

記住我
Since it has been declared as a global pandemic by the World Health Organisation (WHO), the coronavirus disease (COVID-19) has continued its devastating impact on public health. Remdesivir (Veklury, Gilead), a broad-spectrum adenosine analog prodrug initially developed for the treatment of the Ebola virus infections, has been the first drug to recieve FDA approval for use in COVID-19 patients. Despite the limited benefits in hospitalized patients (Consortium, 2020; Goldman et al., 2020; Ader et al., 2022), early administration of remdesivir in non-hospitalized COVID-19 patients reduced the risk of hospitalization or death by 87% compared to placebo (Gottlieb et al., 2021). By the end of 2021, two oral antiviral drugs i.e., Molnupiravir (Lagevrio, Merck) and Nirmatrelvir (in combination with ritonavir, Paxlovid, Pfizer) have been authorized by FDA and other countries/regions for emergency use in COVID-19 patients at high risk of hospitalization (Hammond et al., 2022; Syed, 2022).
The parent nucleoside of remdesivir, GS-441524, has been found to be the major circulating metabolite after administration of remdesivir (Warren et al., 2016; Williamson et al., 2020; Yan and Muller, 2020). GS-441524 exerts antiviral activity i) in cats infected with the feline coronavirus and ii) against SARS-CoV-2 infection in AAV-hACE2-transduced mice (Pedersen et al., 2019; Li et al., 2021). It was well-tolerated in cats at doses exceeding that of remdesivir in humans, and in one human subject (Pedersen et al., 2019; V. Yan, 2021). GS-441524 has also been shown to have a favorable oral bioavailability in mice compared to remdesivir (Xie and Wang, 2021).
Molnupiravir (EIDD-2801, MK-4482) is the orally bioavailable counterpart of the ribonucleoside analogue N4-hydroxycytidine (NHC, EIDD-1931), which was initially developed for the treatment of infections with the influenza virus (Toots et al., 2019). NHC exerts in vitro antiviral activity against multiple RNA viruses of different families by incorporation into viral RNA, resulting in the accumulation of deleterious mutations in the nascent viral RNA, and consequently, error catastrophe of the viral genome (Urakova et al., 2017). Molnupiravir exerts antiviral activity in SARS-CoV-2 infected mice, Syrian hamsters and ferrets (Cox et al., 2020; Wahl et al., 2020; Rosenke et al., 2021), including against several VoCs (Abdelnabi et al., 2021d). Recent data analysis from MOVe-OUT trial (NCT04575597) with molnupiravir showed a relative risk reduction of hospitalization and respiratory interventions in molnpiravir-treated patients of ∼34% (Johnson et al., 2022).
Combinations of antiviral drugs, such as against SARS-CoV-2, may allow to achieve a more potent effect than with monotherapy. This may allow for dose-reductions and result as well as in a reduced risk of resistance development. We previously reported that favipiravir significantly potentiates the antiviral efficacy of molnupiravir when co-adminstered to SARS-CoV-2 infected Syrian hamsters (Abdelnabi et al., 2021a). Similarly, the combined treatment of SARS-CoV-2-infected Syrian hamsters with favipiravir and GS-441524 has been reported to be more efficient as compared to monotherapy with either drug (Chiba et al., 2022). Recently, it has also been reported that combination of suboptimal concentrations of molnupiravir and remdesivir compeletely abrogated the production of SARS-CoV-2 infectious virus particles in the apical wash of human nasal epithelium cultures, although no significant, antiviral effect was detected at the RNA level (Jonsdottir et al., 2022).
Since we previously demonstrated that the combination of molnupiravir and favipiravir results in SARS-CoV-2 infected hamsters in an unexpected potent antiviral effect, we here wanted to assess the combined antiviral potency of the parent nucleoside of remdesivir (GS-441524) with molnupiravir against SARS-CoV-2 infection (in vitro and in the hamster infection model).
Material and methodsSARS-CoV-2SARS-CoV-2 Beta B.1.351 variant (derived from hCoV-19/Belgium/rega-1920/2021; EPI_ISL_896474, 2021-01-11) (Abdelnabi et al., 2021c) and SARS-CoV-2 omicron BA.5 variant [BA.5.2.1, EPI_ISL_14782497] were recovered from nasopharyngeal swabs taken from RT-qPCR confirmed patients. Infectious viruses were first isolated by passaging on Vero E6 cells then passage two virus stocks of the beta variant and the BA.5 variant on Vero E6 and Calu-3 cells, respectively, were used for the studies described here. Live virus-related work was conducted in the high-containment A3 and BSL3+ facilities of the KU Leuven Rega Institute (3CAPS) under licenses AMV 30112018 SBB 219 2018 0892 and AMV 23102017 SBB 219 20170589 according to institutional guidelines.
CellsVero E6 cells (African green monkey kidney, ATCC CRL-1586) were cultured in minimal essential medium (MEM, Gibco) supplemented with 10% fetal bovine serum (Integro), 1% non-essential amino acids (NEAA, Gibco), 1% L-glutamine (Gibco) and 1% bicarbonate (Gibco). A549-Dual™ hACE2-TMPRSS2 cells obtained by Invitrogen (Cat. a549 days-cov2r) were cultured in DMEM 10% FCS (Hyclone) supplemented with 10 μg/ml blasticidin (Invivigen, ant-bl-05), 100 μg/ml hygromycin (Invivogen, ant-hg-1), 0.5 μg/ml puromycin (Invivogen, ant-pr-1) and 100 μg/ml zeocin (Invivogen, ant-zn-05). End-point titrations and antiviral assays were performed with media containing 2% fetal bovine serum instead of 10% and no antibiotics. Both cells were kept in a humidified 5% CO2 incubator at 37°C.
CompoundsMolnupiravir (EIDD-2801) and GS-441524 were purchased from Excenen Pharmatech Co., Ltd. (China). For in vitro assays, 10 mM stocks of compounds were made by dissolving in DMSO. For in vivo studies, molnupiravir was formulated as 50 mg/ml stock in a vehicle containing 10% PEG400 (Sigma) and 2.5% Kolliphor-EL (Sigma) in water. GS-441524 was formulated as a 15 mg/ml stock in 30% PEG-400 (Sigma) in PBS containing 1%DMSO.
In vitro evaluation of combined antiviral activityCombination study was performed by treating A549-Dual™ hACE2-TMPRSS2 cells (15,000 cells/well) with a matrix of 2-fold dilution series of EIDD-1931 and GS-441524 then the cells were infected with the BA.5 variant at MOI of 0.001. After 4 days of incubation, cell viability was quantified by the colorimetric MTS/PMS method (by measuring absorbance at 490 nm) and the obtained absorbance values were used for calculation of %inhibition of virus induced CPE. Data were then analyzed with the SynergyFinder webtool (Ianevski et al., 2017) based on zero interaction potency (ZIP) model. The 50% effective concentration (EC50) of each compound, which is defined as the concentration of compound that is required to inhibit virus-induced cell death (or CPE) by 50%, was determined using logarithmic interpolation.
SARS-CoV-2 infection model in hamstersThe hamster infection model of SARS-CoV-2 has been described before (Abdelnabi et al., 2022). Female Syrian hamsters (Mesocricetus auratus, Janvier Laboratories) were housed as pairs in individually ventilated isolator cages (IsoCage N Bio-containment System, Tecniplast) at 21°C, 55% humidity and 12:12 days/night cycles. Housing conditions and experimental procedures were approved by the ethics committee of animal experimentation of KU Leuven (license P065-2020). For infection, female hamsters of 6–8 weeks old were first anesthetized with ketamine/xylazine/atropine and inoculated intranasally with 50 µL containing 1 × 104 TCID50 SARS-CoV-2 beta variant (day 0). After 4 days of infection, animals were euthanized for collection of the lungs and further analysis by intraperitoneal (i.p.) injection of 500 μL Dolethal (200 mg/ml sodium pentobarbital).
In vivo treatment regimenThe doses of compounds were selected such that they result as monotherapy in a suboptimal/moderate antiviral activity (i.e., ≤ 2log10 reduction in infectious virus titers). The selection of the suboptimal Molnupiravir dose was based on a dose-response study against the WT strain that we have previously published in (Abdelnabi et al., 2021b). For GS-441524, we have tested both 25 and 50 mg/kg (BID) in our model. The 25 mg/kg BID dose did not result in any reduction in infectious viral loads while the 50 mg/kg BID dose resulted in limited antiviral efficacy (unpublished data). Therefore, the GS-441524 (50 mg/kg, BID) dose was selected for the combination study. Hamsters were treated twice daily (BID) via oral gavage with either vehicle, 150 mg/kg EIDD-2801, 50 mg/kg GS-441524 or combination of both compounds starting from the time of infection (d0) with SARS-CoV-2 beta variant. The BID treatments were done 8 h apart and the compounds were given sequentially for the combination therapy. All the treatments continued until day 3 post-infection (pi). Hamsters were monitored for appearance, behavior and weight. At day 4 pos-infection (pi), hamsters were euthanized as mentioned earlier. Lungs were collected for viral loads quantification and histopathology scoring. Viral RNA and infectious virus were quantified by RT-qPCR and end-point virus titration, respectively.
SARS-CoV-2 RT-qPCRHamster lung tissues were homogenized using bead disruption (Precellys) in 350 µL TRK lysis buffer (E.Z.N.A.® Total RNA Kit, Omega Bio-tek) followed by centrifugation (10.000 rpm, 5 min) to pellet the cell debris. RNA was then extracted according to the manufacturer’s instructions. RT-qPCR was performed on a LightCycler96 platform (Roche) using the iTaq universal Probes One-Step RT-qPCR kit (BioRad) with N2 primers and probes (IDT) as described before (Boudewijns et al., 2020). Standards of SARS-CoV-2 cDNA (IDT) were used to express viral genome copies per mg tissue (Kaptein et al., 2020).
End-point virus titrationsLung tissues were homogenized using bead disruption (Precellys) in 350 µL MEM medium and centrifuged (10,000 rpm, 5min, 4°C) to pellet the cell debris. To quantify infectious SARS-CoV-2 particles, endpoint titrations were performed on confluent Vero E6 cells in 96- well plates. Viral titers were calculated by the Reed and Muench method (Reed and Muench, 1938) using the Lindenbach calculator and were expressed as 50% tissue culture infectious dose (TCID50) per mg tissue.
HistologyFor histological examination, the lungs were fixed overnight in 4% formaldehyde and embedded in paraffin. Tissue sections (5 μm) were analyzed after staining with hematoxylin and eosin and scored blindly for lung damage by an expert pathologist. The scored parameters, to which a cumulative score of 1–3 was attributed, were the following: congestion, intra-alveolar hemorrhagic, apoptotic bodies in bronchus wall, necrotizing bronchiolitis, perivascular edema, bronchopneumonia, perivascular inflammation, peribronchial inflammation and vasculitis.
StatisticsThe detailed statistical comparisons, the number of animals and independent experiments that were performed is indicated in the legends to figures. “Independednt experiments” means that experiments were repeated separately on different days. The analysis of histopathology was done blindly. All statistical analyses were performed using GraphPad Prism nine software (GraphPad, San Diego, CA, United States). Statistical significance was determined using the non-parametric Mann Whitney U-test. p-values of <0.05 were considered significant.
Sample size justificationFor the tested antiviral compounds, we need to achieve at least 1 log10 reduction in viral RNA levels in treated subjects compared to the untreated, infected control group. Group size was calculated on the independent t-test with an effect size of 2.0 and a power of 80% (effect size = deltamean/SD = 1 log10 decrease in viral RNA/0.5 log10), resulting in 5-6 animals/group. Sample sizes maximized considering limits in BSL3 housing capacity, numbers of animals that can be handled under BSL3 conditions, and availability of compounds.
ResultsFirst, the dose-response effects of the active metabolite of molnupiravir (EIDD-1931) and GS-441524 were determined against the BA.5 variant in A549-Dual™ hACE2-TMPRSS2 cells using a cytopathic effect (CPE) reduction asssay. Both EIDD-1931 (Figure 1A) and GS-441524 (Figure 1B) efficiently inhibited the BA.5 variant replication with EC50 values of 0.96 ± 0.13 and 2.1 ± 0.27 µM, respectively.
FIGURE 1. In vitro efficacy of the combination of EIDD-1931 and GS-441524 against SARS-CoV-2 BA.5 variant. Dose-response effect of (A) EIDD-1931 and (B) GS-441524 on SAR-CoV-2 (BA.5 variant)-induced cytopathic effect (CPE) at MOI 0.001, as quantified in A549-Dual™ hACE2-TMPRSS2 cells by the MTS/PMS method. Data represented in tehj graphs are mean values ± standard deviations (SD) from two independent experiments. (C) Dose-response matrix for EIDD-1931 and GS-441524 representing %inhibition of virus-induced CPE in BA.5-infected A549-Dual™ hACE2-TMPRSS2 cells. (D) Heat map of the delta scores (%) for the combined EIDD-1931 and GS-441524 antiviral effects based on ZIP analysis where δ = 0, δ > 0, and δ < 0 correspond to zero interaction, synergy, and antagonism, respectively. The overall ZIP score represents the response beyond expectation (in %). In the range −10 < ZIP <10, the compounds are likely to act in an additive manner, Score ≥10 indicate syenrgism. MSA = most synergistic area. The graphics for combination represent the means of two independent experiments.
Next, the combined antiviral activity of EIDD-1931 and GS-441524 against the BA.5 variant was assessed in a checkerboard (matrix) format with seven serial dilutions of each compound. The inhibition of virus-induced CPE by each compound alone or in combination was determined (Figure 1C), and the data were analyzed with the SynergyFinder webtool (Ianevski et al., 2017) based on zero interaction potency (ZIP) model (Figure 1D). The EIDD-1931 and GS-441524 combination resulted in an overall synergy score of 2.28 (Figure 1D). A large surface of the combined antiviral effect of both compound resulted in additive antiviral effects (Figure 1D). However, at certain concentration ranges, the combinations resulted in a marked synergistic effect with a score of 21.32 in the most synergistic area (Figure 1D). No cytotoxicity was observed for at any combined concentrations of both compounds.
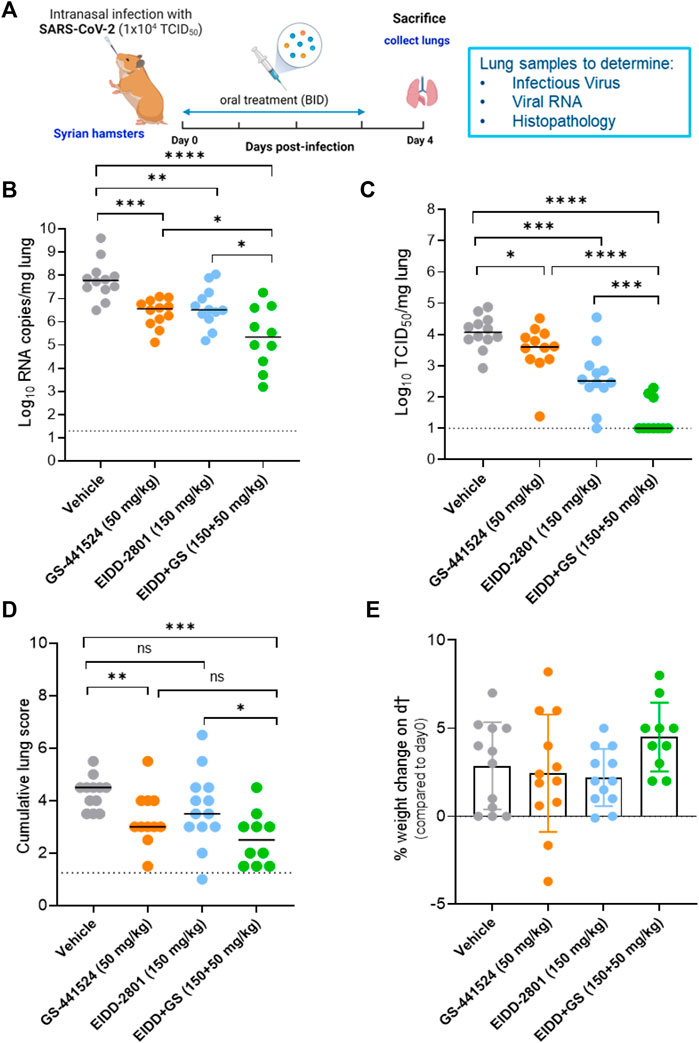
Next, we evaluated the efficacy of single versus combination treatment with molnupiravir [EIDD-2801] (150 mg/kg, oral BID) and GS-441524 (50 mg/kg, oral BID) in SARS-CoV-2-infected hamsters. Briefly, hamsters were treated with vehicle or the intended dose of each compound as single or combined therapy for four consecutive days starting just before the infection with SARS-CoV-2 beta (B.1.351) variant (Figure 2A). At day 4 post-infection (pi), the animals were euthanized and organs were collected for quantification of viral RNA, infectious virus titers and to assess the impact on lung histopathology (Figure 2A). Single treatment with molnupiravir (150 mg/kg BID) significantly reduced viral RNA and infectious virus titers in the lungs by 1.3 (p = 0.0045) and 1.6 (p = 0.0003) log10/mg lung tissue, respectively (Figure 2B, C). As monotherapy, GS-441524 treatment at dose of 50 mg/kg (BID) reduced viral RNA and infectious virus titers in the lung by 1.2 (p = 0.0001) and 0.5 (p = 0.043) log10/mg lung tissue, respectively, compared to the vehicle-treated hamsters ((Figure 2B, C). On the other hand, the combined molnupiravir/GS-441524 treatment resulted in more significant reduction of lung viral RNA loads [2.4 log10 (p < 0.0001)] compared to single treatment with molnupiravir (150 mg/kg, BID) (Figure 2B). Interestingly, the molnupiravir/GS-441524 combination resulted in a markedly enhanced reduction in infectious virus titers (∼4.0 log10 TCID50 per mg lung, p < 0.0001 as compared to molnupiravir alone) (Figure 2C). Notably, there was no detectable infectious virus in the lungs of 7 out of 10 hamsters in the combined treatment group (Figure 2C). A significant improvement in the cumulative histological lung pathology scores was observed in the combined treatment molnupiravir/GS-441524 group (median histopathology score of 2.5) as compared to the vehicle control (median score of 4.5), p = 0.0002 (Figure 2D; Supplementary Figure S1; Supplementary Table S1). The improvement in lung histopathology in the combined molnupiravir/GS-441524 treatment group was also significant compared to the single molnupiravir treatment group (median score of 3.5, p = 0.04) as well as better than the single GS-441524 treatment group (median score of 3, p = 0.13, non-significant) (Figure 2D; Supplementary Figure S1; SupplementaryTable S1). No significant weight loss or toxicity signs were observed in either the single treatment or the molnupiravir/GS-441524 combination groups compared to the vehicle-treated groups (Figure 2E). Animals treated with the combination therapy had the highest, although not significant, weight gain (mean % weight change of 4.5), compared to the other groups (Figure 2E).
FIGURE 2. Combined efficacy of Molnupiravir (EIDD-2801) with GS-441524 against SARS-CoV-2 B.1.351 variant in a hamster infection model (A) Set-up of the study (B) Viral RNA levels in the lungs of control (vehicle-treated), EIDD-2801-treated (150 mg/kg, BID), GS-441524-treated (50 mg/kg, BID) and combination-treated (EIDD-2801+GS-441524 at 150 + 50 mg/kg BID) SARS-CoV-2 (B.1.351)−infected hamsters at day 4 post-infection (pi) are expressed as log10 SARS-CoV-2 RNA copies per mg lung tissue. Individual data and median values are presented. (C) Infectious viral loads in the lungs of control (vehicle-treated), EIDD-2801-treated, GS-441524-treated and combination-treated (EIDD-2801+ GS-441524) SARS-CoV-2−infected hamsters at day 4 pi are expressed as log10 TCID50 per mg lung tissue. Individual data and median values are presented. (D) Cumulative severity score from H&E stained slides of lungs from different treatment groups. Individual data and median values are presented and the dotted line represents the median score of untreated non-infected hamsters. (E) Weight change at day 4 pi in percentage, normalized to the body weight at the time of infection. Bars represent means ± SD. Data were analyzed with the Mann−Whitney U test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns = non-significant, i.e., p < 0.05). EIDD = EIDD-2801. All data [panels (B–E)] are from two independent experiments with 12 animals per group except for the combination group (n = 10). Panel (A) created by Bioredner.
DiscussionCombinations of antiviral drugs proved to be more efficient than monotherapy for the treatment of viral infections such as caused by the human immunodeficiency virus (HIV) and the hepatitis C virus (HCV). It allows also to prevent the emergence of drug-resistant variants (Shyr et al., 2021). Here, we explored the potential efficacy of the combined GS-441524 and molnupiravir treatment against SARS-CoV-2 variants of concerns both in vitro and in infected hamsters.
In SARS-CoV-2 (BA.5) infected A549-Dual™ hACE2-TMPRSS2 cells, the combination resulted in an overall additive antiviral effect with a marked synergism at certain concentrations. In hamsters infected with the beta (B.1.351) variant and that received suboptimal doses of either GS-441524 or molnupiravir, limited to moderate antiviral effects (measured as reduction of viral titers in the lungs) were noted, respectively. However, the combined treatment of both compounds resulted in a potent antiviral effect with low or in most animals undetectable titers of infectious virus in the lungs.
Molnupiravir has been shown to increase the mutation frequency of MERS-CoV viral RNA in infected mice (Sheahan et al., 2020); we made similar observations in SARS-CoV-2-infected hamsters (Abdelnabi, et al., 2021b). These data strongly indicate that molnupiravir exerts its antiviral activity by inducing an error catastrophe of the replicating SARS-CoV-2 genome. Similar observations were made in studies in mice infected with the Venezuelan equine encephalitis virus or the influenza virus (Urakova et al., 2017; Jordan et al., 2018). GS-441524 (the active substance of remdesivir), is metabolized in cells into the pharmacologically active nucleoside triphosphate, which stalls viral RNA synthesis after being incorporated into the growing RNA strand by the RdRp (Kokic et al., 2021). Therefore, the combined actions of inducing lethal mutagenesis (molnupiravir) in the virus with chain termination (GS-441524) appears to be highly effective in reducing SARS-CoV-2 replication and in particular, infectivity.
To conclude, we identified a potent and well-tolerated combination of the nucleoside analogues, molnupiravir and GS-441524 against SARS-CoV-2. Carefully designed combinations may result in potent antiviral effects and (although not studied here) may also reduce the risk of drug-resistance development. This study lays the basis for further exploration of the combined treatment of SARS-CoV-2 infections with molnupiravir and new oral prodrug versions of remdesivir such as GS-621763 (Schäfer et al., 2022).
Data availability statementThe original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statementThe animal study was reviewed and approved by the ethics committee of animal experimentation of KU Leuven (license P065-2020).
Author contributionsRA and JN designed the studies; RA and BW performed the studies and analyzed data; RA made the graphs; BW and JN provided advice on the interpretation of data; RA and JN wrote the paper; SJ and PM provided essential reagents; RA and JN supervised the study; JN acquired funding.
FundingThis project has received funding from the Covid-19-Fund KU Leuven/UZ Leuven and the COVID-19 call of FWO (G0G4820N), the European Union’s Horizon 2020 research and innovation program under grant agreements No 101003627 (SCORE project) and Bill & Melinda Gates Foundation (BGMF) under grant agreement INV-006366. This work was also supported by the Belgian Federal Government for the VirusBank Platform. This work also has been done under the CARE project, which has received funding from the Innovative Medicines Initiative 2 Joint Undertaking (JU) under grant agreement No 101005077. The JU receives support from the European Union’s Horizon 2020 research and innovation programme and EFPIA and Bill & Melinda Gates Foundation, Global Health Drug Discovery Institute, University of Dundee. The content of this publication only reflects the author’s view and the JU is not responsible for any use that may be made of the information it contains.
AcknowledgmentsWe thank Carolien De Keyzer, Lindsey Bervoets, Thibault Francken, Birgit Voeten, Stijn Hendrickx, Niels Cremers for excellent technical assistance. We are grateful to PM for kindly providing the SARS-CoV-2 variants used in this study. We thank Prof. Jef Arnout and Dr. Annelies Sterckx (KU Leuven Faculty of Medicine, Biomedical Sciences Group Management) and Animalia and Biosafety Departments of KU Leuven for facilitating the animal studies.
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2022.1072202/full#supplementary-material
ReferencesAbdelnabi, R., Foo, C. S., Kaptein, S. J. F., Zhang, X., Langendries, L., Vangeel, L., et al. (2021a). The combined treatment of Molnupiravir and Favipiravir results in a marked potentiation of antiviral efficacy in a SARS-CoV-2 hamster infection model. BioRxiv. doi:10.1101/2020.12.10.419242
CrossRef Full Text | Google Scholar
Abdelnabi, R., Foo, C. S., Kaptein, S. J. F., Zhang, X., Do, T. N. D., Langendries, L., et al. (2021b). The combined treatment of Molnupiravir and Favipiravir results in a potentiation of antiviral efficacy in a SARS-CoV-2 hamster infection model. EBioMedicine 72, 103595. doi:10.1016/j.ebiom.2021.103595
PubMed Abstract | CrossRef Full Text | Google Scholar
Abdelnabi, R., Boudewijns, R., Foo, C. S., Seldeslachts, L., Sanchez-Felipe, L., Zhang, X., et al. (2021c). Comparing infectivity and virulence of emerging SARS-CoV-2 variants in Syrian hamsters. EBioMedicine 68, 103403. doi:10.1016/j.ebiom.2021.103403
PubMed Abstract | CrossRef Full Text | Google Scholar
Abdelnabi, R., Foo, C. S., De Jonghe, S., Maes, P., Weynand, B., and Neyts, J. (2021d). Molnupiravir inhibits replication of the emerging SARS-CoV-2 variants of concern in a hamster infection model. J. Infect. Dis. 224 (5), 749–753. doi:10.1093/infdis/jiab361
PubMed Abstract | CrossRef Full Text | Google Scholar
Abdelnabi, R., Foo, C. S., Jochmans, D., Vangeel, L., De Jonghe, S., Augustijns, P., et al. (2022). The oral protease inhibitor (PF-07321332) protects Syrian hamsters against infection with SARS-CoV-2 variants of concern. Nat. Commun. 13 (1), 719. doi:10.1038/s41467-022-28354-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Ader, F., Bouscambert-Duchamp, M., Hites, M., Peiffer-Smadja, N., Poissy, J., Belhadi, D., et al. (2022). Remdesivir plus standard of care versus standard of care alone for the treatment of patients admitted to hospital with COVID-19 (DisCoVeRy): A phase 3, randomised, controlled, open-label trial. Lancet Infect. Dis., 22(2), 209–221. doi: doi: doi:10.1016/S1473-3099(21)00485-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Boudewijns, R., Thibaut, H. J., Kaptein, S. J. F., Li, R., Vergote, V., Seldeslachts, L., et al. (2020). STAT2 signaling restricts viral dissemination but drives severe pneumonia in SARS-CoV-2 infected hamsters. Nat. Commun. 11 (1), 5838. doi:10.1038/s41467-020-19684-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Chiba, S., Kiso, M., Nakajima, N., Iida, S., Maemura, T., Kuroda, M., et al. (2022). Co-Administration of favipiravir and the remdesivir metabolite GS-441524 effectively reduces SARS-CoV-2 replication in the lungs of the Syrian hamster model. MBio 13 (1), e0304421. doi:10.1128/MBIO.03044-21
PubMed Abstract | CrossRef Full Text | Google Scholar
Consortium, W. S. T., Pan, H., Peto, R., Henao-Restrepo, A. M., Preziosi, M. P., Sathiyamoorthy, V., et al. (2020). Repurposed antiviral drugs for covid-19 — interim WHO solidarity trial results. N. Engl. J. Med. 384, 497–511. doi:10.1056/NEJMoa2023184
PubMed Abstract | CrossRef Full Text | Google Scholar
Cox, R. M., Wolf, J. D., and Plemper, R. K. (2020). Therapeutically administered ribonucleoside analogue MK-4482/EIDD-2801 blocks SARS-CoV-2 transmission in ferrets. Nat. Microbiol. 6, 11–18. doi:10.1038/s41564-020-00835-2
PubMed Abstract | CrossRef Full Text | Google Scholar
Goldman, J. D., Lye, D. C. B., Hui, D. S., Marks, K. M., Bruno, R., Montejano, R., et al. (2020). Remdesivir for 5 or 10 Days in patients with severe covid-19. N. Engl. J. Med. 383 (19), 1827–1837. doi:10.1056/NEJMoa2015301
PubMed Abstract | CrossRef Full Text | Google Scholar
Gottlieb, R. L., Vaca, C. E., Paredes, R., Mera, J., Webb, B. J., Perez, G., et al. (2021). Early remdesivir to prevent progression to severe covid-19 in outpatients. N. Engl. J. Med. 386 (4), 305–315. doi:10.1056/NEJMoa2116846
PubMed Abstract | CrossRef Full Text | Google Scholar
Hammond, J., Leister-Tebbe, H., Gardner, A., Abreu, P., Bao, W., Wisemandle, W., et al. (2022). Oral Nirmatrelvir for high-risk, nonhospitalized adults with covid-19. N. Engl. J. Med. 386 (15), 1397–1408. doi:10.1056/NEJMoa2118542
PubMed Abstract | CrossRef Full Text | Google Scholar
Ianevski, A., He, L., Aittokallio, T., and Tang, J. (2017). SynergyFinder: A web application for analyzing drug combination dose-response matrix data. Bioinformatics 33 (15), 2413–2415. doi:10.1093/bioinformatics/btx162
PubMed Abstract | CrossRef Full Text | Google Scholar
Johnson, M. G., Puenpatom, A., Moncada, P. A., Burgess, L., Duke, E. R., Ohmagari, N., et al. (2022). Effect of molnupiravir on biomarkers, respiratory interventions, and medical services in COVID-19 : A randomized, placebo-controlled trial. Ann. Intern. Med. 175, 1126–1134. doi:10.7326/M22-0729
PubMed Abstract | CrossRef Full Text | Google Scholar
Jonsdottir, H. R., Siegrist, D., Julien, T., Padey, B., Bouveret, M., Terrier, O., et al. (2022). Molnupiravir combined with different repurposed drugs further inhibits SARS-CoV-2 infection in human nasal epithelium in vitro. Biomed. Pharmacother. 150, 113058. doi:10.1016/j.biopha.2022.113058
PubMed Abstract | CrossRef Full Text | Google Scholar
Jordan, P. C., Liu, C., Raynaud, P., Lo, M. K., Spiropoulou, C. F., Symons, J. A., et al. (2018). Initiation, extension, and termination of RNA synthesis by a paramyxovirus polymerase. PLoS Pathog. 14 (2), e1006889. doi:10.1371/journal.ppat.1006889
PubMed Abstract | CrossRef Full Text | Google Scholar
Kaptein, S. J. F., Jacobs, S., Langendries, L., Seldeslachts, L., ter Horst, S., Liesenborghs, L., et al. (2020). Favipiravir at high doses has potent antiviral activity in SARS-CoV-2−infected hamsters, whereas hydroxychloroquine lacks activity. Proc. Natl. Acad. Sci. U. S. A. 117 (43), 26955–26965. doi:10.1073/pnas.2014441117
PubMed Abstract | CrossRef Full Text | Google Scholar
Kokic, G., Hillen, H. S., Tegunov, D., Dienemann, C., Seitz, F., Schmitzova, J., et al. (2021). Structure of replicating SARS-CoV-2 polymerase. Nat. Commun. 12 (1), 154–156. doi:10.1038/s41586-020-2368-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, Y., Cao, L., Li, G., Cong, F., Li, Y., Sun, J., et al. (2021). Remdesivir metabolite GS-441524 effectively inhibits SARS-CoV-2 infection in mouse models. J. Med. Chem. 65, 2785–2793. doi:10.1021/acs.jmedchem.0c01929
PubMed Abstract | CrossRef Full Text | Google Scholar
Pedersen, N. C., Perron, M., Bannasch, M., Montgomery, E., Murakami, E., Liepnieks, M., et al. (2019). Efficacy and safety of the nucleoside analog GS-441524 for treatment of cats with naturally occurring feline infectious peritonitis. J. Feline Med. Surg. 21 (4), 271–281. doi:10.1177/1098612X19825701
PubMed Abstract | CrossRef Full Text | Google Scholar
Reed, L. J., and Muench, H. (1938). A simple method of estimating fifty percent endpoints. Am. J. Hyg. 27 (3), 493–497. doi:10.1093/oxfordjournals.aje.a118408
CrossRef Full Text | Google Scholar
Rosenke, K., Hansen, F., Schwarz, B., Feldmann, F., Haddock, E., Rosenke, R., et al. (2021). Orally delivered MK-4482 inhibits SARS-CoV-2 replication in the Syrian hamster model. Nat. Commun. 12 (1), 2295. doi:10.1038/s41467-021-22580-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Schäfer, A., Martinez, D. R., Won, J. J., Meganck, R. M., Moreira, F. R., Brown, A. J., et al. (2022). Therapeutic treatment with an oral prodrug of the remdesivir parental nucleoside is protective against SARS-CoV-2 pathogenesis in mice. Sci. Transl. Med. 14 (643), eabm3410. doi:10.1126/scitranslmed.abm3410
PubMed Abstract | CrossRef Full Text | Google Scholar
Sheahan, T. P., Sims, A. C., Zhou, S., Graham, R. L., Pruijssers, A. J., Agostini, M. L., et al. (2020). An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 12 (541), eabb5883. doi:10.1126/SCITRANSLMED.ABB5883
PubMed Abstract | CrossRef Full Text | Google Scholar
Shyr, Z. A., Cheng, Y. S., Lo, D. C., and Zheng, W. (2021). Drug combination therapy for emerging viral diseases. Drug Discov. Today 26 (10), 2367–2376. doi:10.1016/j.drudis.2021.05.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Toots, M., Yoon, J. J., Cox, R. M., Hart, M., Sticher, Z. M., Makhsous, N., et al. (2019). Characterization of orally efficacious influenza drug with high resistance barrier in ferrets and human airway epithelia. Sci. Transl. Med. 11 (515), eaax5866. doi:10.1126/scitranslmed.aax5866
PubMed Abstract | CrossRef Full Text | Google Scholar
Urakova, N., Kuznetsova, V., Crossman, D. K., Sokratian, A., Guthrie, D. B., Kolykhalov, A. A., et al. (2017). β-D- N 4 -hydroxycytidine is a potent anti-alphavirus compound that induces a high level of mutations in the viral genome. J. Virology 92 (3), 019655–e2017. doi:10.1128/jvi.01965-17
CrossRef Full Text | Google Scholar
Wahl, A., Gralinski, L., Johnson, C., Yao, W., Kovarova, M., Dinnon, K., et al. (2020). Acute SARS-CoV-2 infection is highly cytopathic, elicits a robust innate immune response and is efficiently prevented by EIDD-2801. Res. Square, 80404. doi:10.21203/rs.3.rs-80404/v1
PubMed Abstract | CrossRef Full Text | Google Scholar
Warren, T. K., Jordan, R., Lo, M. K., Ray, A. S., Mackman, R. L., Soloveva, V., et al. (2016). Therapeutic efficacy of the small molecule GS-5734 against Ebola virus in rhesus monkeys. Nature 531 (7594), 381–385. doi:10.1038/nature17180
PubMed Abstract | CrossRef Full Text | Google Scholar
Williamson, B. N., Feldmann, F., Schwarz, B., Meade-White, K., Porter, D. P., Schulz, J., et al. (2020). Clinical benefit of remdesivir in rhesus macaques infected with SARS-CoV-2. Nature 585 (7824), 273–276. doi:10.1038/s41586-020-2423-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Xie, J., and Wang, Z. (2021). Can remdesivir and its parent nucleoside GS-441524 be potential oral drugs? An in vitro and in vivo DMPK assessment. Acta Pharm. Sin. B 11 (6), 1607–1616. doi:10.1016/j.apsb.2021.03.028
PubMed Abstract | CrossRef Full Text | Google Scholar
Yan, V. C., and Muller, F. L. (2020). Advantages of the parent nucleoside GS-441524 over remdesivir for covid-19 treatment. ACS Med. Chem. Lett. 11 (7), 1361–1366. doi:10.1021/acsmedchemlett.0c00316
PubMed Abstract | CrossRef Full Text | Google Scholar
Yan, V. (2021). First-in-Woman safety, tolerability, and pharmacokinetics of orally administered GS-441524: A broad-spectrum antiviral treatment for COVID-19. OSF Preprints.
留言 (0)