
記住我

The experiments described in this study were carried out at the National Microbiology Laboratory (NML) at the Public Health Agency of Canada as described in the Animal use document AUD# H-20-006. Experiments were approved by the Animal Care Committee located at the Canadian Science Center for Human and Animal Health in accordance with the guidelines provided by the Canadian Council on Animal Care. All procedures were performed under anesthesia, and all efforts were made to minimize animal suffering and to reduce the number of animals used. All infectious work with SARS-CoV-2 was performed under biosafety level 4 (BSL-4) conditions.
2.2. Cells and VirusesVero E6 (CRL-1586; American Type Culture Collection, Manassas, VA, USA) and Vero (CCL-81; American Type Culture Collection) were cultured at 37 °C + 5% CO2 in Dulbecco’s modified Eagle’s medium (HyClone, GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS), 1% L-glutamine, and 100 U mL−1 penicillin–streptomycin.
The SARS-CoV-2 strain used in these studies (SARS-CoV-2; hCoV- 19/Canada/ON-VIDO492 01/2020, GISAID accession# EPI_ISL_425177) was isolated from a clinical specimen obtained at the Sunnybrook Research Institute (SRI)/University of Toronto on VeroE6 cells and provided by the Vaccine and Infectious Disease Organization (VIDO) with permission. Infectious stocks were grown by inoculating Vero CCL-81 cells and collecting supernatant upon observation of cytopathic effect; debris were removed by centrifugation at 6000× g for 5 min and stored at −80 °C until thawed.
To generate AAV6.2FF-hACE2, the human ACE2 gene (HG10108-M, Sino Biological Inc., Beijing, China) was cloned into an AAV2 genome plasmid downstream of the composite CASI promoter [41] followed by a woodchuck hepatitis virus posttranscriptional regulatory element (WPRE), a simian virus 40 polyadenylation sequence and flanked by AAV2 inverted terminal repeats (ITRs) as described previously [42]. AAV expressing firefly luciferase (AAV-Luc) was described previously [39]. AAV vectors were produced by two plasmid transfection of human embryonic kidney 293 cells (HEK293; CRL-1573, ATCC, Manassas, VA, USA) and heparin affinity column chromatography as described previously [43]. AAV vectors were titered by TaqMan quantitative polymerase chain reaction (qPCR) analysis as described [44,45]. 2.3. Mouse ExperimentsBALB/c and C57BL/6 were purchased from Jackson Laboratories (Bar Harbor, ME, USA). Animals were housed in groups and fed standard chow diets with food and water provided ad libitum. Mice of different ages and sexes (eight to ten weeks old to over 1 year old) were administered 1 × 1011 vector genomes (vg) of AAV6.2FF-hACE2 or AAV6.2FF-Luciferase via a modified-intranasal administration [46]. Individual groups consisted of young male Balb/C mice, young female Balb/C, old male Balb/C mice, old female Balb/C mice, young male C57BL/6 mice and young female C57BL/6 mice. Each group had 36 mice equally divided between male and female with 18 mice transduced with AAV6.2FF-hACE2 and 18 mice transduced with AAV6.2FF-Luciferase. In brief, mice were anesthetized with isoflurane and the viral vector administered via the intranasal route (i.n) in a volume of 50 μL. The mouth was covered during administration to promote inhalation through the nose and subsequent distribution of the vector throughout the lower respiratory tract. Ten days after AAV transduction, mice were inoculated with 105 TCID50 of SARS-CoV-2 in a 50 μL volume given i.n. Weights were monitored on a daily basis, and animals were sacrificed at 2, 4, or 28 dpi (n = 6 mice (3 male + 3 female) per time point) and tissues were harvested. Mice were euthanized under isoflurane anesthesia after serum collection via cardiac puncture and cervical dislocation before tissue collection. 2.4. Infectious Virus QuantificationFor infectious virus assays, 50% tissue culture infective dose (TCID50) was calculated using the Reed and Muench method [47] and expressed as TCID50 per gram of sample (TCID50/g). Briefly, harvested tissue samples for infectious assays were flash frozen and stored at −80 °C. Thawed tissue samples were weighed and placed in 1 mL of minimal essential medium (MEM, HyClone, GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 1% FBS (Gibco, Life Technologies, Grand Island, NY, USA), 1× L-glutamine (Gibco, Life Technologies, Grand Island, NY, USA), and 100 U mL−1 penicillin–streptomycin (Gibco, Life Technologies, Grand Island, NY, USA) before being homogenized in a Bead Ruptor Elite Bead Mill Homogenizer (Omni International, Kennesaw, GA, USA) at 4 m/s for 30 s. Lysates were clarified by centrifugation at 1500× g for 6 min. Tissue homogenates were serially diluted in MEM supplemented with 1% heat-inactivated FBS, 1× L-glutamine, and 200 U mL−1 penicillin–streptomycin. One hundred microliter volumes of sample dilutions were added to 96-well plates of 95% confluent Vero cells containing 50 μL of the same medium in three replicates and incubated for 5 days at 37 °C with 5% CO2. Plates were monitored daily and scored for the presence of cytopathic effect on day 3–5 after infection. 2.5. Reverse Transcriptase-Quantitative Polymerase Chain ReactionTissue samples harvested for viral RNA (vRNA) detection were immersed in RNAlater (Ambion, Austin, TX, USA) 4 °C for 1 day, then stored at −80 °C until later use. Tissue samples were thawed and weighed and homogenized in 600 μL RLT buffer using a Bead Ruptor Elite Bead Mill Homogenizer (Omni International, Kennesaw, GA, USA) with a stainless steel bead at 4 m/s for 30 s. Viral RNA from 30 mg tissue samples was extracted with the RNeasy Plus Mini kit (Qiagen). For detection of SARS-CoV-2 RNA, SARS-CoV-2 E Sarbeco real-time RT–PCR assay [48], recommended by the WHO was used. The primers used were E_Sarbeco_F1 (5′-ACAGGTACGTTAATAGTTAATAGCGT-3′) and E_Sarbeco_R2 (5′-554 ATATTGCAGCAGTACGCACACA-3′). The probe was E_Sarbeco_P1 (5′-FAM555 ACACTAGCCATCCTTACTGCGCTTCG-BBQ-3′). The assay was set up using the TaqPath 1-Step Multiplex Master Mix kit (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) on a QuantStudio 5 real-time PCR system (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA), as per manufacturer’s instructions. A standard curve was generated by using plasmids coding for the SARS-CoV-2 E gene and were used on each plate for the quantification of copy numbers. 2.6. Serum ELISASARS-CoV-2 spike specific IgM and IgG antibody responses were determined using an in-house enzyme-linked immunosorbent assay (ELISA). Briefly, 96 well plates (Corning Inc, Corning, NY, USA) were pre-coated with full length spike or EBOV GP (negative control) overnight and blocked with phosphate-buffered saline (PBS) containing 5% skim milk for one hour. A serial dilution of mouse serum was carried out in triplicate in PBS containing 5% skim milk before being applied to the plate, incubated for one hour, and then washed three times in PBS plus 0.1% Tween 20 using a BioTek plate washer. Mouse IgG was detected with a Goat anti-Mouse IgG (H + L) Secondary Antibody conjugated with HRP (Catalog # 31430; 1:10,000 dilution, Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Mouse IgM was detected with a Goat anti-Mouse IgM (H + L) Secondary Antibody conjugated with HRP (Catalog # 31440; 1:10,000 dilution, Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Both IgG and IgM secondary antibodies were incubated for one hour Following a second set of washes, the plate was incubated with 3,3′,5,5′-tetramethylbenzidine (TMB) Single Solution substrate (Thermo Fisher Scientific, Waltham, MA, USA). The reaction was stopped with 1N sulfuric acid and the absorbance was read at 450 nm using a BioTek Synergy HT plate reader (BioTek, Winooski, VT, USA).
2.7. Cytokine AnalysesThe murine cytokine response was quantified by a custom ordered TaqMan™ Gene Expression Assay (FAM) (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA). The following cytokines were analysed: IL-1 alpha (Mm00439620_m1), IL-1 beta (Mm00434228_m1), IFN alpha (Mm03030145_gH), IFN beta (Mm00439552_s1), IFN gamma (Mm00434256_m1), granzyme A (Mm00439191_m1), granzyme B (Mm00442834_m1), TNF-alpha (Mm00434256_m1), TGF-beta (Mm01227699_m1), IL-2 (Mm00434256_m1), IL-4 (Mm00434256_m1), IL-6 (Mm00434256_m1), IL-8 (Mm04208136_m1), IL-10 (Mm00434256_m1), IL-12p40 (Mm01288993_m1), IL-17 (Mm00439618_m1), MCP1 (Mm99999056_m1), VEGF (Mm00437306_m1), and GAPDH (Mm99999915_g1) as a housekeeping gene control. RNA was extracted from tissues using the RNeasy plus mini kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. For removal of genomic DNA and reverse transcription, SuperScript IV VILO Master Mix with ezDNase Enzyme (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) was used as per manufacturers protocols. TaqMan®Fast Universal PCR MasterMix (2×) (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) was used according to the manufacturer’s instructions in a two-step qRT-PCR reaction in triplicate. The PCR was performed on a Quantstudio 5 (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA). One microliter (10 ng) of cDNA sample was assayed per reaction. Each reaction consisted of 1 cycle of 95 °C for 20 s, followed by 50 cycles of 95 °C for 3 s and 60 °C for 30 s. The ΔΔCt method was used with normalization within a sample on GAPDH (ΔCt) calculated for each gene. The comparison was against hACE2-transduced mice vs. Luciferase transduced mice (ΔΔCt) or Luciferase transduced mice vs. mock-uninfected mice (ΔΔCt).
2.8. Western BlotHEK 293 cells were transfected with pACASI-hACE2-WPRE or pACASI-Luc-WPRE using PEI MAX™ (Polysciences, Inc., Warrington, PA, USA) as per manufacturer’s instructions. 48 h post-transfection, cells were washed twice with cold PBS, collected and pelleted at 450 × g 5 min. The cell pellet was lysed in 500 μL cold RIPA buffer (50 mM Tris HCl, pH 7.5; 150 mM NaCl; 1% Triton X-100; 0.1% SDS; 10 mM EDTA; 1% sodium deoxycholate) supplemented with protease inhibitor cocktail (Roche Applied Science, Penzberg, Germany) and incubated on ice for 30 min. Samples were centrifuged at 18,000× g for 15 min at 4 °C to remove cell debris. Twenty microlitres of cell lysate was separated on a 12% SDS PAGE gel, transferred to polyvinylidene fluoride (PVDF) membrane and blocked overnight at 4 °C in blocking solution (5% skimmed milk and 0.1% Tween 20 in PBS). The membrane was incubated 1 h at RT with a goat anti-ACE2 antibody (Cat no. AF933 R&D Systems, Minneapolis, MN, USA) diluted 1:500 in blocking solution. Membranes were washed five times in washing buffer (0.1% Tween 20-PBS) for five minutes and then incubated for 1 h at RT with an HRP conjugated donkey anti-goat IgG secondary antibody (Cat. No. PA1-28664, Thermo Fisher Scientific, Waltham, MA, USA) diluted 1:5000 in blocking solution. The membrane was washed five times with washing buffer and the chemiluminescent signal was detected using SuperSignal West Pico PLUS chemiluminescent substrate (Thermo Fisher Scientific, Waltham, MA, USA) and imaged with a ChemiDocTM Molecular Imager (Bio-Rad, Hercules, CA, USA). Molecular Weight (MW): GeneDireX BLUelf Prestained Protein Ladder Cat. PM008-0500 (GeneDireX Inc, Taiwan, China).
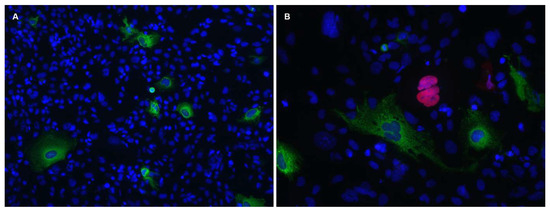
2.9. Immunofluorescence Analysis of hACE2 Expression in Murine LungsGroups of four mice transduced with 1 × 1011 vg of AAV6.2FF-hACE2 or AAV6.2FF-Luciferase intranasally were euthanized 10 days post-vector administration and exsanguinated via heart puncture. Lungs were perfused via the heart with PBS and then instilled with optimal cutting temperature compound (OCT)/PBS (1:1) via the trachea. Lungs were removed en bloc and placed directly into an OCT containing cryomold and placed on dry ice to freeze. Solid blocks of lung tissue were stored at −80 °C. 8 μm cryosections were placed on Superfrost Plus microscope slides (Thermo Fisher Scientific, Waltham, MA, USA) and allowed to dry at RT for 3 h. Slides were fixed in ice-cold acetone for 15 min and then washed twice in 0.1% Tween 20-PBS for 5 min each followed by a 5 min wash in PBS. Antigen retrieval was performed by heating antigen retrieval buffer (10 mM sodium citrate, 0.05% Tween-20, pH 6.0) to 100 °C in a microwave and placing slides in heated solution for 10 min for a total of three times. Slides were then washed in PBS for 5 min and a hydrophobic circle was drawn around the lung tissue using a Dako pen. Lung tissue was permeabilized in 0.1% Triton X-100-PBS for 10 min at RT and blocked with 10% FBS-PBS for 1 h at RT. Rabbit anti-ACE2 mAb (Cat no. ab108209 Abcam Inc., Toronto, ON, Canada) diluted 1:100 in the blocking buffer was added to slides and incubated at 4 °C for two days. Slides were washed three times with PBS for 10 min each before being incubated with goat anti-rabbit Alexa 488 (Cat. no. A-11034, Life technologies, Carlsbad, CA, USA) diluted 1:500 in blocking buffer for 1 h at RT. Slides were washed three times with PBS for 10 min each and mounted using ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific, Waltham, MA, USA). Slides were imaged using an Axio Observer inverted fluorescence microscope (Carl Zeiss MicroImaging GmbH, Göttingen, Germany).
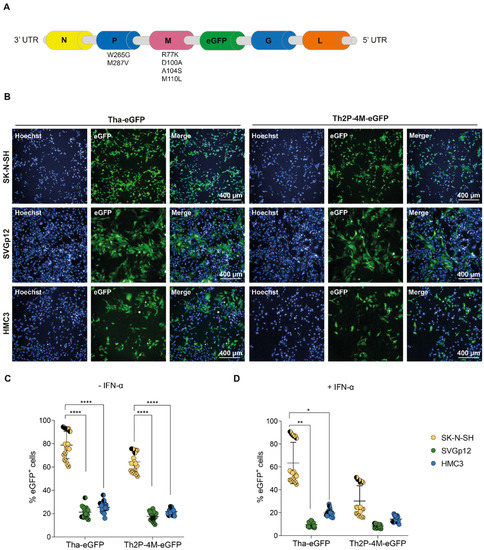
2.10. In Vivo Luciferase ImagingMice that were administered a total of 1 × 1011 vector genomes of AAV6.2FF-Luc or AAV6.2FF-hACE2 intranasally (Section 2.3) were used for detection of luciferase expression. Bioluminescence imaging was performed on days 3, 5, 7, and 10 after intraperitoneal (IP) administration of XenoLight RediJect D-Luciferin (Perkin Elmer, Waltham, MA, USA) at a concentration of 150 mg per kg using the IVIS SpectrumCT instrument (Perkin Elmer, Waltham, MA, USA). Resultant data were analysed and the signal intensity quantified using Living Image software (Perkin Elmer, Waltham, MA, USA). 2.11. Histology and ImmunohistochemistryTissues were fixed in 10% neutral phosphate buffered formalin, routinely processed, sectioned at 5 μm and stained with hematoxylin and eosin (HE) for histopathologic examination. Paraffin tissue sections were quenched for 10 min in aqueous 3% hydrogen peroxide. Epitope retrieval was performed using an in-house glycan retrieval solution in a Biocare Medical Decloaking Chamber (Biocare Medical, Pacheco, CA, USA). The primary antibody applied to the sections was SARS-CoV-2 (2019-nCoV) Nucleocapsid, Rabbit MAb (#40143-R019, Sino Biological Inc., Beijing, China) used at a 1:6000 dilution for thirty minutes. They were then visualized using a horse radish peroxidase labelled polymer, Envision® + system (anti-rabbit) (Dako, Santa Clara, CA, USA) and reacted with the chromogen diaminobenzidine (DAB). The sections were then counter stained with Gill’s hematoxylin. Semi-quantitative lesion scoring was performed as follows: The percentage affected of each section examined was scored as 0 = no pathological changes, 1 = ≤25% of lung section affected, 2 = >25% and ≤50% of lung section affected, 3 = >50% and ≤75% of lung section affected and 4 = >75% of lung section affected. Additionally, each section was assigned a severity score or 0 = no lesions, 1 = mild lesions, 2 = moderate lesions and 3 = severe lesions Additionally, a score of 0 for not present and 1 for present was given for each of the following parameters and an average score out of 8 was assigned for histological features present: bronchiolitis (including inter-epithelial inflammatory cells, necrosis of bronchiolar epithelium and debris in lumen), diffuse alveolar damage (including necrosis of alveolar epithelial cells, cellular debris in alveoli and intra-alveolar fibrin), alveolar edema, alveolar hemorrhage, hyperplasia of type II pneumocytes, perivasculitis and vasculitis and presence of multinucleated cells. In total, a score was assigned for each section which included percentage affected (/4), severity (/3) and histological features (/8) for a total score /15. For most groups, in which 4 sections were examined the scores from all the sections in each group were added together and are presented in Table S1. In one group in which 8 sections were examined the total score was divided by 2. 2.12. Data AnalysisResults were analysed and graphed using Prism 8.2.1 software (Graphpad Software, La Jolla, CA, USA). The statistical significance between the groups was determined using a Mann–Whitney test or one-way and two-way analysis of variance (ANOVA).
4. DiscussionIn this study, we have developed and characterized a mouse model of SARS-CoV-2 infection using AAV6.2FF-hACE2 mediated gene delivery to the mouse respiratory system and characterized the infection in BALB/c and C57BL/6J mice of different ages and sex. As expected, we confirmed that that AAV6.2FF mediated gene delivery was able to transduce gene expression in the lungs and nasal turbinates of mice, as previously demonstrated [37]. In both mouse species, early in the infection, mice transduced with AAV-hACE2 and infected with SARS-CoV-2 have high viral titres in respiratory tissues not seen in control AAV-Luc-transduced mice. AAV6.2FF mediated gene delivery of hACE2 was able to successfully generate a mouse model for SARS-CoV-2 infection that shows sites of infection that are comparable to those that are infected in species that are naturally susceptible to infection such as Syrian hamsters and deer mice [7,29]. This outcome is similar to the results of others that have similarly demonstrated AAV mediated transduction of the mouse airway with hACE2 using AAV9 and AAV2/8 [34,38,49]. Mice transduced with AAV-hACE2 produce both a cytokine and chemokine response in the lungs and nasal turbinates, whereas mice transduced with AAV-Luc did not produce a strong inflammatory response as a consequence of SARS-CoV-2 infection. Our results show that an IgG and IgM antibody response was generated in AAV-hACE2 mice after infection, while only a weak response was detected in AAV-Luc transduced mice. Thus, this animal would be a valuable model to improve our understanding of SARS-CoV-2 infection using a readily accessible small animal model. In addition, this model will allow for testing of vaccine and therapeutic candidates, as has been done by our group [42].Although there are a few limitations of the AAV6.2FF system including large multiplicity of infection with AAV-hACE2, and potentially the cloning capacity of the AAV vector, development of AAV vectors expressing the hACE2 was efficient and rapid, resulting in the generation of a reproducible murine model for SARS-CoV-2 within three weeks. This short time course compares favorably with the much longer time required to develop transgenic mice expressing hACE2. Further, hACE2 expression after AAV-hACE2 intranasal inoculation is restricted to the respiratory system whereas transgene expression may be widespread [26,50,51] and result in infection in organs that are not usually associated with SARS-CoV-2 infection such as direct infection in brains of K18 hACE2 transgenic mice, which is a major factor contributing to the fatal outcome. Although AAV6.2FF-Luc transduced mice also had lesions and inflammation upon SARS-CoV-2 infection in the absence of SARS-CoV-2 antigen detection, inflammation associated with DMEM intranasal instillation of liquid into the lungs has been shown to cause instillation-associated histopathology [52]. Advantages that AAV vectors have over adenovirus-vectors (Ad) is that AAV is not strongly immunogenic, which was confirmed during the cytokine analysis in the mice given AAV-Luc, AAV-based transduction results in long lasting gene expression, and the tissue tropism of AAV can also be tailored using different serotypes.We observed that the virus titers in the nasal turbinates and lungs, as well as manifestations of infection, were somewhat variable across the different groups of AAV-hACE2-transduced mice. One interesting group of mice were the older male BALB/c mice. These mice exhibited more apparent clinical signs such as rapid breathing and ruffled fur and considerably greater weight loss compared to other groups. Cytokine and chemokine responses were not dramatically different from the other groups of mice; however, the old male BALB/c transduced with AAV6.2FF-Luc did show a cytokine and chemokine response in 14/18 genes tested which suggests that these mice have some age-related innate immune system differences compared to the other groups of mice. Further to differences related to age, IHC results indicated that AAV6.2FF-hACE2-transduced old male mice did not have detectable viral protein antigens in contrast to the old female animals that had scores similar to the other mouse groups. In considering an explanation for the lack of antigen detection in older male mice by IHC despite detection of significant viral replication, we suggest that there may be differences in the distribution of replicating virus in older male animals compared to other groups. While the same lung lobes were consistently collected from each animal in each group for evaluation of virus titer or pathology, measurement of virus titers was done on a different lobe than that used for pathology. The differences in the older male animals might suggest differences in the sites of preferential virus replication in aged male mice compared to the other groups, and that virus replication was considerably limited in the lung lobe used for pathology. Further evaluation of the tissue specific distribution of viral replication in each group would be needed to confirm this possibility. However, the results do suggest possible aged and sex-related differences in outcome. Similar to infected patients, young AAV6.2FF-hACE2–transduced mice with normal immune systems developed mild disease whereas older male mice, like old male patients [53], were more profoundly affected, as weight loss and rapid breathing were observed in these mice. In COVID-19 patients with acute respiratory illness, the main clinical manifestation is severe lung inflammation. Future studies using other strains of mice may be needed to address the susceptibility of old male mice to infection with SARS-CoV-2 using this model.Several infectious models have been characterized such as Syrian hamsters [7,8,9], ferrets [11,12,13,14], deer mice [10], and non-human primates [15,16,17,18,19]. These models are valuable tools for the study of SARS-CoV-2 infection and COVID-19 disease; however, the mouse’s small size, small housing requirements, and availability make the mouse model ideal for large scale studies without the lethal infection of the central nervous system (CNS) that is seen with the hACE2-transgenic mouse models. Continued development of this mouse model for SARS-CoV-2 will contribute to the development of vaccines, therapeutic agents and other countermeasures as a means to further enhance protection of the human population against infections by this pathogen.
留言 (0)