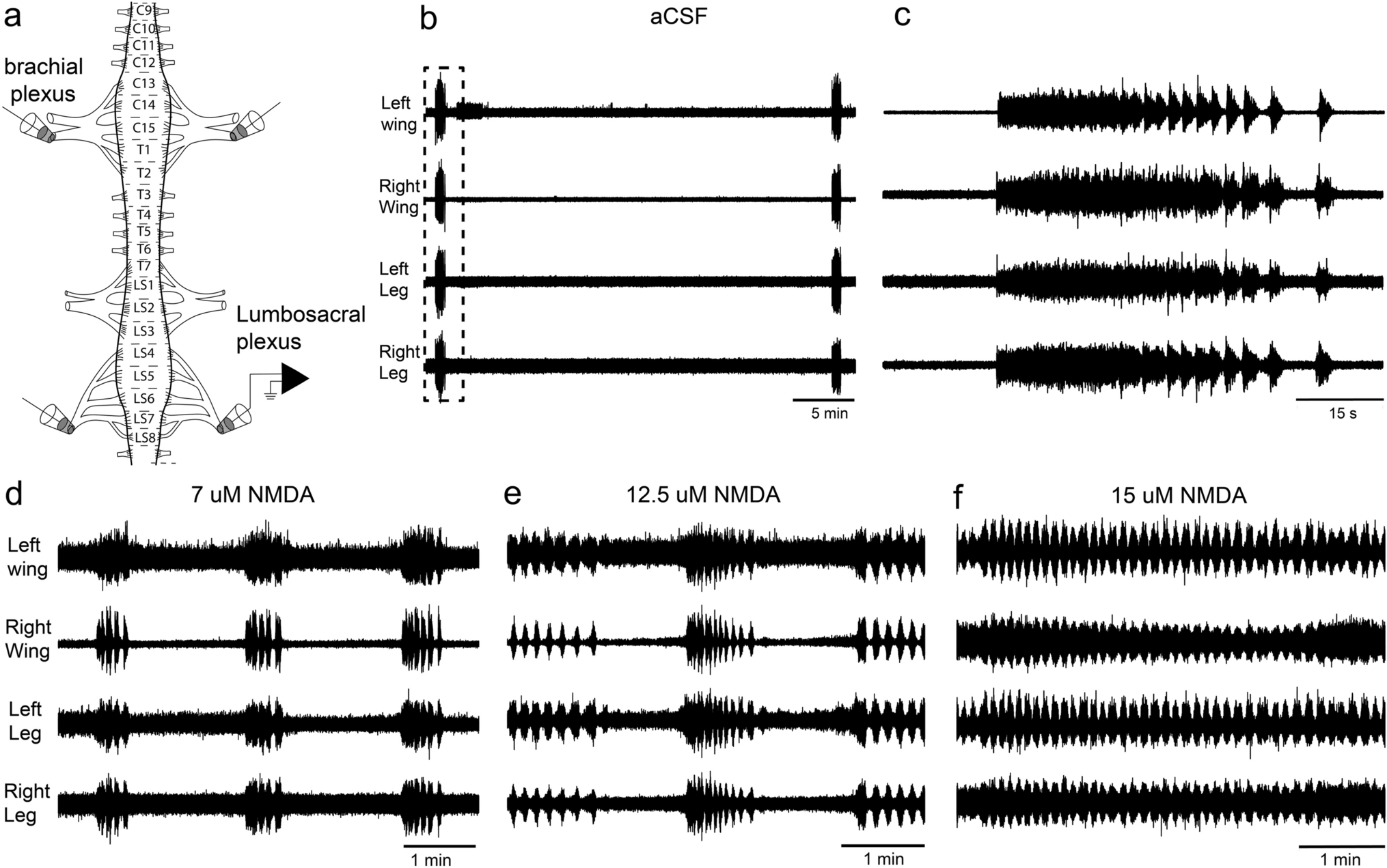
Here we have shown that, under a variety of conditions, the chick spinal cord produces, in vitro, rhythmic outputs that are coordinated bilaterally and between the wing and leg segments. This coordination occurs spontaneously and under a variety of agonist and antagonists, including NMDA, 5-HT and dopamine (Figs. 1, 2). Previous studies in the in vitro chick spinal cord have shown that rhythmic activity is coordinated between different segments of the lumbosacral spinal cord, and between the brachial and lumbosacral segments (Ho and O’Donovan 1993), but to the best of our knowledge, there has been no study looking at the bilateral coordination in in vitro preparations. Most previous studies in the chick spinal cord have focused on the spontaneous activity generated at early stages and the role this has in the development of CPGs (e.g. O’Donovan and Landmesser 1987; O’Donovan et al. 1992; O’Donovan 1999). Interestingly, we found that serotonin (5-HT) alone is not capable of initiating a rhythmic motor output in the chick spinal cord but does modulates the rhythmic pattern (Fig. 2). This is similar to what occurs in lamprey in vitro spinal cord preparations, where fictive locomotion cannot be initiated by serotonin alone but also acts a modulator of the pattern in the presence of NMDA (Harris-Warrick and Cohen 1985). In contrast, in neonatal mice and rats, serotonin alone is capable of initiating rhythmic locomotor activity in vitro, including alternation between left and right sides (Cazalets et al. 1992; Beato et al. 1997; Branchereau et al. 2000). Serotoninergic projections from the brainstem to the spinal cord have a well-established role in modulating locomotion in all vertebrates (Harris-Warrick and Cohen 1985; McDearmid et al. 1997; Schmidt and Jordan 2000; Gabriel et al. 2009; Flaive et al. 2020). In mammals, serotonin can initiate locomotion in neonates but not in adults (reviewed in Sławińska and Jordan 2019). It is possible that the differences in the role of serotonin in initiating locomotion are related to the different developmental modes of rodents and chickens. Chickens are precocial and can walk almost immediately after hatching, while rats, for example, are altricial and only start walking 12–13 days after birth (Muir 2000). Thus, it is possible that in precocial birds, there is no developmental change in the role of serotonin in modulating locomotion.
Differences between brachial and lumbosacral locomotor output
Despite finding bilateral coordination in the brachial and lumbosacral segments, we also found that even at early embryonic stages (E11–E14), the lumbosacral (legs) and the brachial (wing) segments show some differences in their locomotor output in response to the same pharmacological treatment (e.g. Figs. 3i, 4e–g, 5). This provides support to the idea that in the avian spinal cord the circuits of the CPGs that produce the motor outputs of wigs and legs are wired differently to produce distinct motor outputs; coordination with the wings and alternations with the legs (Gatesy and Dial 1996). The first evidence in this regard comes from early transplantation experiments in chick embryos, where the brachial segments were replaced by lumbosacral segments, which resulted in alternating movements of the wings (Straznicky 1963). In a similar experiment, transplantation of brachial segments to the lumbar cord results in synchronous leg movements (Narayanan and Hamburger 1971). Recently, Haimson et al., (2021) showed that in the chick brachial spinal cord there is a larger number of excitatory interneurons crossing the midline compared to the lumbosacral segments, and this is related to changes in the structure of a spinal midline protein that in mammals instructs the wiring that enables limb alternation, ephrin-B3. This is similar to the phenotype of ephrin-B3 and ephrin-A4 null mice which present rabbit-like hopping gait (Kullander et al. 2001, 2003; Butt et al. 2005; Akay et al. 2006; Borgius et al. 2014).
Here we found two pieces of evidence that show that even at early stages, the brachial and lumbosacral spinal cord segment circuits differ. First, we found that in the intact spinal cord both segments produce coordinated rhythmic outputs with burst of the same duration and frequency (Figs. 1, 2, 3), but when separated, and under the same conditions, the brachial spinal cord produces a slower motor rhythm, with longer bursts (Fig. 3 I). While is no possible to deduce why the brachial segments produce a slower rhythm than the lumbosacral, it does suggest that the CPGs in these two different areas of the spinal cord have differences in either the circuit wiring or expression of different receptors. In mammals, different glutamate receptors have been shown to regulate the speed of the locomotor-like activity (Talpalar and Kiehn 2010), thus it is possible that the brachial and lumbosacral spinal cord of chicks differ in their composition of glutamate receptors. Alternatively, our own results show that different concentrations of 5-HT can influence the duration and frequency of bursts (Fig. 2d, e) and therefore it is also possible that the two regions of the chick spinal cord differ in the amount and type of 5-HT receptors. Serotonin has been shown to control frequency and other aspects of locomotion in other vertebrates, including lampreys and several species of mammals (Harris-Warrick and Cohen 1985; Schmidt and Jordan 2000; Sławińska and Jordan 2019).
Second, we found that the brachial and lumbosacral rhythmic outputs are affected differentially by antagonists of inhibitory neurotransmitter, picrotoxin and strychnine (Figs. 4, 5). Both of these antagonist result in a disruption of the rhythmic activity in the lumbosacral spinal cord at lower concentrations than the brachial spinal cord (Figs. 4, 5). Further, strychnine has no effect on the rhythmic output of the brachial spinal cord, while completely supressing it in the lumbosacral (Fig. 5), again suggesting that the brachial and lumbosacral regions have different CPGs. While picrotoxin was originally considered a non-competitive GABA receptor antagonist (Robbins and Van Der Kloot 1958; Takeuchi and Takeuchi 1969), several studies have now shown that it can also inhibits glycine receptors (Wang and Slaughter 2005; Wang et al. 2006). Particularly, picrotoxin has a stronger effect as an antagonist in glycine receptors that contain a α2 subunits (Wang and Slaughter 2005; Li and Slaughter 2007). The chick lumbosacral spinal cord white matter has been shown to express glycine receptors with a α2 subunit at E13 (Harvey et al. 2000), which suggest that the effect we found of picrotoxin in the rhythmic output is, at least partially, mediated by glycine receptors. Consistent with this, the effects of picrotoxin and strychnine, a glycine receptor antagonist (Curtis et al. 1971; Young and Snyder 1973) are very similar (Figs. 4, 5). The disappearance of a rhythmic output in the lumbosacral spinal when glycine receptors are blocked (Fig. 5) shows that even when producing coordinated motor outputs between left and right, the rhythmicity of the motor output in the lumbosacral spinal is glycine dependent, while that of the brachial segments is glycine independent (Fig. 5). This would suggest that the brachial and lumbosacral spinal cord of the chick differ in their glycinergic circuity or receptors. This is not be surprising as glycine and glycinergic interneurons have been shown to be an important part of the left and right alternation circuit in several vertebrates (Droge and Tao 1993; Hagevik and McClellan 1994; Kremer and Lev-Tov 1997). This effect of glycine in the alternation of left and right is at least partially mediated by a group of commissural glycinergic interneurons which have been found in most vertebrates (Gosgnach et al. 2017). In mice, deletion of this population of neurons results in coordination between the left and right fictive locomotion and a hooping phenotype (Talpalar et al. 2013). While these glycinergic interneurons are clearly involved in the alternation of left and right motor outputs, other neuronal populations, including excitatory commissural interneurons are also involved in the alternation of left and right locomotor outputs, and therefore, left–right alteration ultimately depends on a balance of excitatory and inhibitory inputs (Restrepo et al. 2011). As mentioned above, Haimson et al., (2021) has recently show that the brachial segments of the avian spinal cord have an increased number of excitatory interneurons that project to the contralateral side. Our results suggest that the differences in the excitatory/inhibitory balance between the brachial and lumbosacral segments may not be solely due to an increase in excitatory inputs but also a reduction of inhibitory inputs, specifically glycinergic inputs.
Bilateral coordination
Despite testing several different concentrations of NMDA, 5-HT and dopamine, and combinations thereof, (Figs. 1, 2, 3), we were not able to produce an alternating rhythmic output in the lumbosacral spinal cord of the chick. It is possible that at this embryonic stage (E11–E14) the chick spinal cord is not able to produce alternating locomotor patterns. Previous studies have shown that the chick spinal cord from about E10 to E14 can produce alternation of antagonist muscles on the same side (Barry and O’Donovan 1987; Ho and O’Donovan 1993; Sholomenko and O’Donovan 1995), which we observed as well (Fig. 4a, b). In chick embryos older than E15 (but not before), stimulation of the brainstem locomotor centres can evoke locomotor activity, including both simultaneous and alternating leg movements, as well as wing flapping (Valenzuela et al. 1990), which could suggest that the alternating circuit is not completed until E15. Unfortunately, the size of the spinal cord in E15 embryos makes it impossible to survive in an in vitro preparation. In mice, the left–right alternating circuit also seems to develop later, as early mice embryos (E15) also produce only synchronous rhythmic patterns (Branchereau et al. 2000). One possibility would be that in the whole spinal cord, the brachial segments, which should produce left/right synchronous outputs are driving the lumbosacral segments. Our results show not only that the isolated lumbosacral segments produce a coordinated output in the absence of connections with the brachial segments (Figs. 3, 5), but suggest that the lumbosacral segments are driving the rhythmic brachial output. First, we found, under a variety of conditions, that the brachial output always lags behind that of the lumbosacral output (Figs. 2b, f, g, 3e, 4e–g, 5a–c). This has been reported previously in a chick in vitro preparation (Ho and O’Donovan 1993). Second, we found that after separating the brachial segments from the lumbosacral segments, the burst duration and burst frequency of the brachial segments was significantly slower than the burst frequency of the brachial segments when they were still connected to the rest of the spinal cord, while that the burst frequency of the lumbosacral segments did not change after loosing its connection to the brachial segments. (Fig. 3c–i). This shows that the burst characteristic in the whole spinal cord is closer to that of the isolated lumbosacral than the brachial segments. Finally, when either strychnine or picrotoxin was added to the whole spinal cord, the leg rhythm was disrupted first followed by the brachial segments (Figs. 4e–h, 5), even though the brachial segments were not affected by the blocking of glycine when not attached to the lumbosacral segments, suggesting that disruption of the lumbosacral rhythm eventually propagates to the brachial segments. The complete coordination between the two CPGs suggests that there are strong connections between the rhythmic centers at this embryonic stage, particularly from the lumbosacral to the brachial ones.
Alternatively, the lack in alternating movements in the lumbosacral spinal cord of chick could also reflect spinal circuits that are not only able to produce post-hatching behaviors, like walking and flying, but also behaviors present before hatching, particularly hatching behavior. To be able to exit the eggs, chicks perform a series of coordinated movements, including kicking of their legs together in a rhythmic fashion (Bekoff and Kauer 1982, 1984). Chick embryos show movement as early as E4 (Hamburger 1968; Bekoff 1976), and while this movements are highly variable and not always coordinated between the two legs or between the legs and the wings (Provine 1980; Watson and Bekoff 1990; Chambers et al. 1995), their motor patterns resemble the motor patterns of both hatching and walking (Watson and Bekoff 1990; Bekoff 1992, 1995). Therefore, is possible that the lumbosacral CPGs in chicks develop the capacity of producing both coordinated and alternating leg movements, and that in the absence of central control they default to the motor pattern that occurs earlier in development, like kicking with the legs.









留言 (0)