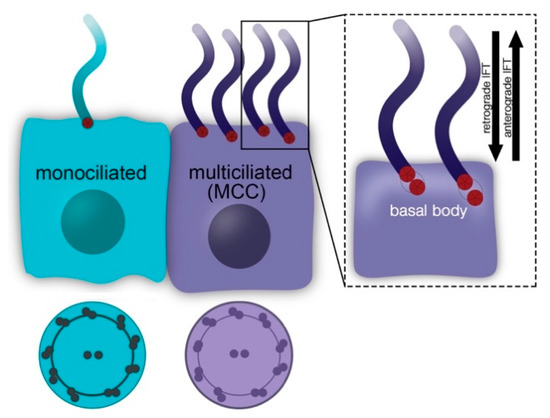
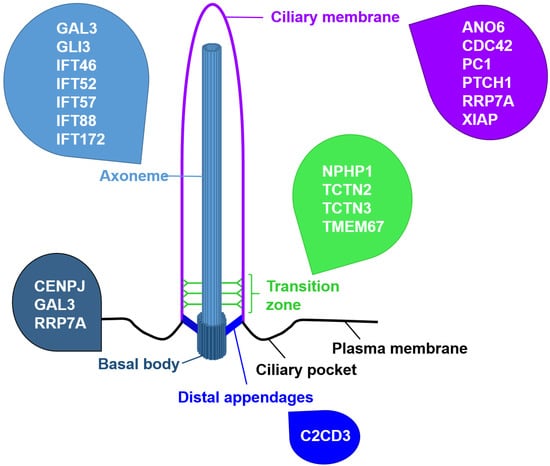
To shed further light on the relationship between primary cilia and apoptosis, it might be useful to focus on other ciliary proteins than the IFT proteins. In this context, we suggest investigating proteins that localise to the ciliary base. Schock et al. revealed that apoptosis is unchanged in talpid2 (ta2) mutant avian embryos suffering from an orofaciodigital syndrome-like ciliopathy [
50]. However, Dvorak and Fallon detected reduced cell death in ta2 mutant avian embryos [
51]. Since the investigations of Schock et al. were limited to cranial neural crest cells and the analyses of Dvorak and Fallon refer to limb buds and limbs, cell type-specific differences should be taken into account in the deciphering of the relationship between primary cilia and cell death. Mutations in C2 Domain-Containing Protein 3 (C2CD3) have been shown to be the causal genetic lesion for the avian ta2 mutant [
52]. C2CD3 localises to the distal appendages that anchor the BB to the cell membrane [
53,
54]. It is required for ciliogenesis, for recruiting proteins to the BB, for removing protein from the BB, for the docking of vesicles to the BB, for distal appendage assembly, for controlling centriole elongation, and for regulating HH signalling in a tissue-specific manner [
54,
55,
56,
57]. In addition to the BB protein C2CD3, some proteins that localise to the TZ are associated with cell death. Several studies revealed that the deficiency of the MKS/B9 module components Tectonic-2 (TCTN2), Tectonic-3 (TCTN3), and Transmembrane Protein 67 (TMEM67 alias MKS3) in mice and rats increases apoptosis. In Tctn2-negative mouse embryos, increased apoptosis was detected in the ventral neuroectoderm and facial ectoderm [
58]. Loss of TCTN3 engenders raised apoptosis in the brain of murine embryos [
59]. In Tmem67 mutant rats, elevated apoptosis was found in the eyes [
60]. In the TZ, TCTN2 ensures the proper amount of TMEM67 [
61]. Previously, it was shown that TCTN2, TCTN3, and TMEM67 control ciliary gating and, hence, ciliary protein composition [
62]. During the investigations focussing on the link between TCTN2, TCTN3 and apoptosis, it was hypothesised that these proteins negatively regulate apoptosis by controlling HH signalling. In both Tctn2−/− and Tctn3−/− mouse embryos, HH signalling was reduced [
58,
59]. Importantly, the decrease in Patched-1 (PTCH1), a receptor of the HH ligand and a negative regulator of HH signalling, within Tctn2-negative mouse embryos rescued apoptosis in the ventral neuroectoderm and facial ectoderm whereby verifying the hypothesis [
58]. Independently of the studies involving Tctn mutant mice, Aoto and Trainor revealed a regulation of apoptosis by PTCH1 via controlling CASP9 activity [
63]. Mechanistically, the association of PTCH1 with X-linked Inhibitory Apoptosis Protein (XIAP) is of great importance and both proteins localise to primary cilia. To our knowledge, XIAP is the only protein involved in the known cell death signalling pathways that localises to primary cilia. In the absence of the HH ligand, PTCH1 separates from XIAP, and XIAP leaves the cilium. In this state, the C-terminal domain of PTCH1 is processed in a CASP9-dependent manner, which results in mitochondrial dysfunction and apoptosis. In the presence of the HH ligand, it binds to PTCH1 which subsequently exits the cilium and gets degraded [
63]. Combining these results with the above-mentioned studies, TCTN2 and most likely TCTN3 as well might affect apoptosis by regulating HH signalling. However, to date it is unknown where TCTN2 and TCTN3 act in the HH signalling pathway and how these proteins affect apoptosis. Following the results of Aoto and Trainor, it could be assumed that the amount of PTCH1 might be higher in the Tctn2−/− and the Tctn3−/− state. However, Sang and colleagues did not detect an altered PTCH1 amount in Tctn2−/− mouse embryonic fibroblasts (MEFs) (Sang et al., 2011), but they showed that TCTN2 is crucial for the processing and function of the HH signalling mediator Glioma-Associated Oncogene Family Zinc Finger 3 (GLI3) (Sang et al., 2011). Full length GLI3 either functions as a transcriptional activator or, after getting processed by partial proteolysis, acts as a repressor that regulates HH target gene expression [
64]. Previous work elucidated an involvement of GLI3 in the regulation of apoptosis [
65,
66,
67]. This raises the intriguing question if TCTN2 and TCTN3 might govern apoptosis via GLI3. In order to approach this question, we suggest analysing other TZ proteins involved in the regulation of GLI3 processing and function. As a prominent example, RPGRIP1L functions as a component of the NPHP1-4-8 module and controls the processing of GLI3 by regulating proteasomal activity at the ciliary base [
68]. Moreover, it was suggested that RPGRIP1L governs the amount of the GLI3 activator [
69]. Contrary to the situation in the absence of TCTN2 and TCTN3, loss of RPGRIP1L does not result in altered apoptosis in mouse embryonic hearts, in mouse embryonic hypothalami, or in the mouse adult central nervous system [
69,
70]. Subject to the possible existence of a cell type-specific relationship between primary cilia and apoptosis, these data indicate that TCTN2 and TCTN3 do not govern apoptosis by regulating GLI3. Furthermore, loss of TCTN2 and TCTN3 results in a reduction of Smoothened (SMO), another mediator of HH signalling which functions upstream of GLI3, reflecting impaired ciliary gating [
62]. Interestingly, although RPGRIP1L also governs ciliary gating, SMO enters the cilium properly in the absence of RPGRIP1L [
9,
68]. This difference might be related to the different effects of TCTN2/TCTN3 and RPGRIP1L on apoptosis. In endothelial cells, it was elucidated that SMO negatively regulates apoptosis whereby supporting this hypothesis [
71]. However, this line of argument seems to be countered by the fact that TMEM67 deficiency does not alter the ciliary SMO amount, although mutation of TMEM67 leads to enhanced apoptosis in the eyes of rats [
60,
62]. Considering that the connecting cilium in photoreceptors differs from the cilia of other cells, the SMO hypothesis has to be carefully tested by including cell type-specific effects into the experimental design [
15]. Recently, it was shown that the TZ also functions as a gatekeeper for ciliary exit of retrograde IFT trains [
72]. In this context, it is not surprising that TCTN2 ensures the proper ciliary localisation of IFT88 as TCTN2 is deeply involved in the regulation of ciliary gating [
61,
62]. Since both the loss of TCTN2 and the loss of IFT88 can result in increased apoptosis [
42,
43,
44,
58], it is conceivable that TCTN2 governs cell survival via IFT88. However, the proper localisation of IFT88 to cilia also depends on RPGRIP1L [
61] but—as mentioned before—loss of RPGRIP1L does not provoke enhanced apoptosis [
69,
70]. Moreover, IFT88 (component of the IFT-B complex) and IFT139 (component of the IFT-A complex) localise independently of TCTN1, another TZ protein that controls ciliary gating [
62]. Interestingly, depletion of TCTN1 promotes apoptosis [
73,
74]. According to these data, it seems unlikely that TZ proteins regulate apoptosis via ensuring the proper ciliary localisation of IFT proteins. Another possibility for how TCTN3 controls apoptosis is its physical interaction with Nephrocystin 1 (NPHP1), another component of the NPHP1-4-8 module. Depletion of NPHP1 led to elevated apoptosis and, in the absence of TCTN3, the amount of NPHP1 is decreased in mouse embryonic brains [
59,
75]. Interestingly, it has been suggested that NPHP1 regulates apoptosis in cooperation with Polycystin 1 (PC1), but the mechanism of this regulation remains elusive [
75]. PC1 deficiency causes polycystic kidney disease, and elevated apoptosis is one feature of polycystic kidney disease [
76]. Although apoptosis is unaltered in the absence of RPGRIP1L, it was reported that RPGRIP1L deficiency reduces the ciliary amount of NPHP1 querying a potential regulation of apoptosis by TCTN3 via NPHP1 [
12,
70]. To conclude, several ciliary base proteins have an impact on apoptosis, but the mechanism underlying the regulation of apoptosis by these proteins needs to be elucidated. We suggest regulation via HH signalling (especially via the HH signalling components PTCH1, GLI3, and SMO), via IFT proteins or via NPHP1. In every case, there are arguments for and against this hypothesis. To complete the puzzle of how TZ proteins act to control apoptosis, more research is required.






留言 (0)