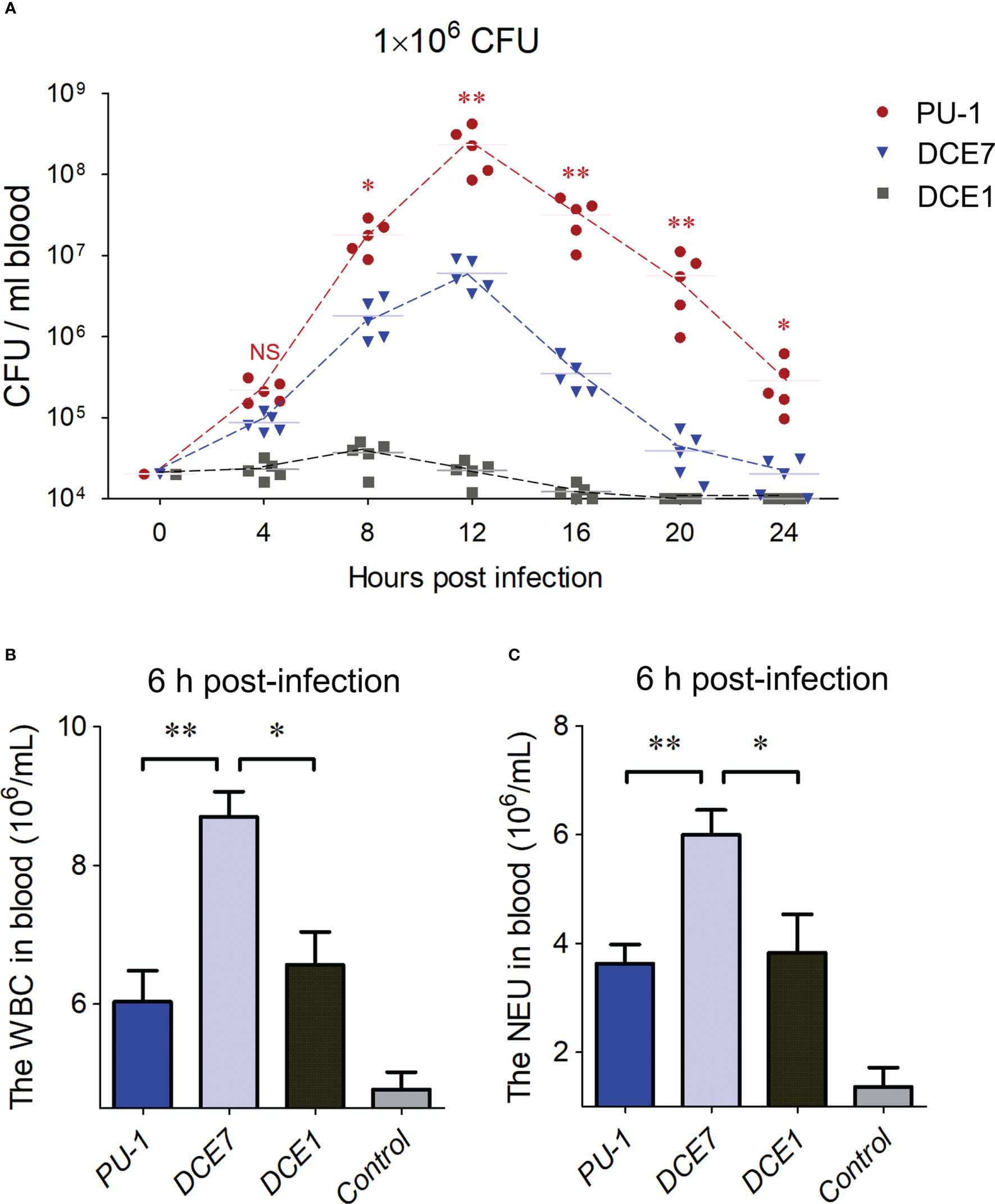
ResultsLow levels of leukocytic activation were induced by ExPEC strain PU-1 during the severe bloodstream infectionPU-1 is an O2:K1 ExPEC strain causing acute sepsis in mouse infection model (Ma et al., 2020; Ma et al., 2021). The curve of blood bacterial loads in mice infected by strain PU-1 showed the maximum value (more than 108 CFU/mL) at 12 h post-infection (Figure 1A), which was remarkably higher than the infection groups of virulent ExPEC strain DCE7 from phylogenetic group D (Zhu et al., 2017), and strain DCE1 from group A. Furthermore, the blood of mice (with obvious clinical symptoms: tremble, orbital hemorrhage, anorexia, ataxia, anaesthesia et al.) in PU-1 infection group showed a significantly slighter decrease in bacterial loads after 12 h post-infection than the mice challenged with strains DCE7 and DCE1 (Figure 1A). The blood routine testing results showed that white blood cell (WBC) and neutrophil cell (NEU) counts of peripheral blood from the PU-1 infection group were significantly less than that of DCE7 infection group (Figures 1B, C) at the 6 h post-infection (no significant differences in blood bacterial loads between PU-1 and DCE7 infection groups at this point in time). These data suggested that strain PU-1 has the potential to modulate the host immune responses for its optimal blood infection in vivo.
Figure 1 ExPEC strain PU-1 caused a heavy bacterial load in infected mouse blood. (A) The curves showed the bacterial loads in bloods from the mice challenged with different E. coli strains until 24 h post infection. The ExPEC strains DCE7 and DCE1 were used as controls here, respectively. Statistical significance of PU-1 infection group was determined by a one-way ANOVA test based on comparisons with the DCE7 infection group (**P < 0.01, *P < 0.05). (B, C) Blood routine testing detected the white blood cell (WBC) and neutrophil cell (NEU) counts of peripheral blood from the mice infected with indicated ExPEC strains at the 6 h post-infection. Statistical significance was determined by a one-way ANOVA test based on comparisons with the wild-type group (**P < 0.01, *P < 0.05). NS, no significance.
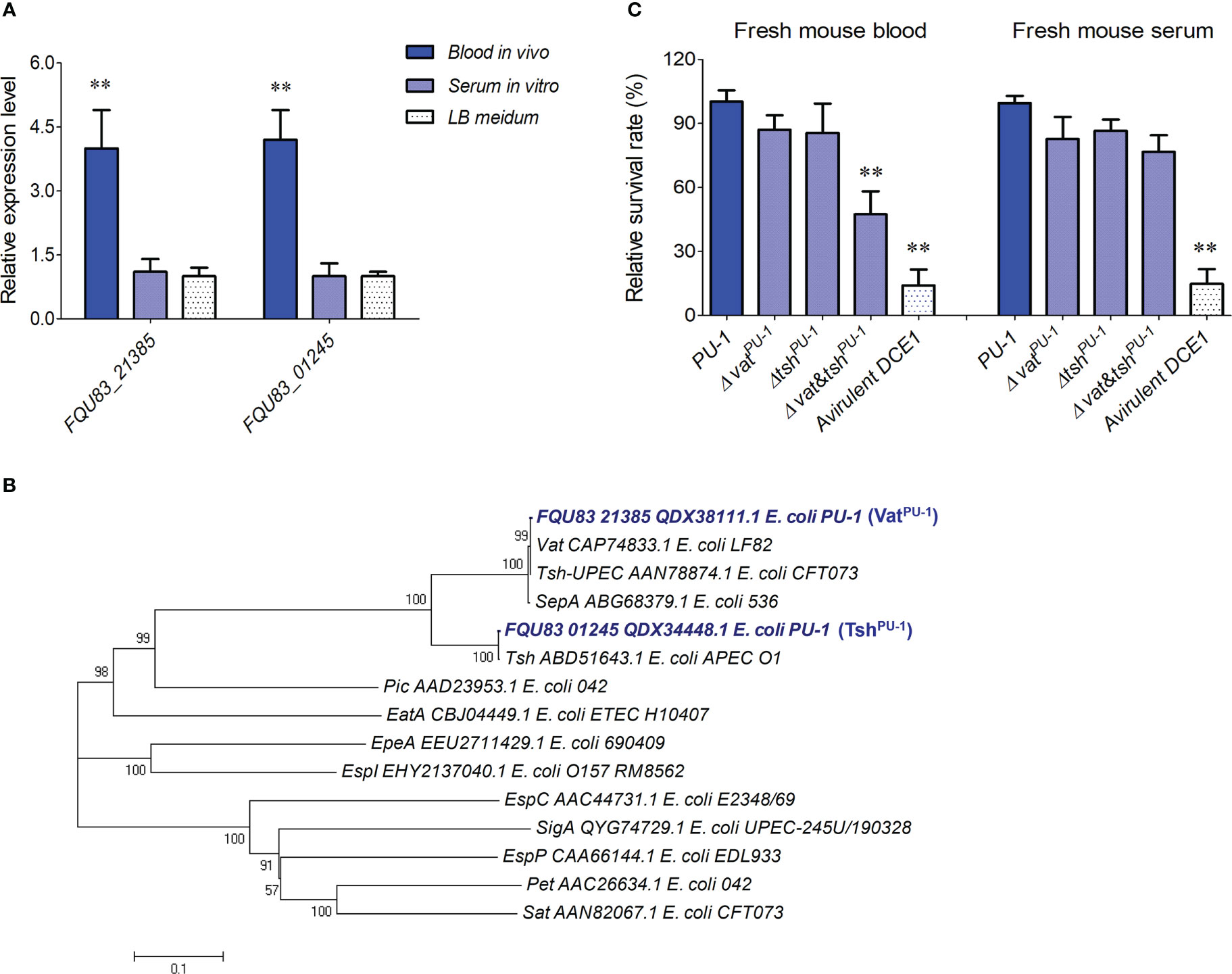
Two serine protease autotransporters, VatPU-1 and TshPU-1, were identified to contribute to blood survival of strain PU-1 in vitroComparative analysis based on our previous transcriptome data (Ma et al., 2020) found two special genes, FQU83_21385 (chromosome encoding) and FQU83_01245 (plasmid encoding), which encoded two serine protease autotransporters of enterobacteria (SPATEs), and showed the significant upregulation of more than 3 folds in animal blood but not in serum (Figure 2A). Phylogenetic analysis showed that the FQU83_21385 and FQU83_01245 belong to the class-2 of SPATEs (Figure 2B), and share highest sequence identities with the well-studied virulence factors Vat homologue of strain LF82 and Tsh homologue of strain APEC O1, respectively, thereby were redesignated as VatPU-1 and TshPU-1 in strain PU-1. To explore whether these two SPATEs involving in ExPEC’s bloodstream survival, non-polar deletion mutant strains were constructed under the background of wild-type strain PU-1. The results of bacterial counting demonstrated that the inactivation of VatPU-1 alone or TshPU-1 alone did not affect the bacterial survival in host blood (Figure 2C). It should be noted that mutant strain with double deletions of vatPU-1 and tshPU-1 significantly decreased survival in host blood, while had no significant effect for the bacterial survival in host serum (Figure 2C). These results suggested that at least one SPATE in strain PU-1 was required to resist the clearance mediated by immunocytes within blood, while was not involved in resistance to the bactericidal effects of serum.
Figure 2 Identification of two serine protease autotransporters as the potential facilitators for optimal blood infection in strain PU-1. (A) The transcriptional changes of indicated genes in ExPEC strain PU-1 response to host blood and serum. The data were normalized to the housekeeping gene tus transcript. Mean values and SDs of triplicate samples are indicated. Statistical significance was determined by a two-way ANOVA test based on comparisons with the bacterial cells cultured in LB medium (**P < 0.01). (B) Phylogenetic analysis of SPATEs from E coli. A neighbor-joining tree (bootstrap n = 1000; Poisson correction) was constructed based on a ClustalW alignment of the amino acid sequences of SPATEs using the MEGA software version 5.0. (C) Incubations of indicated bacterial strains within fresh blood and serum. The survival rates were calculated by measuring the bacterial counts. The porcine ExPEC strain DCE1 from phylogenetic group A were used as a control here. Statistical significance was determined by a one-way ANOVA test based on comparisons with the wild-type group (**P < 0.01).
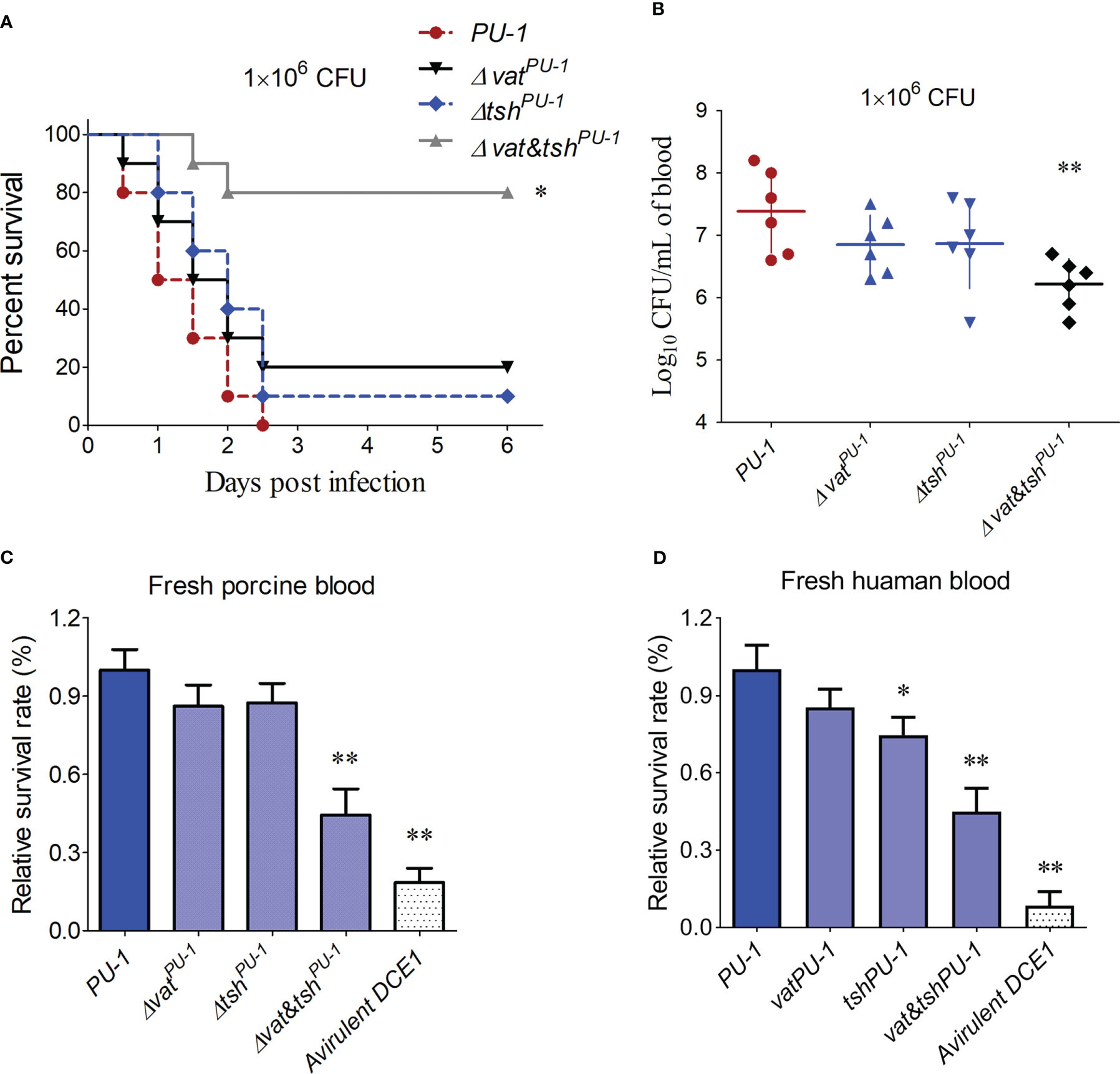
VatPU-1 and TshPU-1 contributed to bloodstream infectionAn animal infection test using the BALB/c mice was employed to further certify the pathogenic roles of these two SPATEs during bloodstream infection. As shown in Figure 3A, the mice infected by the Δvat&tshPU-1, but not the single deletion mutants ΔvatPU-1 and ΔtshPU-1, showed a significantly higher survival rate (80%), compared with the 100% death of mice infected by the wild-type strain. Consistently, only the double deletions of vatPU-1 and tshPU-1 significantly attenuated the bacterial loads in mice blood compared with the wild-type strain (Figure 3B). Porcine and human bloods were then used to assess the survival rates of indicated ExPEC strains. The results demonstrated that the mutant strain with double deletions of vatPU-1 and tshPU-1 significantly decreased survival in both of these two types of blood (Figures 3C, D), while the inactivation of VatPU-1 alone or TshPU-1 alone did not affect the bacterial survival here. These data indicated that VatPU-1 and TshPU-1 are critical for the full virulence and blood infection in ExPEC strain PU-1.
Figure 3 Mouse infection assay identified the pathogenic roles of VatPU-1 and TshPU-1. (A) Effect of vatPU-1 or tshPU-1 deletion on strain PU-1 pathogenicity. Survival curve of mice infected with 1 × 106 CFU/mouse bacteria (ten mice per group). (B) Systemic infection experiment was conducted to assess bacterial load in mouse blood. Bacterial reisolation from the blood at 16 h post-inoculation was quantified by plate count. (C, D) Porcine and human bloods were then used to assess the bacterial survival of indicated ExPEC strains. The survival rates were calculated by measuring the bacterial counts. Statistical significance was determined by a one-way ANOVA test (**P < 0.01, *P < 0.05).
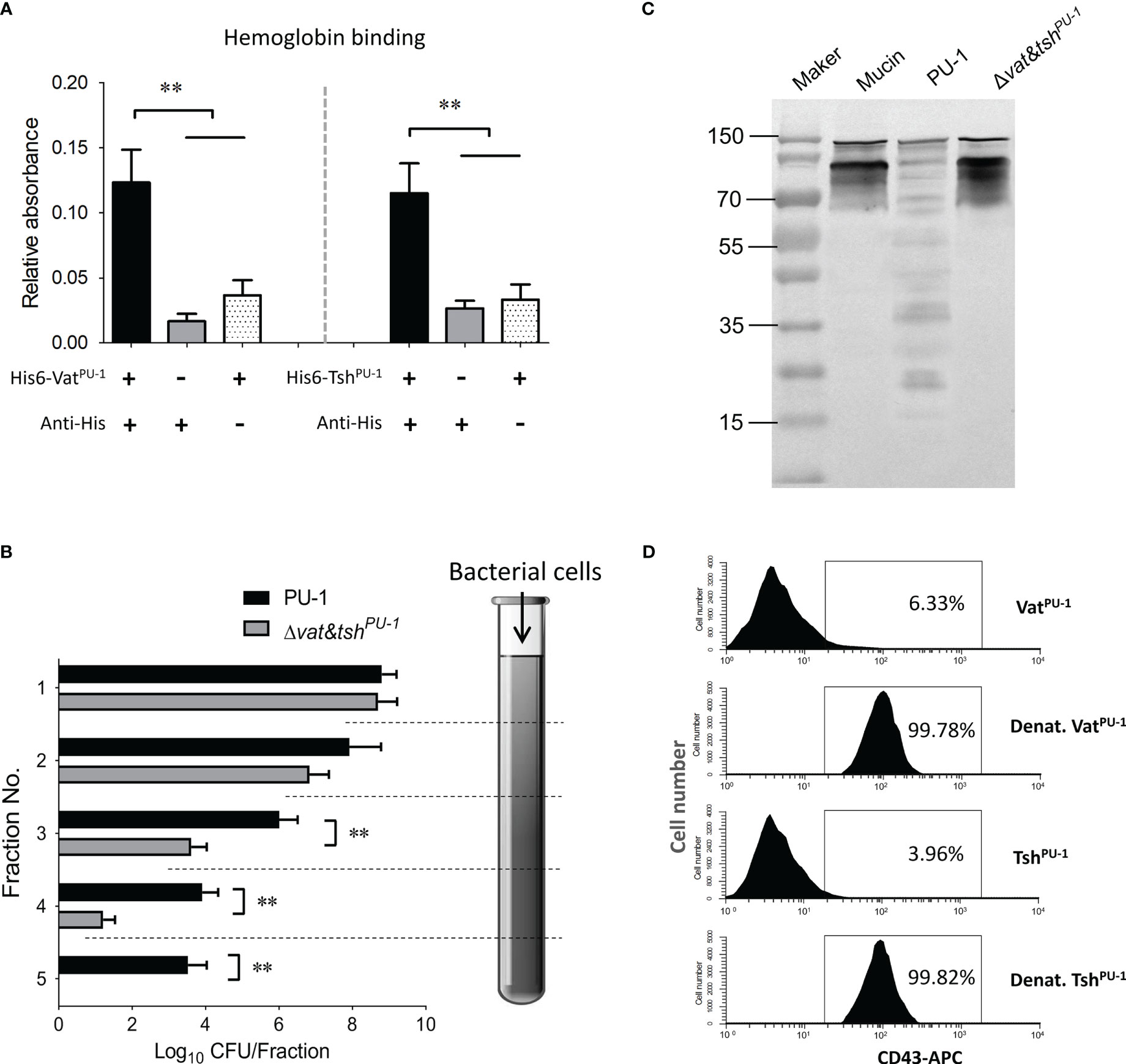
VatPU-1 and TshPU-1 interacted and degraded mucin-like O-glycoproteinsNumerous class-2 SPATEs of pathogenic E. coli have been reported to bind to and cleave mucin-like proteins for optimal colonization (Henderson et al., 1999; Dutta et al., 2002; Leyton et al., 2003; Harrington et al., 2009), including Pic, Tsh, Vat, et al. In this study, both of VatPU-1 and TshPU-1 were identified to have the abilities to bind to hemoglobin (Figure 4A), a well-known mucin-like glycoprotein widely presenting in red blood cells (RBCs). We then investigated the ability of VatPU-1 and TshPU-1 to degrade mucus by using a column penetration assay. Unlike Δvat&tshPU-1, wild-type strain PU-1 penetrated through the entire mucus column (fractions 1 to 5, Figure 4B). In the fractions 3 and 4 (middle of the column), the efficiency of Δvat&tshPU-1 was about two orders of magnitude lower than that of PU-1 strain at penetrating the mucus column. In order to investigate the mucinolytic activity of VatPU-1 and TshPU-1, we examined the ability of related strains to hydrolyze porcine respiratory mucins by Western blotting. The cultural supernatant of wild-type strain PU-1 was able to degrade respiratory mucins unlike those of Δvat&tshPU-1 (Figure 4C). Overall, these results suggest that VatPU-1 and TshPU-1 promote the bacterial penetration through the mucus layer by altering the gel-forming mucins.
Figure 4 VatPU-1 and TshPU-1 bound to and cleaved diverse O-glycosylated mucin-like proteins. (A) SPATEs bind to the hemoglobin. Human hemoglobin was coated onto a 96-well microplate and incubated with the purified proteins. Binding was detected by an indirect ELISA. (B) The assessment of mucin degradation by using a column penetration assay. Quantification of indicated ExPEC strains in fractions eluted from columns filled with gel-forming mucus (1 to 5 fractions: top to bottom of the column). (C) Mucin extracted from porcine respiratory tract was incubated overnight with filtered culture supernatants of indicated ExPEC strains. Mucin degradation was observed by Western blot using an anti-MUC2 antibody. (D) The degradation mediated by VatPU-1 and TshPU-1 to the extracellular domain of O-glycosylated mucin-like protein CD43 on human leukocytes. The PMNs were isolated from human blood and incubated with the purified VatPU-1, TshPU-1, or denatured proteins at 37°C for 30 min. Flow cytometry was employed to analyze these samples using monoclonal antibodies against the extracellular domains of CD43. Flow cytometry data are representative of at least three independent experiments. **P value < 0.05, which is considered statistically significant.
SPATEs have been confirmed to target a broad range of mucin-like glycoproteins present not only on the RBCs and epithelial cells, but also on the surface of leukocytes. To determine whether VatPU-1 and TshPU-1 cleaved such proteins, the CD43, a major cell surface glycoprotein expressed on leukocytes, was addressed. We performed flow cytometry analyses of human PMNs treated with purified VatPU-1, TshPU-1 and denatured proteins to further confirm the CD43 cleavage on cell surface. Staining with anti-CD43 mAbs revealed that the extracellular CD43 was present on the surfaces of PMNs after treatment with denatured VatPU-1 and denatured TshPU-1 (Figure 4D), while was cleaved, becoming undetectable on cells treated with purified VatPU-1 and TshPU-1. These results, coupled with the previous study verifying the cleavages of CD44, CD45, CD93 and CD162 by the Tsh homologue from Avian Pathogenic E. coli (APEC) (Ruiz-Perez et al., 2011), suggested that VatPU-1 and TshPU-1 have the potential to cleave diverse O-glycosylated mucin-like proteins located on peripheral blood leukocytes, which may facilitate the bacterial survival within host bloodstream.
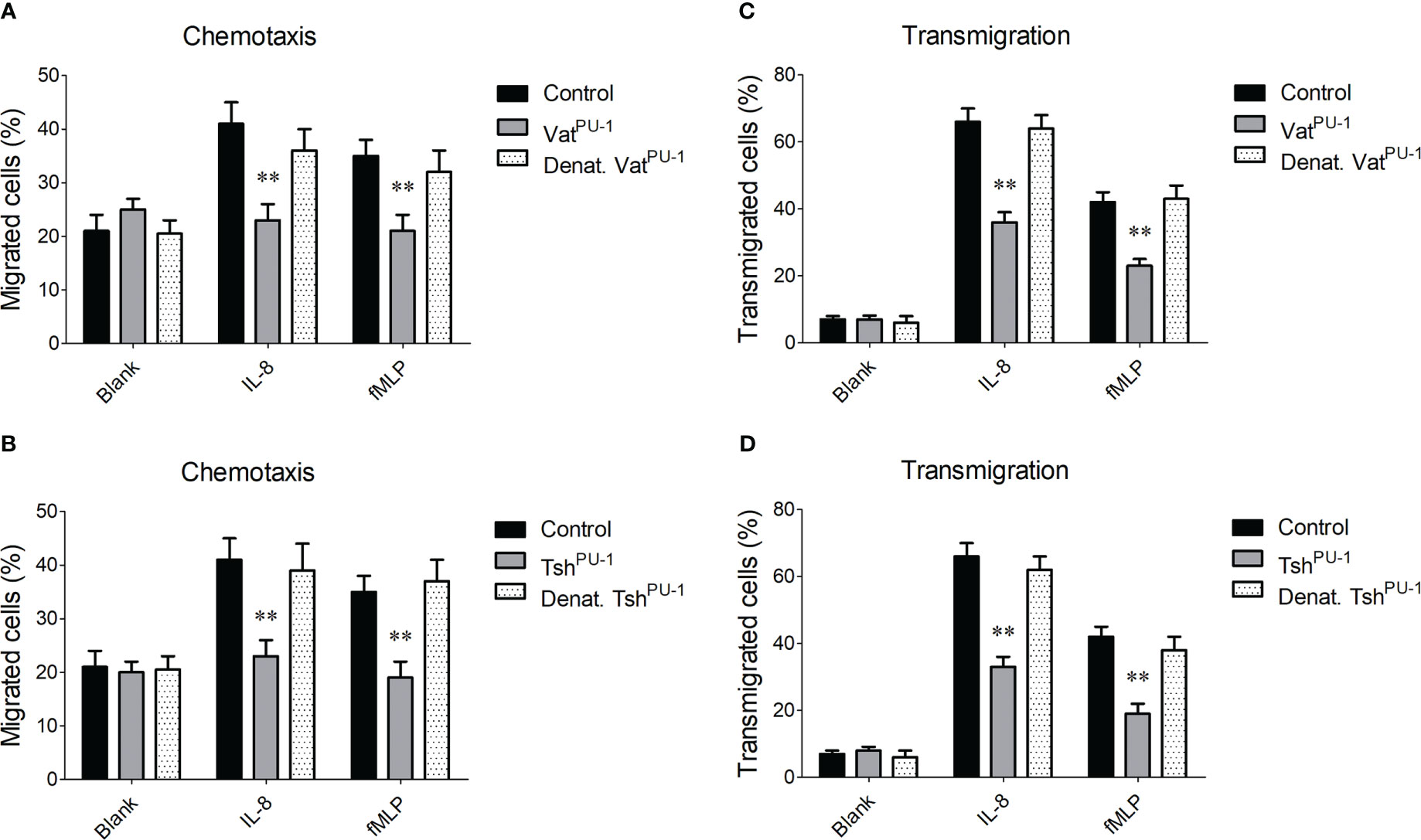
VatPU-1 and TshPU-1 impaired the PMNs’ chemotaxis and transmigrationWe next explored whether the cleavage of mucin-like glycoproteins mediated by VatPU-1 and TshPU-1 disturb the function of peripheral blood leukocytes during ExPEC infection. Here chemotaxis and transmigration assays of PMNs were performed according to previously described protocols (Ruiz-Perez et al., 2011; Ma et al., 2018). For the chemotaxis assays, IL-8 or fMLP within the lower chamber of Transwell were used to stimulate the translocation of PMNs through an abiological membrane from the upper chamber, and PMNs that migrated in the lower chamber were measured. The movements of PMNs stimulated by IL-8 and fMLP were significantly enhanced compared with the untreated PMNs in this model (Figure 5A), while incubation with the VatPU-1 but not the denatured VatPU-1 was significantly reduced the activated movement to the similar levels with the untreated PMNs. Similar results were also observed when the PMNs were incubated with the TshPU-1 (Figure 5B). These observations suggested that both two SPATEs can impair the PMNs’ chemotaxis. For the transmigration assays, the PMNs within the upper chamber of Transwell were stimulated to transmigrate through the endothelial cell monolayers, and PMNs in the lower chamber were measured after 4 h incubation. Expectedly, the transmigration of PMNs incubated with VatPU-1 or TshPU-1 was significantly reduced compared with the buffer alone as negative control (Figures 5C, D); denaturation of VatPU-1 or TshPU-1 by heating completely abolished the above effects. Taken together, these observations suggest that the PMNs’ functions were significantly impaired by the interaction with VatPU-1 and TshPU-1 during the bloodstream infection of ExPEC.
Figure 5 The impairment mediated by VatPU-1 and TshPU-1 to the PMNs’ chemotaxis and transmigration through endothelial cell monolayers. (A, B) Assessment of PMNs’ chemotaxis. The calcein- labeled PMNs were treated with indicated proteins, or PBS vehicle control in the upper chamber of Transwell; IL-8 or fMLP were added to the lower chamber. Penetration of cells through the membrane was measured after 4 h fluorometrically. (C, D) Assessment of PMNs’ transmigration. The preincubated PMNs were applied to the upper chamber of Transwell supporting HBMEC, and transmigrated PMNs were enumerated as mentioned before. Statistical significance was determined by a one-way ANOVA test based on comparisons with the control group (** P<0.01). Error bars represent the SDs for three independent experiments.
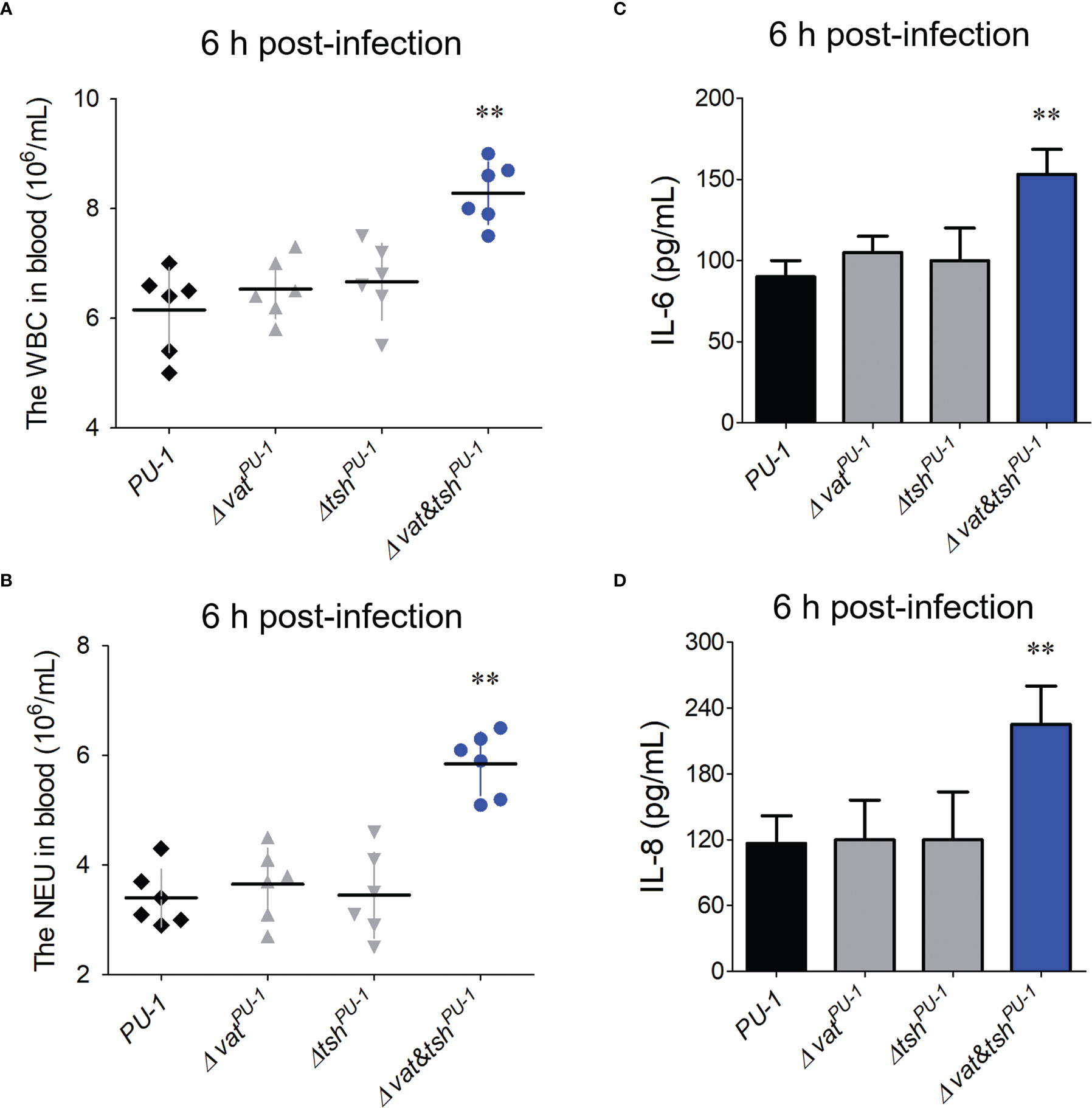
VatPU-1 and TshPU-1 modulated the host immune responses in vivoFurther study managed to investigate the potential modulations of SPATEs to the immune responses in a mouse infection model. Blood routine testing results showed that white blood cell (WBC) and neutrophil cell (NEU) counts of peripheral blood from Δvat&tshPU-1 infection group were significantly higher than that of wild-type infection group (Figures 6A, B). The cytokines’ detection was then performed. Compared with wild-type strain, the inactivation of VatPU-1 alone or TshPU-1 alone did not affect the release levels of IL-6 and IL-8 in blood (Figures 6C, D). However, the double deletion mutant Δvat&tshPU-1 induced the significantly higher levels of IL-6 and IL-8 at the 6 h post-infection (no significant differences in blood bacterial loads compared with the wild-type at this point in time), indicating that a greater inflammatory response was activated compared with the wild-type infection. These data suggested that VatPU-1 and TshPU-1 coordinately attenuate the host immune responses, thus may mediate the evasion of ExPEC strain PU-1 from the immune clearance of blood leukocytes.
Figure 6 The assessment VatPU-1 and TshPU-1 in modulation of host immune responses during blood infection. (A, B) Blood routine testing detected the white blood cell (WBC) and neutrophil cell (NEU) counts of peripheral blood from the mice infected with indicated ExPEC strains at the 6 h post-infection. (C, D) Detection of IL-6 and IL-8 levels in bloods from the mice infected with indicated ExPEC strains at the 6 h post-infection. Statistical significance was determined by a one-way ANOVA test based on comparisons with the wild-type group (**P < 0.01).
DiscussionAlthough numerous virulence factors have been identified in diverse ExPEC pathotypes, the underlying mechanisms of ExPEC causing bloodstream infection is incompletely understood. ExPEC strain PU-1 showed a robust ability in survival within the blood, and this peculiarity may contribute to better show the full picture of ExPEC blood infection, suggesting that this strain deserves to be an excellent platform for further study.
In strain PU-1, the encoding genes of VatPU-1 and TshPU-1 were significantly upregulated in host blood but not in serum in vitro, suggesting that the contact with host blood cells may be one of the necessary conditions for their transcriptional activation. Furthermore, inactivation of VatPU-1 alone or TshPU-1 alone did not affect the bacterial survival in host blood, suggesting that these two SPATEs of strain PU-1 have obvious compensatory effects on each other, while they do not seem functionally redundant, as the encoding features of genes located in chromosome and plasmid may regulated by different mechanisms. Otherwise, the above analyses partially explain why we could not screen these two SPATEs as blood colonization factors in strain PU-1 through high-throughput technologies in previous studies (Ma et al., 2020; Ma et al., 2021). Although these studies have identified the carbon central metabolism, anaerobic respiratory chains, de novo biosynthetic pathways of nucleotide, extracellular polysaccharide biosynthesis and iron uptake playing critical roles for heavy bacterial load within bloodstream, the underlying mechanisms of strain PU-1 escaping from the host immune clearance via the interaction with the blood components (including RBCs, leukocytes, lymphocytes, et al.) were still incompletely understood. In this study, the roles of VatPU-1 and TshPU-1 interacting with hemoglobin, mucus and leukocytes during blood infection were revealed, which filled in the potential loopholes of our previous screening strategies.
When proliferating in host blood, bacteria not only have to resist the bactericidal effects of serum, but also evade from the clearance of immune cells (including diverse leukocytes and lymphocytes) (Huja et al., 2014; Ma et al., 2018). The mucin-like glycoproteins are widely present on the surface of leukocytes, and play critical roles for the immune modulation (Ruiz-Perez et al., 2011; Ruiz-Perez and Nataro, 2014). Several homologues of class-2 SPATE have been reported to efficiently cleave leukocyte surface glycoproteins involved in diverse immune functions (Ruiz-Perez et al., 2011; Ruiz-Perez and Nataro, 2014), such as the cleavages of Pic and Tsh homologues to CD43, CD44, CD45, CD93, CD162, PSGL-1, and fractalkine, while the contributions of these phenotypes to the bacterial blood infection in vivo have never been verified. Among the most abundant mucin-like leukocyte surface glycoproteins, CD43 is widely expressed on nearly all lineages of hematopoietic cells (Rosenstein et al., 1999). The adhesion of leukocytes to neighboring cells can be prevented by CD43, which facilitate the leukocytes’ migration in response to chemokine attraction, while extra processing in CD43 mediated by pathogens may disrupt this native function (Manjunath et al., 1993; Seveau et al., 2000). Indeed, several secreted mucinases from diverse pathotypes of E. coli have been confirmed to inhibit PMNs’ chemoattraction and function by cleavage of CD43 (Szabady et al., 2009; Ruiz-Perez et al., 2011). Our study identified the cleavages of CD43 by the homologues of Vat and Tsh from strain PU-1, which is consistent with the previous findings of Tsh homologue in APEC strain (Ruiz-Perez et al., 2011). Cleavage of these diverse substrates from leukocyte surface could result in paralysis of the leukocyte-mediated response for optimal bloodstream infection.
The interactions with pathogens are not only mediated by immune cells during blood infection, but also widely occur in RBCs and platelets. Mucin-like glycoproteins widely present on the surface of nearly all lineages of hematopoietic cells (Ruiz-Perez et al., 2011; Ruiz-Perez and Nataro, 2014), including RBCs. By virtue of their ability to adhere glycoproteins, both the secreted and surface located “Tsh/hbp” SPATEs were found to agglutinate RBCs and bind to hemoglobin and extracellular matrix proteins (such as collagen IV and fibronectin), and these interactions were independent of its protease activity (Provence and Curtiss, 1994; Stathopoulos et al., 1999; Kostakioti and Stathopoulos, 2004). Our data verified that both of VatPU-1 and TshPU-1 have the ability to bind to hemoglobin (Figure 4A), further works need to study the contributions of this phenotype to the bacterial blood infection. Several SPATEs have been reported to degrade the bound hemoglobin, and subsequently bind the released heme, thus contribute as an iron source generator (Otto et al., 1998; Otto et al., 2002). Numerous studies have confirmed the critical roles of iron uptake during bacterial infection within host bloodstream (Russo et al., 2001; Torres et al., 2001).
In summary, high virulent ExPEC strain PU-1 delivers two SPATEs to interact with the RBCs and blood leukocytes, which play critical roles to cause a heavy bacterial load within bloodstream by impairing PMNs’ functions via mucin-like O-glycoproteins’ cleavages. These findings provide a more comprehensive understanding how ExPEC colonizes within host bloodstream and causes severe sepsis.
Data availability statementThe original contributions presented in the study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding author.
Ethics statementThe animal study was reviewed and approved by Animal Research Committee Guidelines of Jiangsu Province (License Number: SYXK (SU) 2017-0007).
Author contributionsXP, JM, and HY: conceived and designed the experiments. XP and RC: performed the experiments. XP, YatZ, YinZ and JM: analyzed the data. JM and HY: reviewed the study. XP and JM: wrote the manuscript. All authors contributed to the article and approved the submitted version.
FundingThis work was supported by the National Key Research and Development Program of China (2022YFD1800904), and the National Natural Science Foundation of China (31802187).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1138801/full#supplementary-material
ReferencesDatsenko, K. A., Wanner, B. L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U.S.A. 97, 6640–6645. doi: 10.1073/pnas.120163297
PubMed Abstract | CrossRef Full Text | Google Scholar
Diekema, D. J., Pfaller, M. A., Jones, R. N., Group, S. P. (2002). Age-related trends in pathogen frequency and antimicrobial susceptibility of bloodstream isolates in north America: SENTRY antimicrobial surveillance program 1997-2000. Int. J. Antimicrob. Agents 20, 412–418. doi: 10.1016/S0924-8579(02)00204-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Ding, Y., Tang, X., Lu, P., Wu, B., Xu, Z., Liu, W., et al. (2012). Clonal analysis and virulent traits of pathogenic extraintestinal Escherichia coli isolates from swine in China. BMC Vet. Res. 8, 140. doi: 10.1186/1746-6148-8-140
PubMed Abstract | CrossRef Full Text | Google Scholar
Dutta, P. R., Cappello, R., Navarro-Garcia, F., Nataro, J. P. (2002). Functional comparison of serine protease autotransporters of enterobacteriaceae. Infect. Immun. 70, 7105–7113. doi: 10.1128/IAI.70.12.7105-7113.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Ejrnaes, K. (2011). Bacterial characteristics of importance for recurrent urinary tract infections caused by Escherichia coli. Dan Med. Bull. 58, B4187.
PubMed Abstract | Google Scholar
Gibold, L., Garenaux, E., Dalmasso, G., Gallucci, C., Cia, D., Mottet-Auselo, B., et al. (2016). The vat-AIEC protease promotes crossing of the intestinal mucus layer by crohn's disease-associated Escherichia coli. Cell Microbiol. 18, 617–631. doi: 10.1111/cmi.12539
PubMed Abstract | CrossRef Full Text | Google Scholar
Harrington, S. M., Sheikh, J., Henderson, I. R., Ruiz-Perez, F., Cohen, P. S., Nataro, J. P. (2009). The pic protease of enteroaggregative Escherichia coli promotes intestinal colonization and growth in the presence of mucin. Infect. Immun. 77, 2465–2473. doi: 10.1128/IAI.01494-08
PubMed Abstract | CrossRef Full Text | Google Scholar
Heimer, S. R., Rasko, D. A., Lockatell, C. V., Johnson, D. E., Mobley, H. L. (2004). Autotransporter genes pic and tsh are associated with Escherichia coli strains that cause acute pyelonephritis and are expressed during urinary tract infection. Infect. Immun. 72, 593–597. doi: 10.1128/IAI.72.1.593-597.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Henderson, I. R., Czeczulin, J., Eslava, C., Noriega, F., Nataro, J. P. (1999). Characterization of pic, a secreted protease of shigella flexneri and enteroaggregative Escherichia coli. Infect. Immun. 67, 5587–5596. doi: 10.1128/IAI.67.11.5587-5596.1999
PubMed Abstract | CrossRef Full Text | Google Scholar
Huja, S., Oren, Y., Biran, D., Meyer, S., Dobrindt, U., Bernhard, J., et al. (2014). Fur is the master regulator of the extraintestinal pathogenic Escherichia coli response to serum. MBio 5, e0146014. doi: 10.1128/mBio.01460-14
CrossRef Full Text | Google Scholar
Kostakioti, M., Stathopoulos, C. (2004). Functional analysis of the tsh autotransporter from an avian pathogenic Escherichia coli strain. Infect. Immun. 72, 5548–5554. doi: 10.1128/IAI.72.10.5548-5554.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Leyton, D. L., Sloan, J., Hill, R. E., Doughty, S., Hartland, E. L. (2003). Transfer region of pO113 from enterohemorrhagic Escherichia coli: similarity with R64 and identification of a novel plasmid-encoded autotransporter, EpeA. Infect. Immun. 71, 6307–6319. doi: 10.1128/IAI.71.11.6307-6319.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Ma, J., An, C., Jiang, F., Yao, H., Logue, C., Nolan, L. K., et al. (2018). Extraintestinal pathogenic Escherichia coli increase extracytoplasmic polysaccharide biosynthesis for serum resistance in response to bloodstream signals. Mol. Microbiol. 110, 689–706. doi: 10.1111/mmi.13987
PubMed Abstract | CrossRef Full Text | Google Scholar
Ma, J., Cheng, Z., Bai, Q., Zhao, K., Pan, Z., Yao, H. (2021). Screening virulence factors of porcine extraintestinal pathogenic Escherichia coli (an emerging pathotype) required for optimal growth in swine blood. Transbound Emerg. Dis. 68, 2005–2016. doi: 10.1111/tbed.13848
PubMed Abstract | CrossRef Full Text | Google Scholar
Ma, J., Pan, X., Zhong, X., Bai, Q., Liu, G., Yao, H. (2020). Preferential use of carbon central metabolism and anaerobic respiratory chains in porcine extraintestinal pathogenic Escherichia coli during bloodstream infection. Vet. Microbiol. 249, 108830. doi: 10.1016/j.vetmic.2020.108830
PubMed Abstract | CrossRef Full Text | Google Scholar
Manjunath, N., Johnson, R. S., Staunton, D. E., Pasqualini, R., Ardman, B. (1993). Targeted disruption of CD43 gene enhances T lymphocyte adhesion. J. Immunol. 151, 1528–1534. doi: 10.4049/jimmunol.151.3.1528
PubMed Abstract | CrossRef Full Text | Google Scholar
Navarro-Garcia, F., Gutierrez-Jimenez, J., Garcia-Tovar, C., Castro, L. A., Salazar-Gonzalez, H., Cordova, V. (2010). Pic, an autotransporter protein secreted by different pathogens in the enterobacteriaceae family, is a potent mucus secretagogue. Infect. Immun. 78, 4101–4109. doi: 10.1128/IAI.00523-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Otto, B. R., Van Dooren, S. J., Dozois, C. M., Luirink, J., Oudega, B. (2002). Escherichia coli hemoglobin protease autotransporter contributes to synergistic abscess formation and heme-dependent growth of bacteroides fragilis. Infect. Immun. 70, 5–10. doi: 10.1128/IAI.70.1.5-10.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Otto, B. R., Van Dooren, S. J., Nuijens, J. H., Luirink, J., Oudega, B. (1998). Characterization of a hemoglobin protease secreted by the pathogenic Escherichia coli strain EB1. J. Exp. Med. 188, 1091–1103. doi: 10.1084/jem.188.6.1091
PubMed Abstract | CrossRef Full Text | Google Scholar
Provence, D. L., Curtiss, R., 3rd (1994). Isolation and characterization of a gene involved in hemagglutination by an avian pathogenic escherichia coli strain. Infect. Immun. 62, 1369–1380. doi: 10.1128/iai.62.4.1369-1380.1994
PubMed Abstract | CrossRef Full Text | Google Scholar
Ruiz-Perez, F., Nataro, J. P. (2014). Bacterial serine proteases secreted by the autotransporter pathway: classification, specificity, and role in virulence. Cell Mol. Life Sci. 71, 745–770. doi: 10.1007/s00018-013-1355-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Ruiz-Perez, F., Wahid, R., Faherty, C. S., Kolappaswamy, K., Rodriguez, L., Santiago, A., et al. (2011). Serine protease autotransporters from shigella flexneri and pathogenic Escherichia coli target a broad range of leukocyte glycoproteins. Proc. Natl. Acad. Sci. U.S.A. 108, 12881–12886. doi: 10.1073/pnas.1101006108
PubMed Abstract | CrossRef Full Text | Google Scholar
Russo, T. A., Carlino, U. B., Johnson, J. R. (2001). Identification of a new iron-regulated virulence gene, ireA, in an extraintestinal pathogenic isolate of Escherichia coli. Infect. Immun. 69, 6209–6216. doi: 10.1128/IAI.69.10.6209-6216.2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Seveau, S., Keller, H., Maxfield, F. R., Piller, F., Halbwachs-Mecarelli, L. (2000). Neutrophil polarity and locomotion are associated with surface redistribution of leukosialin (CD43), an antiadhesive membrane molecule. Blood 95, 2462–2470. doi: 10.1182/blood.V95.8.2462
PubMed Abstract | CrossRef Full Text | Google Scholar
Silva, A. J., Pham, K., Benitez, J. A. (2003). Haemagglutinin/protease expression and mucin gel penetration in El tor biotype Vibrio cholerae. Microbiol. (Reading) 149, 1883–1891. doi: 10.1099/mic.0.26086-0
CrossRef Full Text | Google Scholar
Stathopoulos, C., Provence, D. L., Curtiss, R., 3rd (1999). Characterization of the avian pathogenic escherichia coli hemagglutinin tsh, a member of the immunoglobulin a protease-type family of autotransporters. Infect. Immun. 67, 772–781. doi: 10.1128/IAI.67.2.772-781.1999
留言 (0)