1. IntroductionRodents (Rodentia) are a large order of mammals that are important hosts for a number of zoonotic viral agents, including mammarenaviruses. According to the International Committee on the Taxonomy of viruses (ICTV) taxonomy, the genus Mammarenavirus, family Arenaviridae, comprises 39 species of 46 viruses. These species can be further grouped into the New World and Old World complexes based on antigenic and phylogenetic similarities and geographic distribution [
1]. Rodents are often persistently infected with mammarenaviruses, and human exposures occur during contact with contaminated rodent excreta (urine, fecal material, and saliva) that arise opportunistically due to the peri-domestic habits of many of these hosts [
2,
3]. Lassa fever exerts a significant zoonotic disease burden in Western African countries in which it is endemic due to the host range of its reservoir Mastomys sp. [
4]. Many Lassa virus (LASV) infections are mild or even asymptomatic, though severe clinical cases have an observed fatality rate of 15% [
5]. Another emergent arenavirus, Lujo virus (LUJV), exemplifies the need for active viral surveillance to discover and describe this diversity. LUJV was responsible for a small outbreak cluster with extremely high fatalities in 2008, when a critically ill person was airlifted from Zambia to a hospital in South Africa [
6]. Both the index case and three persons in direct contact with the patient died of hemorrhagic fever, with a fifth person recovering. Since the first outbreak, no additional cases of LUJV have been reported, and the virus’s host origins remain unknown.Biosurveillance studies in several African rodent species have identified a large diversity of mammarenaviruses (see [
7]), such as Mobala (MOBV), Ippy, Mopeia (MOPV), Gairo (GAIV), Luna (LUNV), Lunk and Mariental viruses [
8,
9,
10,
11,
12,
13,
14,
15,
16]. These Old-World arenaviruses have mostly originated from genera within the Muridae family (e.g., Mastomys, Praomys, Mus, Grammomys, and Micaelamys) from a wide geographical distribution (West, Central, East, and Southern Africa). Moreover, since mammarenaviruses are also generally associated with specific rodent species, with speciation possibly affected by host-switching, recombination, and reassortment [
7], investigating related species to identify the diversity of potentially zoonotic agents among non-volant small mammals is of increasing interest. The zoonotic potential of many mammarenaviruses has also not been determined, though serological findings suggest that exposures to widespread viruses such as MOPV from Southern Africa have occurred [
8].Current species demarcation criteria by the International committee on the taxonomy of viruses (ICTV) for arenaviruses include both genetic and phenotypic features [
17]. Viral species are differentiated if there is less than 80% and 76% nucleotide sequence identity within the S and L genomic segments, respectively. An amino acid divergence greater than 12% within the nucleoprotein gene represents another criterion to delimit separate species. Additional features include the association with distinct hosts (or sympatric hosts), distinct geographical distribution, and the ability to cause human disease.
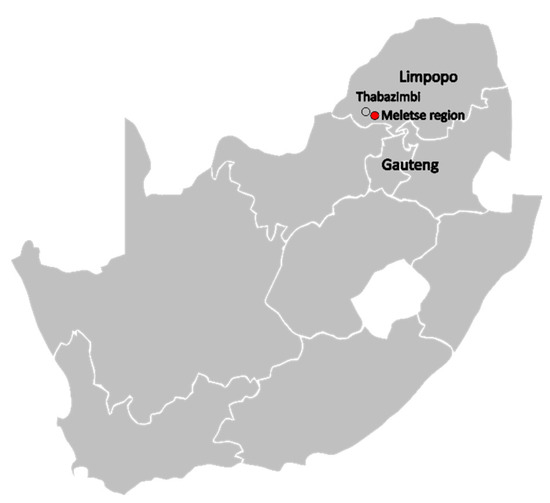
Here, we report the findings of mammarenavirus seasonal host surveillance carried out over two years among non-volant small mammals (rodents, sengis, and shrews) in the Limpopo province of South Africa. The finding suggests widespread distribution of mammarenaviruses closely associated with Mariental and Lunk mammarenaviruses in Southern Africa.
4. DiscussionUrban expansion, habitat loss, and climate-driven landscape changes are major drivers of the increase in opportunities for novel zoonotic pathogens to emerge [
38,
39]. Arenaviruses with both known and unknown zoonotic potential have been reported from African small mammal species. Improving the characterization of such viruses and our ecological understanding of this viral diversity would strengthen our preparedness for potential future viral outbreaks and aid in developing tools to help investigate other related highly pathogenic viruses [
3,
40].We identified two arenaviruses from two different rodent species during two years of sampling from the Meletse region in Limpopo. The percentage positivity for arenaviruses was low overall (Mus (Nannomys) and Micaelamys genera. Though Mastomys spp. are known hosts of MOPV, LASV, GAIV, LUNV, and Merino-Walk viruses, none of the 73 fecal or 57 kidney samples (representing 23% of all samples tested) were shown positive for arenavirus RNA. This is in contrast with previous reports, which found Mastomys as the most regular host for Old World arenaviruses [
8,
41]. Surveillance of this species at the site will be continued as the sample sizes of Mastomys sp. collected may have been too few per season to identify low prevalence mammarenaviruses present in the populations.UP12291/M. namaquensis/LP/RSA/2017 and UP12472/M. minutoides/LP/RSA/2017 genomes share the closest genetic similarity to Mariental and Lunk viruses, respectively, which were initially reported from other southern African countries [
10,
11]. These thus represent the first descriptions of these arenaviruses from rodent species in South Africa. Mariental virus was initially described from the lung tissues of a Micaelamys namaquensis rodents captured in 2012 in the city of Mariental in Namibia. UP12291/M. namaquensis/LP/RSA/2017 was identified from the kidney, lung, liver, spleen, urine and fecal of the same host species, though separated geographically by 977 km (Mariental town in Namibia to the Meletse region in South Africa). Detection of viral RNA from both urine and fecal material is suggestive of active excretion of viral particles into the environment. According to the International Union for Conservation of Nature (IUCN) red list, the distribution of Micaelamys namaquensis, is throughout Southern Africa (including southern Angola, Western Mozambique, Botswana, Zimbabwe, Namibia, Eswatini and South Africa [
42]). Other arenaviruses reported from the host, include Bitu, Okahandja, and Witsand viruses from Angola, Namibia, and South Africa [
8,
9,
11], though group in a basal clade to Mariental and UP12291/M. namaquensis/LP/RSA/2017 (
Figure 3 and
Figures S1–S3). The former viruses and the Mariental lineage are rather divergent and share only 67–71.8% amino acid similarity among the GP protein and 58–63% among NP proteins.Lunk virus was described from the kidney of Mus minutoides captured in 2010 in Kasama, Zambia. RNA of UP12472/M. minutoides/LP/RSA/2017 was detected from the kidney, liver, lung, and spleen of the same host species, though capture sites are separated by 2622 km (Kasama to the Meletse region). Mus minutoides has been recorded by the IUCN red list to be extant within Zambia, Zimbabwe, Eswatini, Western Malawi and Mozambique and along coastal regions of Southern Namibia and South Africa [
43]. Lunk and UP12472/M. minutoides/LP/RSA/2017 are grouped in a phylogenetic clade (
Figure 3 and
Figures S1–S3) comprising another Mus-hosted arenavirus, Rat arenavirus 1-YN2013, and well as Dandenong virus from humans and LCMV that is associated with both, and share 78–81% and 77–78% amino acid similarity among NP and GP proteins, respectively. The findings would suggest diverse arenaviruses present among the Mus host genera.The arenaviruses described here meet many, though not all, of the ICTV’s species demarcation criteria to be considered members of either the Mariental or Lunk mammarenavirus species. Both viruses from UP12291/M. namaquensis/LP/RSA/2017 and UP12472/M. minutoides/LP/RSA/2017 originated from the same host species as Mariental and Lunk viruses, respectively, and in similar geographical regions (South Africa, Namibia and Zambia) as these hosts are broadly distributed throughout southern Africa [
11]. The nucleoprotein amino acid similarities are above the minimum required 88% sequence identity shared by species, though in the case of UP12291/M. namaquensis/LP/RSA/2017 and Mariental virus, only 88.8% [
44]. However, the nucleotide sequence identities of the S and L segments are below the threshold of 80% and 76%, respectively, for either UP12291/M. namaquensis/LP/RSA/2017 and Mariental virus as well as UP12472/M. minutoides/LP/RSA/2017 and Lunk virus. Thus, whether these newly described arenaviruses represent strains of existing species or new species will be based on the discretion of the ICTV’s Arenaviridae study group.The zoonotic risk of many of the diverse Old World mammarenaviruses described from several African countries is undetermined. Moreover, limited research has been performed on recently identified arenaviruses, particularly regarding their pathogenicity and capability of infecting people or other animals. Known arenavirus receptors include alpha-dystroglycan and transferrin receptor 1 [
35], the former used by known pathogenic Old World mammarenaviruses like LASV, MOBV, LUNV, MOPV, Morogoro, and Dandenong viruses as well as LCMV [
36]. Alpha-dystroglycan is a conserved and ubiquitous cell surface protein assisting in the adherence to the extracellular matrix, that may be utilized by arenaviruses to avoid the early endocytic pathway [
32]. Specifically, arenaviruses interact with the matrigylcan carbohydrate on alpha-dystroglycan, though the exact molecular interactions for recognition have recently been better described [
36]. The GP1 subunit terminal residues are conserved as they serve as a recognition motif in cleaving the glycoprotein precursor into GP1 and GP2 subunits and have been recognized as involved in stabilizing the spike glycoprotein to form the matriglycan binding site [
32,
36]. The conserved residue is ‘RRLL’ among most Old World alpha-dystroglycan tropic mammarenaviruses, with variations including ‘RRLM ‘and ‘RRLA’ [
36] (
Figure S9). Inference of GP1 subunit similarities and the RRLL recognition motif may suggest that UP12472/M. minutoides/LP/RSA/2017 (and Lunk virus) could be capable of utilizing the alpha-dystroglycan receptor (similarly to LCMV and Dandenong within the same phylogenetic clade). The consequences of a Leu261 replacement with Ile261 on the structural conformation of a stable matriglycan binding site for either Mariental virus or UP12291/M. namaquensis/LP/RSA/2017 is unknown and a unique variation of the RRLL recognition motif (
Figure S9). Whether the viruses described here can utilize the alpha-dystroglycan receptor and potentially infect human cells can be practically confirmed with functional studies assessing zoonotic potential [
8]. The scope of clinical disease associated with known pathogenic mammarenavirus infection is worldwide, with a spectrum of disease severity ranging from asymptomatic to fatal hemorrhagic fever. Improved serosurveillance studies in communities at risk of contact with rodent species that are known hosts for Old World arenaviruses could provide more insights into possible exposure events.


留言 (0)