
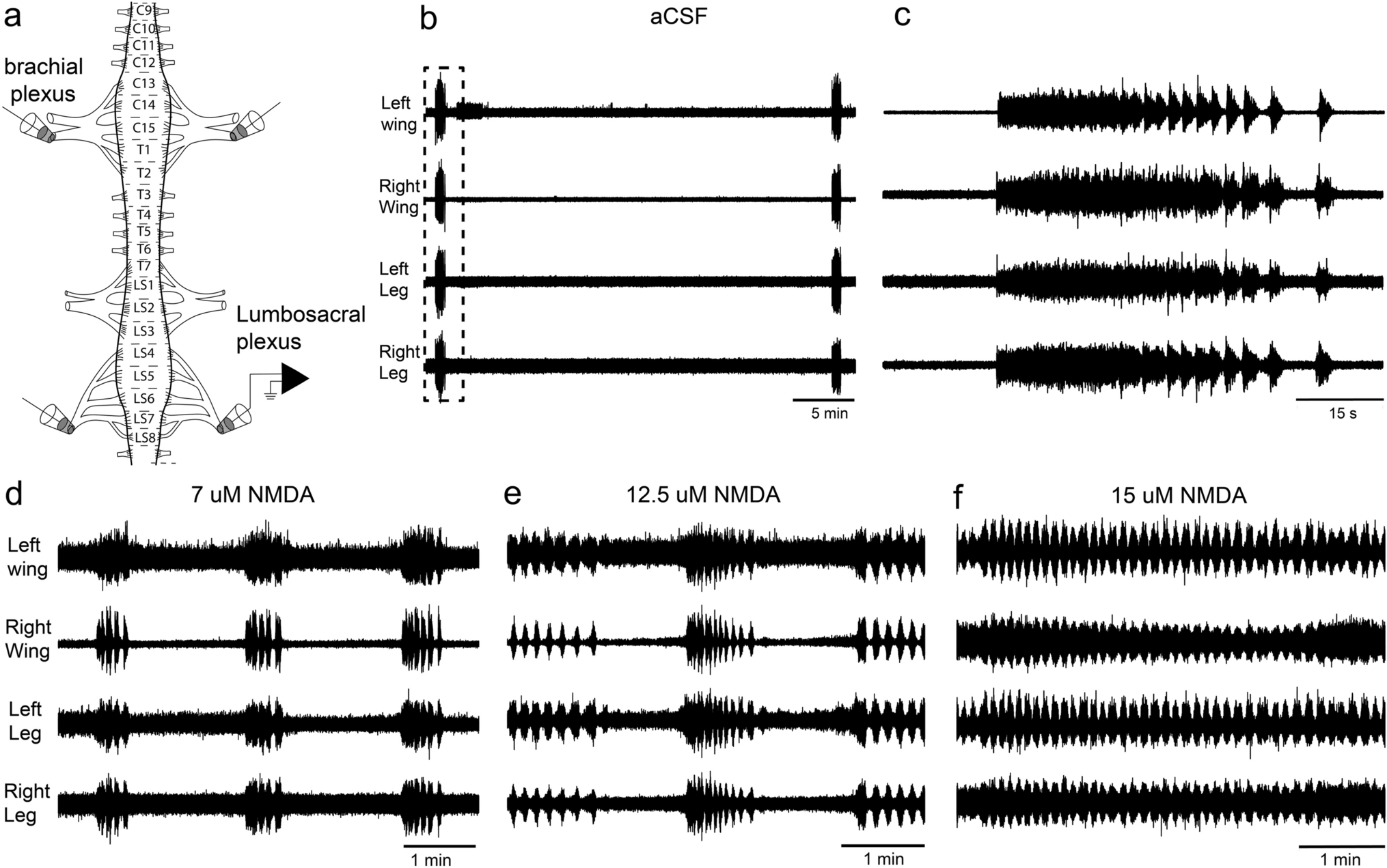
記住我








To highlight the variety of CRY/PL subfamilies and classify insect CRY/PLs in their general phylogeny, we used the unrooted phylogenetic tree generated in Deppisch et al. (2022) and added 787 insects CRY/PL sequences from the present study. The former tree contained the sequences from maximally five species per order and two species per family, among which were just 125 insect sequences, making the tree phylogenetically quite balanced (Fig. 1a; Deppisch et al. 2022). The new tree contains 912 insect sequences (from 340 insect species), which were highlighted in color, and 2124 non-insect sequences (Fig. 1b). Due to the high number of insect sequences, this tree is not balanced and only of limited use for phylogenetic statements classification. However, when comparing it with the original tree in Deppisch et al. (2022) (Fig. 1a), it clearly demonstrates that insects possess mainly members of the subfamilies MCRY (red), 6-4 photolyase (pink), DCRY (blue), and CPDII photolyase (light green), while the other subfamilies are underrepresented (Fig. 1a). None of the insect sequences clusters with the plant photolyase (PPL, ochre green), plant CRY (dark green), the marine organism specific PCRY-like (turquoise), the CPDIII photolyase (brown), the chordate-specific CRY4 cluster (purple) and the cnidarian-specific ACRY cluster (dark blue). Also, CPDI (dark brown) and DASH-CRY (orange) hardly occur in insects. Only three insect sequences cluster with DASH-CRY and two with CPDI photolyases. Five sequences do not cluster with any known CRYPL subfamily and remain unexplored (dark grey in Fig. 1b). These belong to the cat flea Ctenocephalides felis, the bug Apolygus lucorum, and several walking stick species (Timema).
Fig. 1
Unrooted phylogenetic trees representing all major CRY/PL subfamilies. a Unrooted tree representing the known CRY/PL subfamilies (adapted from Deppisch et al. 2022). ACRYs (dark blue), DCRYs (blue), CRY4s (purple), 6-4 PLs (pink), and MCRYs (red) belong to the 6-4 PL cluster (I). The CPDII PLs (light green) are the only CRY/PLs in the CPDII PL cluster (II). The CPDI (dark brown) as well as CPDIII photolyases (light brown), the plant cryptochrome PCRY (dark green) and the PCRY-like (turquoise) belong to the CPDI/III cluster (III). DASH-CRYs (orange) and the plant photolyase PPLs (ochre green) belong to the DASH-CRY cluster (IV). b Unrooted tree to which all so far sequenced and annotated insect CRY/PL sequences were added and shown in color. Insect-specific sequences were present in the CPDII PL, 6-4 PL, MCRY, and DCRY clusters. A small number of insect sequences clustered with DASH-CRY and CPDI PLs, while few sequences (dark grey) were not assignable to the known CRY/PLs
Most insect CRY/PLs are members of the CPDII photolyase cluster, followed by the MCRY cluster. DCRY and 6-4 photolyases are clustered about equally. Interestingly, some insects (Ischnura elegans, Ladona fulva, and all flies of the superfamily Tephridoidea) appear to possess two CPDII photolyases. While repeated annotations and protein isoforms usually cluster together in the phylogenetic tree, these duplicates were located nearby but in different branches (Supplementary files 4 and 5).
Motif analysisTo verify the accuracy of the phylogenetic tree, we performed a motif analysis using the Annotate & Predict tool in Geneious Prime software, applying all previously known CRY/PL motifs summarized by Deppisch et al. (2022). We found that almost all identified CRY/PLs exhibit their typical subfamilial motifs (Fig. 2). All protein sequences with their protein motifs are listed in Supplementary file 7. Based on the motif analysis, we could assign the three unclassifiable sequences from walking sticks (Timema spec.) to DCRY. Apparently, they were annotated as smaller fragments, but all possess DCRY specific motifs (Supplementary file 7). Protein sequence alignment of all putative Timema DCRY sequences compared to DCRY from Drosophila melanogaster (Supplementary file 8) showed that the different fragments belong to the same DCRY sequence. Therefore, we added these Timema sequences to DCRY in Fig. 2. The unclassifiable sequence from cat flea Ctenocephalides felis (XP_026474410.1) carries 6-4 PL cluster-typical motifs that are common in 6-4 PLs as well as in MCRYs and DCRYs. As we cannot assign it more precisely, we did not include it in Fig. 2 and disregarded it for further analyses. We did the same for the unclassified sequence of the bug, Apolygus lucorum (KAF6198968.1), because we did not find any CRY/PL motifs, suggesting that this sequence is not a CRY/PL or at least not a hitherto known CRY/PL.
Fig. 2
Schematical presentation of the insect CRY/PL subfamilies with their most frequent motifs. Only motifs that occur with a frequency of at least 30% are shown. We used the same motifs and search parameters as described in Deppisch et al. (2022). All protein sequences including their motifs are listed in Supplementary file 7. The number of sequences studied is indicated above the subfamily name. In the case of DCRY of walking sticks (Timema spec.), we included sequences that clustered differently but had DCRY motifs. In the case of the cc3 motif of CPDII PL (*), both plant (9%) and microorganism/animal (69%) motifs were combined
Due to the large number of sequences examined, we can make precise statements about the occurrence and frequency of individual motifs in CPDII PL, 6-4 PL, MCRY, and DCRY, while we cannot do so for the rarely occurring CRY/PLs (DASH-CRY and CPDI-PL) (Fig. 2). In the following, we will describe the distribution of the individual motifs in more detail.
CPDII photolyases are characterized by the alpha helix motif α10, the CPDII-PL-specific motifs cc1 (CPDII conserved1), cc2 (CPDII conserved2), cc3 (CPDII conserved3), and mts (mitochondrial target sequence) (Fig. 2). The α10 motif is very abundant in all CRY/PLs except for the CPDI photolyases, which have completely different motifs. The 6-4 photolyases are characterized by several additional alpha helixes (α8, α10, α12, α14, α15, α16, α17, and α18), the sulfur loop (sl), and the plant n-terminal (pn) motifs. These motifs are very common not only in the 6-4 photolyase itself, but also in MCRY and DCRY, which are derived from it. In addition, the 6-4 photolyases carry the 6-4 motif (6-4) and the MCRY specific protrusion loop (ml) (Fig. 2). The 6-4 motif is only found with a frequency of 44% in insect 6-4 photolyase, whereas it is almost twice as frequent (81%) in other organisms (Deppisch et al. 2022). In contrast, the MCRY specific protrusion loop (ml) occurs in 38% of insect 6-4 photolyases while it was only present in 17% of all 6-4 photolyases studied in Deppisch et al. (2022) and was, therefore, not considered. The ml motif is in different places in 6-4 photolyases and MCRYs (Fig. 2): in 6-4 photolyases, it is usually located at the C-terminal end, while it is between the alpha helices α10 and α11 in MCRY. Exceptions are the 6-4 PLs of the cotton bag worm Eumeta japonica where it also occurs between α10 and α11, that of the marine midge Clunio marinus where the motif occurs twice in succession between α10 and α11, and that of Drosophila innubila, where it occurs in the N-terminus between plant N-terminus (pn) and α10 (Supplementary file 7). Drosophila innubila is the only Muscomorpha in which the ml motif occurs. The ml motif is generally rare in flies (Brachycera). We found another ml motif in the 6-4 PL of black soldier flies (Hermetia illucens), and there it occurs at the C-terminal end, as in all other insects (Supplementary files 6 and 7). The 38% frequency of the ml motif in Fig. 2 was calculated regardless of its position.
MCRYs carry an additional alpha helix, α11, besides the already mentioned alpha helices of 6-4 photolyases (Fig. 2). With a percentage of 93%, α11 is very frequent in MCRYs while it occurs in only 13% of the 6-4 PLs and is, therefore, not included in Fig. 2. Furthermore, MCRYs possess the MCRY-specific motifs m2pu (MCRY 2nd pocket up), m2pl (MCRY 2nd pocket low), mpm (MCRY phosphate motif), mnc (MCRY-non-chordates), and the already mentioned protrusion loop, ml. Similarly, DCRYs are characterized by the additional DCRY motifs d2pu (DCRY 2nd pocket up), d2pl (DCRY 2nd pocket low), dm1 (DCRY motif1), and dcl (DCRY c-terminal lid).
In DASH-CRY sequences, we find in addition to the alpha helices 8 and 10, the DASH-CRY-specific dlr (CPD lesion repair), db1 (D-R-salt bridge 1), and db2 (D-R-salt bridge 2), as well as the c3 motif (CPDIII conserved). In the two CPDI photolyases, the c1/3-1 (CPDI/III conserved1) and c3 motifs are present. Both motif analyses reveal that the DASH-CRYs and CPDI photolyases found in insects do indeed belong to these subfamilies and are not resulting from incorrect clustering in the phylogenetic tree.
CRY/PL distributionTo obtain an overview of the distribution of CRY/PL family members in the different insect groups, we subdivided the individual insects into orders and superfamilies (if subdivision into superfamilies was not given, into families) and examined the CRY/PL distribution in these (Fig. 3). As already evident in the unrooted tree (Fig. 1), MCRY, DCRY and the two photolyases 6-4 and CPDII are the CRY/PLs most abundant in insects. In the following, we will refer to them as the four main insect CRY/PLs or even as the four CRY/PLs. Nevertheless, the four CRY/PLs are not present in all insects. Furthermore, few insects possess in addition or instead the DASH-CRY and CPDI photolyases, and few other insects carry a CPDII duplication (Fig. 3).
Fig. 3
Distribution of CRY/PL subfamilies within insect orders and superfamilies (MCRY dark red, DCRY blue, 6-4 PL light magenta, CPDII light green, CPDII duplication green, CPDI brown, DASH-CRY orange). The prevalence of a CRY/PL within an insect superfamily is shown as a fraction, with the denominator representing the total number of animals examined in a superfamily and the numerator representing the number of animals with the respective CRY/PL. MCRY, DCRY, 6-4 PL, and CPDII PL are the most common CRY/PLs in insects. All studied insects belonging to the orders Ephemeroptera, Odonata, Phasmatodea, Thysanoptera, and Neuroptera possess them. Most Lepidoptera and many insects belonging to Hemiptera (depending on the superfamily) also have all of them. In Hymenoptera, the CRY/PLs are greatly reduced, and some groups retain only MCRY. Such reductions are also observed in insects belonging to other orders. Many superfamilies of Diptera lack MCRY and possess only DCRY, 6-4 PL, and CPDII PL. Some individual insects also have DASH-CRY or CPDI photolyase and others possess a CPDII photolyase duplication. DCRY is only fragmentarily sequenced and annotated in most Timematoidea (*). But in all studied Timema species we could find DCRY fragments
All four CRY/PLs in PalaeopteraThe mayflies (order: Ephemeroptera) and dragonflies (order: Odonata) belong to the more primitive infraclass Palaeoptera (Fig. 5). A characteristic of this infraclass is the inability of the insects to fold their wings over the abdomen as the insects of the Neoptera do. In our analysis, we found only two mayflies and two dragonflies with a sequenced and annotated genome. All of them contain the four insect CRY/PLs: MCRY, DCRY, 6-4 and CPDII photolyases (Fig. 3). The two dragonflies, Ischnura elegans and Ladona fulva, appear furthermore to possess a CPDII duplication (Fig. 4). While the blue-tailed damselfly Ischnura elegans, clearly has two CDPII photolyases (CPDIIa and CPDIIb), this is less certain in the scarce chaser Ladona fulva due to incomplete sequences (Fig. 4, Supplementary file 8).
Fig. 4
Protein sequence alignment of CPDII photolyase duplicates from Tephritoidea with Drosophila melanogaster sequences indicates a CPDII PL duplication in Tephritoidea and dragon flies resulting in CPDIIa and CPDIIb sequences. a All Tephritoidea sequences lack the CPDII antenna binding (cab) motif that is present in Drosophila. The duplicates b additionally lack the cc1 (1) motif and some b duplicates lack also the mitochondrial targeting motif (m) of CPDII. b Protein sequence alignment of the two CPDII photolyases from Ischnura elegans and Ladona fulva compared with the CPDII PL from Drosophila melanogaster. While XP_046383944.1 has a cab motif but no cc1 motif, XP_046383389.1 carries a cc1 motif and no cab motif. The two Ladona sequences are incomplete and therefore can be derived from the same sequence as well as from two different CPDII-PLs
All other insects studied belong to the Infraclass Neoptera that can fold their wings over the abdomen and that can be further divided into Polyneoptera (orders: Orthoptera, Phasmatodea, and Blattodea), Paraneoptera (orders: Hemiptera, Thysanoptera, and Phthiraptera), and Endopterygota (orders: Hymenoptera, Neuroptera, Coleoptera, Diptera, and Lepidoptera) (Fig. 5). In the following, we will describe the CRY/PL member composition in the different orders briefly.
Fig. 5
Distribution of CRY/PL subfamilies in hexapoda down to the resolution of certain insect species in order to demonstrate the loss and rare reappearance of CRY/PL family members. The total number of animals examined within a taxon (denominator) and the number of animals with CRY/PL distribution shown (numerator) are given in parentheses next to the taxonomic name. Data labeled 1 (in superscript) are from the publication by Kotwica-Rolinska et al. (2021) and data labeled 2 (in superscript) are from Deppisch et al. (2022). The ancestors of insects probably had five members of the CRY/PL family-MCRY, DCRY, 6-4 PL, CPDII PL, and DASH-CRY. The more primitive hexapods, which belong to the class Collembola, possess these five CRY/PLs. However, DASH-CRY is lost in most insects. The insects of the superfamily Sciaroidea and Aleyrodoidea again possess DASH-CRYs. In Zygotera (suborder) and Tephritoidea (superfamily) we see a CPDII duplication. In Epiprocta (suborder), we cannot yet confirm this duplication. Other CRY/PL members of the insects have been lost at different phylogenetic levels. In the Hymenoptera, DCRY and 6-4 PL have already been lost at the order level. CPDII has been lost only at the level of the superfamily Formicoidea. Similarly, MCRY has been lost in the Muscomorpha only in some superfamilies
OrthopteraWe examined nine species belonging to the order Orthoptera, three of which belong to the suborder Grilloidea (crickets) and are not considered in the final calculation as their genomes are not fully sequenced (Supplementary file 9). All other six Orthoptera belong to the suborder Acrididae (locusts) and possess all four CRY/PLs (Fig. 3).
PhasmatodeaThe nine species sequenced belong all to the already discussed walking sticks (Timematoidea). The majority of these species appear to possess all 4 CRY/PL members. Only the CPDII photolyases were found in 67% of the species (Fig. 3). Since the Timema sequences are more likely to be fragments and, therefore, have smaller e-values than complete sequences, their CPDII PLs may have been missed in the BLASTP analysis with our applied parameters.
BlattodeaThough we found a total of eight Blattodea in our analysis, only four of them are fully sequenced and were considered for further analyses. Of these only the German cockroach Blattella germanica possesses DCRY and CPDII photolyase in addition to MCRY (Fig. 3). The other three sequenced termites seem to retain only MCRY (Fig. 3).
ThysanopteraThe two thrips examined again possess all four CRY/PLs.
HemipteraThe eight investigated superfamilies of the order Hemiptera show a heterogeneous CRY/PL composition (Fig. 3). While nearly all species of the suborder Sternorrhyncha (consisting of the superfamilies Aleyrodoidea, Psylloiidae, Aphidoidea, Phylloxeroidea, and Membracoidea) have all four CRY/PLs, species from the other suborders (Auchenorrhyncha and Heteroptera) have significantly fewer (Fig. 3). The whitefly Bemisia tabaci from the superfamily Aleyrodoidea has an additional DASH-CRY. The Cimicoidea and Fulgoroidea lack the 6-4 photolyase, and within the Cimicoidea, the bed bug, Cimex lectularis lacks also DCRY. The same reduction (no 6-4 photolyase and no DCRY) is found in the Pentatomoidea. Although the fire bug from the superfamily Pyrrhocoris apterus was not included in our analysis due to incomplete sequencing (Supplementary file 6, 10), it also appears to possess MCRY and CPDII PL only (Kotwica-Rolinska et al. 2021).
PhirapteraThe body louse Pediculus humanus corporis is the only Phthiraptera studied and possesses only MCRY (Figs. 3, 5).
HymenopteraOf all the orders studied, the order Hymenoptera is probably the most conserved. All Hymenoptera examined lack DCRY and the 6-4 photolyase (Fig. 3). Almost all of them (59 out of 60), except those belonging to the superfamily Formicoidea (ants), carry the MCRY and the CPDII photolyase (Figs. 3, 5, Supplementary file 6). All 30 ants examined lost the CPDII photolyase and retain only MCRY (Fig. 5).
NeuropteraThe common green lacewing Chrysoperla carnea, the only Neuroptera studied, again possesses all four CRY/PLs.
ColeopteraMost insects of the order Coleoptera (beetles) have rather a reduced CRY/PL composition (Fig. 3). The only exception is the emerald ash borer Agrilus planipennis (superfamily: Buprestoidea), which carries all four CRY/PLs. All other beetles studied have MCRY as their major CRY/PL family member and in most beetles, this is also the only one. Many beetles of the superfamily Chrysomeloidea (8/9) and all of the superfamily Cucujoidea (4/4) additionally possess the CPDII photolyase (Supplementary file 6). The firefly Abscondita terminalis (formerly also called Luciola terminalis; superfamily Elateroidea) is exceptional because it possesses a CPDI photolyase and no CPDII photolyase besides MCRY. (Figs. 3, 5). CPDI photolyases are rarely found in insects but are abundant in bacteria. Since fireflies are known to have an extensive microbiome because microbial endosymbionts such as Tenericutes spec. contribute to the metabolism and biosynthesis of luciferin (Fallon et al. 2018).Thus, it is quite likely that the found CPDI is a contamination from them.
DipteraDiptera are classically divided into Nematocera and Brachycera (Fig. 5). Of the Nematocera, 18 species belonging to the infraorder Culicomorpha and three species belonging to the infraorder Bibionomorpha (superfamily Sciaroidea) have been sequenced (Fig. 3). The latter three are all gnats (fungus or turnip gnats) having a special position among all diptereans. The gnats lack DCRY, while most Culicomorpha possess the main insect CRY/PLs. Furthermore, the two fungus gnats Bradysia coprophila & Bradysia odoriphaga possess DASH-CRY. B. odoriphaga additionally seems to possess a CPDI photolyase, However, since this clusters with the CPDI photolyases of bacteria, including those of Escherichia coli and since the sister species Bradysia coprophila does not have any CPDI photolyase, we assume that it is more likely a bacterial contamination of the sequenced DNA.
Of the Brachycera, 67 species of the infraorder Muscomorpha and 1 species of the infraorder Stratiomyomorpha (the black soldier fly Hermetia illucens) have been sequenced (Fig. 5). While the black soldier fly possesses all four CRY/PL members, all sequenced species from the Muscomorpha have lost MCRY (Figs. 3, 5). The following superfamilies belong to the Muscomorpha: Oestoidea, Muscuoidea, Hippoboscoidea, Ephydroidea, Diopsoidea and Tephritoidea (Fig. 3). They all have DCRY as their main cryptochrome and the great majority have in addition the photolyases 6-4 and CPDII (Fig. 3). The CPDII photolyase was not detected in the fly Drosophila immigrans (superfamily Ephydroidea), but since it is present in its sister species D. albomicans, this speaks for an incomplete annotation. Thus, we can assume that 6-4 and CPDII photolyases are likely present in all Muscomorpha. The only flies that appear exceptional are the Tephritoidea (9 sequenced species), which have two CPDII photolyases, the CPDIIa and CPDIIb duplicates (Figs. 3, 5).
LepidopteraAlmost all lepidopterans examined possess the four CRY/PLs. However, in some species one or the other member is missing. In some cases, this is probably due to sequencing and annotation errors.
Protein and nucleotide sequence alignments of CPDII to verify gene duplicationsTo verify whether the CPDII PL duplicates found in the fruit flies of the superfamily Tephritoidea and the two dragon flies (Odonata) were gene duplications or alternatively spliced mRNAs, we aligned their protein sequences with the CPDII photolyase of Drosophila melanogaster (Fig. 4).
In the nine sequenced Tephritiodea, both duplicates differ mainly in their CPDII conserved1 motif (cc1, Fig. 5a), which consists of the amino acid sequence EEAVVRREL. A cc1 motif is only present in the CPDIIa sequence but absent in its CPDIIb duplication (Fig. 4a). Furthermore, some CPDIIb duplicates lack in addition the CPDII mitochondrial target motif (mts), which consists of the sequence IHGFLRMYWAK (Fig. 4a). Nucleotide alignments of the CPDIIa and CPDIIb duplicates of the different species further confirm that they are true duplicates because the associated nucleotides of each duplicate align better between the species than between each other (Supplementary file 8). For example, the B. dorsalis CPDIIa sequence XP_011207519.3, aligns better with the B. oleae CPDIIa sequence XP_014095123.2 than with the B. dorsalis CPDIIb duplicate XP_011207520.2. In addition, we analyzed the coding regions of the photolyases CPDIIa and CPDIIb of B. dorsalis for their composition and found that both sequences are located directly adjacent to each other on chromosome 3. This shows that these are actual duplications and not isoforms of a gene where they would share the same gene span.
Also, in the case of the blue-tailed damselfly Ischnura elegans, we searched for the coding regions of CPDIIa and CPDIIb photolyases on their assembly. In this case, both photolyases are found inverted on the second chromosome more than 30 million base pairs apart, proving that they are true duplicates. These two protein sequences differ from each other not only in the cc1 motif, but additionally in the CPDII antenna binding (cab) motif, which is present in XP_046383389.1, but not in XP_046383944.1 (Fig. 4b). In the dragonfly Ladona fulva, the situation looks similar but is less clear because the two Ladona sequences found are incomplete. While KAG8236888.1 lacks the C-terminal end, KAG8231874.1 seems to consist only of this (Fig. 4b). Therefore, we cannot completely exclude that these are only two partial sequences from a single CPDII photolyase. All alignments are shown in more detail in Supplementary file 8.
A closer look at rarely found CRY/PLs in insectsOur study discovered three DASH-CRYs and two CPDI PLs that are rarely found in insects. By further investigation of their protein and nucleotide sequences, we aimed to determine whether they were true insect genes, possibly the result of HGT (Horizontal Gene Transfer), or contaminants. Interestingly, both CPDI PLs are derived from predictions of whole genome shotgun analyses, and both seem to map in the microbiomes of both species. Thus, the whole genome shotgun sequence JABVZW010000398 from which the Abscondita CPDI PL is derived has 13 other protein sequences. All these protein sequences yielded results from the bacterium Acinetobacter in addition to Abscondita in BLASTP searches. Since Acinetobacter is a component of the microbiome of the firefly Photuris versicolor, we assume that it is also present in the microbiome of Abscondita terminalis. Similarly, the whole genome shotgun sequence of Bradysia odoriphaga has two more protein sequences, all of them showing hits from the bacterium Serratia in addition to Bradysia in a BLASTP analysis. Serratia marcescens is an intestinal microorganism that has already been identified in insects. Therefore, we conclude that with certainty neither Bradysia nor Abscondita, and thus probably also all insects, do not possess CPDI photolyase (Fig. 5).
The Bradysia odoriphaga DASH-CRY was also predicted by whole genome shotgun analyses (JAFDOW010000828.1). In addition to the DASH-CRY protein, 68 other proteins were identified in the same fragment. BLASTP analyses of the other proteins revealed insect hits in addition to Bradysia itself, suggesting that these are in fact genes present in the Bradysia genome. BLASTP analyses of DASH-CRY revealed DASH-CRY results from other arthropods (Orchesella, Allacma, Daphnia, Nematostella, etc.) in addition to the two Bradysia DASHs. In the case of Bradysia coprophila DASH, As the chromosome in which the gene is localized is also known (Chr4), this additionally is, a clear indication for no microbiome contamination. Again, BLASTP analysis of Bradysia coprophila DASH-CRY revealed similar results as that of Bradysia odoriphaga and thus fits our phylogenetic tree (Table 1).
Table 1 COUSIN59 scores of CRY/PL nucleotide sequences of Bemisia tabacci are givenWe used the Codon Usage Similarity Index Cousin59 (https://cousin.ird.fr/) to evaluate whether DASH-CRY from Bemisia tabacci had a specific codon usage preference that could indicate HGT (Bourret et al. 2019). Because we did not know the exact donor of DASH-CRY, we compared its nucleotide sequence with the CUPrefs (Codon Usage Preferences) of Bemisia itself (generated via https://cousin.ird.fr/) and with those of the yellow spruce Xanthoria parietina, as well as with those of the tobacco plant Nicotiana tabacum (available via https://www.kazusa.or.jp/codon/). Nicotina tabacum is one of the major hosts of Bemisia tabacci and thus the potential candidate host, while the DASH-CRYs of several Xanthoria species are similar to the DASH-CRYs of Bemisia according to BLASTP. We compared the distribution of Cousin59 indices of all cryptochrome genes of Bemisia. We detect negative Cousin59 values when comparing the CRY/PL sequences with CUPrefs of Xanthoria. Consequently, we tend to exclude Xanthoria as a donor. The highest Cousin59 values are obtained when comparing the sequences with the CUPref of Bemisia, whereby the Cousin59 values with Nicotiana are also quite high. The Cousin59 values of DASH-CRY are all three times rather towards 0. Therefore, we assume that Bemisia DASH-CRY does not have a unique CUPref, although it is closest to Bemisia CUPrefs.
留言 (0)