
記住我



According to WHO Classification of Breast Tumors, 5% of invasive breast carcinomas (BC) present features of both invasive carcinoma of no special type (IBC-NST) and invasive lobular carcinomas (ILC), an entity called invasive ductolobular carcinoma (IDLC).1 Whereas some studies have considered that IDLC differs from ILC and IBC-NST,2–4 others consider that IDLC can be separated into IBC-NST-like and ILC-like cases and does not represent a molecularly distinct subtype.5 Discrepancies among studies could be partially because of differences in the selection of cases and whether or not the studies included samples from the 2 components.
The diagnosis of IDLC is currently based on morphology and basically describes a growth pattern. A major impact on IDLC definition relies on the definition of ILC, which is currently also based on morphologic criteria. However, by incorporating immunohistochemical criteria (loss or aberrant E-cadherin expression in the lobular component), the diagnosis of both ILC and IDLC may be more robust.3 A more accurate classification of IDLC will improve our understanding of the disease and whether or not this entity has specific clinical and biological features.
The relationship between the 2 invasive components of IDLC has not been fully elucidated. IDLC could represent a collision tumor (IBC-NST and ILC coexisting in a common area) or a single tumor in which one of the components arises from the other. Some studies, based on the molecular analysis of a relative low number of cases, have suggested a clonal relationship between the 2 components6–8 with a common ancestor, and that the lobular component derives from the ductal component.7
In this study, the 2 components of 20 IDLCs were analyzed by massive parallel sequencing and fluorescent in situ hybridization (FISH) in order to gain insights into their clonal relationships. Only cases with preserved E-cadherin expression in the ductal component and loss of E-cadherin in the lobular component were selected. By incorporating this criterion, we avoided heterogeneity because of the use of exclusive morphologic criteria. Our study indicated that most IDLC occurred by progression of an IBC-NST in which a clonal cell population lost E-cadherin expression and acquired a lobular phenotype.
METHODS Case SelectionCases were selected from the Pathology Departments of 3 hospitals: Ramón y Cajal University Hospital (Madrid, Spain), La Paz University Hospital (Madrid, Spain) and Virgen del Rocío University Hospital (Sevilla, Spain). Cases were included if they had a typical morphologic pattern of IDLC, absence of E-cadherin expression in the lobular component and preserved E-cadherin expression in the ductal component. In addition, cases were selected if there was tissue available for additional immunohistochemical and molecular studies.
Clinical data were obtained from clinical records. Histologic typing was performed according to WHO recommendations and cases were graded according to the 3-tiered Nottingham histologic grading system.
Approval for the study was obtained from the Local Ethics Committee (Ramón y Cajal Research Ethics Committee reference 223/18).
ImmunohistochemistryAll cases underwent a new immunohistochemical study. The antibodies used are shown in Supplementary Table S1, Supplemental Digital Content 1, https://links.lww.com/PAS/B382. Immunostaining was performed using the EnVision detection system (K5007; Dako, Glostrup, Denmark). A cut-off value of 1% was used to define estrogen receptor (ER) and progesterone receptor (PR) positivity. Ki67 index was defined as the percentage of positive tumor cells independently of intensity. HER2 expression was interpreted according to 2018 American Society of Clinical Oncology and the College of American Pathologists (ASCO-CAP) guidelines.9 HER2 equivocal cases (2+) underwent FISH, using the Path Vision HER2 DNA Probe Kit (Abbot Laboratories, City, Country) and the results were interpreted according to 2018 ASCO-CAP guidelines.9
Tumors expressing ER and/or PR were classified as Luminal. Luminal tumors with Ki67 <15% were classified as Luminal A, those with Ki67 >15% as Luminal B,10 and those that also were HER2 positive as Luminal HER2. ER and PR negative tumors that were HER2 positive were classified as HER2 enriched, and those negative for ER, PR and HER2 as triple negative.
Massive Parallel SequencingTo obtain DNA mainly from tumor cells, samples from different lesions were obtained by “punching” paraffin blocks in selected areas previously marked on hematoxylin-eosin slides. The QIAamp DNA FFPE Tissue Kit (Qiagen, Valencia, CA) was used to extract DNA from all samples. The quality of DNA was measured using TapeStation (Agilent 2200 TapeStation; Santa Clara, CA), whereas quantification was performed by QUBIT 2.0. (Thermo Fisher Scientific Qubit 2.0; Fluorometer, Waltham, MA).
A custom gene panel was designed using the SureDesign platform by Agilent Tech. (Santa Clara) to consistently target 38 genes (AKT1, ARID1A, ARID1B, BRCA1, BRCA2, CASP8, CCND1, CDH1, CTNNA1, CTNNB1, CTNND1, ERBB2, ESR1, FGFR1, GATA3, GRB7, GSDMB, JUP, MAP2K4, KRAS, MAP3K1, MLL3, MYC, NCOR1, NF1, PGAP, PIK3CA, PNMT, PTEN, RB1, SF3B1, STARD3, TBX3, TCAP, TP53, VGLL1, ZNF217, ZNF703). This panel is a modification of the previously reported panel, 11 to include the complete sequences of genes coding for α-catenin (CTNNA1), β-catenin (CTNNB1), p120 catenin (CTNND1), and γ-catenin (JUP). For library construction, a modified protocol for Agilent SureSelect was selected, based on the enzymatic fragmentation of DNA and subsequent probe-mediated hybridization capture.12 Sequencing of equimolar libraries was performed using the Miseq v2 2×150 bp method by Illumina Inc. (San Diego, CA).
Bioinformatics analysis was carried out as previously reported11 using a specific pipeline using Novoalign13 as an aligner and VarScan as a variant-caller, with no filters. Variant annotation was performed using the VEP from Ensembl version 88,14 which corresponds to GRCh38-hg20 version of the human reference genome. Variants were latterly filtered using the functional information (selecting only deleterious variants), the variant allele frequency (>0.05) and the strand-bias from both the variant and the reference allele. Finally, visual inspection was performed as the final selection criterion. Eight ILC (5 with CDH1 mutations and 3 without CDH1 mutations) previously studied11 were included as controls.
Fluorescent In Situ HybridizationSince our panel was not able to accurately detect gene copy number variations, we used FISH to evaluate CCND1, MYC, FGFR1, and MDM4, the genes most frequently amplified in BC, as previously reported.15 The following probes were used: ZytoLight SPEC CCND1/CEN11, ZytoLight SPEC MYC/CEN8, ZytoLight SPEC FGFR1/CEN8, and ZytoLight SPEC MDM4/1p12 dual color Probe Kit (Zytovision GmbH, Bremen, DE) FISH slides were observed with a fluorescence microscope at ×100 with immersion oil. A detailed scoring of at least 40 neoplastic cells per sample was performed, evaluating separately 20 cells of each tumoral component. Ductal and lobular areas were previously marked on FISH slides to ease their recognition with the fluorescent microscope. Amplification was considered when the tumor cell population had at least twice as many gene signals than centromere signals of the respective chromosome (ratio ≥2), and polysomy when the average of centromere signals on tumor cells were >3.
For CDH1 gene alterations, we used CDH1 FISH probe Spectrum Orange (Empire Genomics, Buffalo, NY) with Vysis CEP16 (D16Z3) Spectrum Green Probe (Abbott Laboratories, Chicago, IL). A detailed scoring per sample of at least 40 neoplastic cells of each area with positive and negative expression of E-cadherin was performed. Heterozygous deletion was considered when equal or more than 20% of evaluated nuclei presented at least 2 centromere signals and just 1 CDH1 signal. Homozygous deletion was considered when equal or more than 20% of evaluated nuclei presented no copy of CDH1 gene.
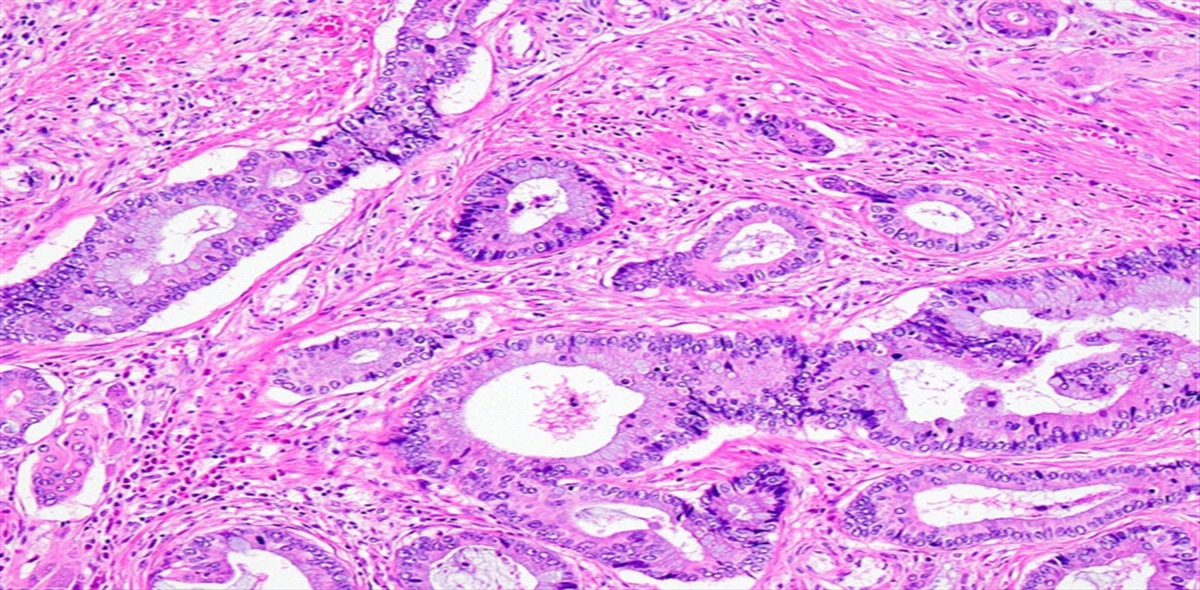
RESULTS Clinicopathologic FeaturesThe study included 40 tumor samples from 20 patients. Fifteen additional IDLC were excluded because of the poor quality of DNA or sequencing since the study included cases from 2004 to 2019. All tumors developed in females. Clinicopathologic features are presented in Table 1 and Supplementary Table S2, Supplemental Digital Content 2, https://links.lww.com/PAS/B383; and representative histopathologic images in Figure 1. The median age was 57.5 years and median tumor size was 2.65 cm. The percentage of the lobular component ranged from 15% to 70%, being predominant in only 4 cases (20%). All lobular components and 18 (90%) ductal components were histologic grade 2. Areas of in situ ductal carcinoma and in situ lobular carcinoma were observed in 13 and 6 tumors, respectively. Only 1 tumor had both types of in situ carcinoma. Regarding immunohistochemical features, 18 tumors were ER positive and 16 PR positive. Ten tumors were HER2 negative 0+, 1 tumor was HER2 negative 1+, and 7 tumors HER2 equivocal 2+. None of the equivocal cases showed HER2 amplification by FISH. Only 2 tumors were triple negative. These IHC characteristics did not differ between the ductal and lobular components. After a median follow up of 48 months (range: 12 to 162 mo), 15 patients remain alive without disease, 1 alive with disease and 4 have died of disease.
TABLE 1 - Clinicopathologic Features Category n (%) Age (years), median (IQR) 57.5 (19.5) Menopause 15 (75) Size (cm), median (IQR) 2.65 (1.6) Nottingham grade Grade 1 0 Grade 2 18 (90) Grade 3 2 (10) pT pT1a 0 pT1b 1 (5) pT1c 5 (25) pT2 12 (60) pT3 2 (10) pN pN0 7 (35) pN+ 10 (50) pNx 3 (15) Associated DCIS 13 (65) Associated LCIS 6 (30) ER positive (>1%) 18 (90) PR positive (>1%) 16 (80) HER2 positive 0 ki67 <15% 13 (65) Subrogated molecular subtypes Luminal A 14 (70) Luminal B 4 (20) Luminal B HER2 0 Triple negative 2 (10) HER2 enriched 0 Follow up Alive without disease 15 (75) Alive with disease 1 (5) Death from illness 4 (20)IQR indicates interquartile range.
 FIGURE 1:
FIGURE 1: A and B, Invasive ductolobular carcinoma (T5) where an in situ ductal component is intermingled with both ductal and lobular invasive components. A, hematoxylin-eosin (H&E) staining. B, E-cadherin immunohistochemistry. C and D, Invasive ductolobular carcinoma (T7) where the ductal component shows micropapillary architecture. C, H&E staining. D, E-cadherin immunohistochemistry. E and F, Metastasis of invasive ductolobular carcinoma in an axillary lymph node. Note the presence of both components. E, H&E staining. F, E-cadherin immunohistochemistry.
Molecular AlterationsAll somatic pathogenic and likely pathogenic somatic variants and gene amplifications are presented in Supplemental Table S3, Supplemental Digital Content 3, https://links.lww.com/PAS/B384. Mutations and/or amplifications were detected in 18 of 20 tumors (90%). Somatic mutations were present in 15 tumors (75%) and gene amplification in 8 of 18 tumors analyzed (44%) (Fig. 2). The most frequent alterations were mutations in PIK3CA (10 tumors, 50%), CDH1 (9 tumors, 45%) and ARID1B (3 tumors, 15%), and amplifications in CCND1 (5 tumors, 28%) and MDM4 (3 tumors, 17%).
 FIGURE 2:
FIGURE 2: Oncoplot showing the mutations and amplifications present in each component of each case.
In 16 tumors (80%), common driver alterations were detected in the ductal and lobular components, suggesting a clonal relationship (Fig. 3). The same PIK3CA mutation was detected in both components in 6 tumors (one of them with ARID1B mutation and other with CCND1 amplification as additional common drivers); the same TP53 mutation in 2 tumors (1 with an additional common CCND1 amplification), GATA3 (1 tumor), AKT1 (1 tumor) and ARID1B (1 tumor) were observed in 5 additional cases. Moreover, the lobular and ductal components in 5 tumors shared common amplifications in CCND1/MDM4 (2 tumors), CCND1 (1 tumor), MDM4 (1 tumor), and MYC (1 tumor).
 FIGURE 3:
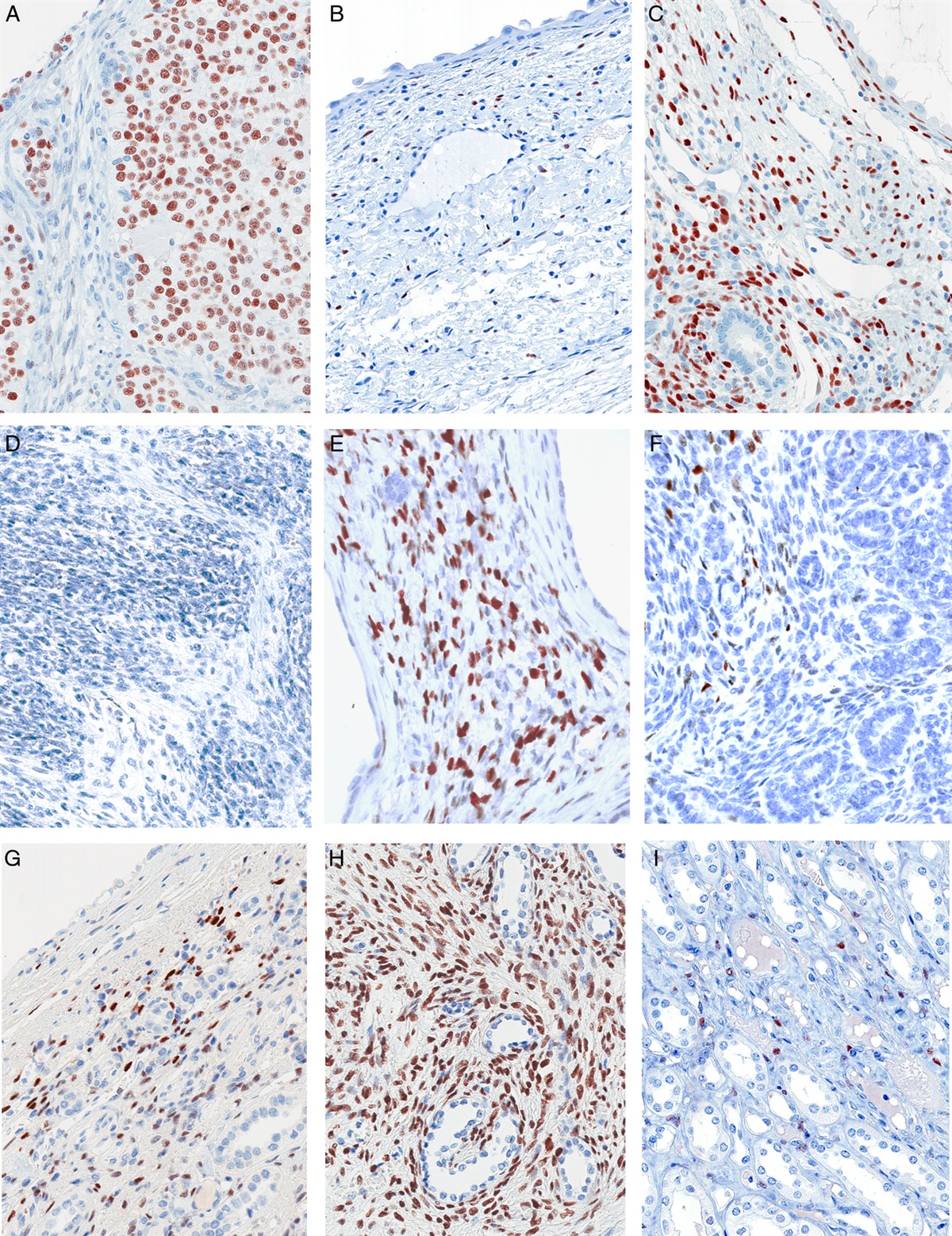
FIGURE 3: Histopathologic images of mixed ductal and lobular carcinoma in a clonal related case (Case T_3). A, Hematoxylin-eosin-staining of a region where invasive lobular component is intermingled between ductal nests. B, Immunohistochemical staining for E-cadherin in the previously depicted hematoxylin-eosin area. The ductal component shows E-cadherin membrane staining while the lobular one is negative. C, Immunohistochemical staining for p53 showing moderate to intense staining in both components. D, Clonal mutation in TP53 is found in both components, whereas a CDH1 mutation is only detected in the lobular component (E) and a PI3KCA mutation is just found in the ductal one (F).
In 6 of these tumors (37.5%) with common drivers, there were no other alterations in the ductal or lobular component. The remaining 9 tumors showed additional mutations in the lobular component. Eight of them presented mutations in CDH1 (one with an additional MAP3K1 mutation, another with an ARID1B mutation, a third with an AKT1 mutation and another with a PTEN mutation) and 1 in PIK3CA and ARID1A. Regarding the ductal component, 6 presented additional mutations. Two showed PIK3CA mutations (1 with and additional PTEN mutation), 1 ARID1A and NF1 mutation, 1 ERBB2 mutation, 1 PTEN mutation, and 1 MYC amplification (Fig. 4).
 FIGURE 4:
FIGURE 4: Evolutionary trees of different mixed ductal and lobular carcinomas.
There was no evidence of clonality in 4 IDLC. In 2 tumors (50%), no mutations nor amplifications were detected. In 1 tumor, only a BRCA2 mutation was detected in the ductal component; and a divergent pattern of mutations was observed in the remaining case, with mutations in PIK3CA and GATA3 in the ductal component, but only a mutation of CDH1 in the lobular component (Fig. 5).
 FIGURE 5:
FIGURE 5: Histopathologic images of mixed ductal and lobular carcinoma with different mutations in both components (case T_1). A, Hematoxylin-eosin-staining of a region where lobular and ductal components are adjacent to each other. B, Immunohistochemical staining for E-cadherin in the previously depicted hematoxylin-eosin area. The ductal component shows E-cadherin membrane staining while the lobular one is negative. C, Immunohistochemistry for estrogen receptor showing intense staining in both components. D, CDH1 mutation is present just in the lobular component, whereas GATA3 (E) and PI3KCA (F) mutations are just found in ductal one.
Since the frequency of CDH1 mutations in the lobular components in this series was lower than expected (45%), we investigated if homozygous CDH1 deletion could be the cause of E-cadherin loss of expression in some cases by FISH. CDH1 FISH analysis was performed in 14 tumors (7 with CDH1 mutations and 7 without mutations). All ductal areas had heterozygous loss of CDH1. No additional changes were seen in the lobular component of 7 cases without mutations. Chromosome 16 monosomy was observed in 2 cases with a mutation. Another case (T_6) in which 3 areas where sequenced, 1 lobular area showed chromosome 16 disomic cells with homozygous CDH1 loss while other lobular area showed chromosome 16 monosomy and CDH1 mutation (Fig. 6).
 FIGURE 6:
FIGURE 6: Different causes of E-cadherin loss in T_6. A, Evolutionary tree, with PIK3CA mutation as common driver. B and D, T_6 CDH1 FISH, with CDH1 signal in red and chromosome 16 signal in green. B, The ductal component (D) shows an additional PTEN mutation and CDH1 heterozygous loss. C, One lobular component (L1) shows chromosome 16 monosomy and CDH1 mutation. D, A second lobular component (L2) shows CDH1 homozygous loss.
DISCUSSIONIn this study, the clonal relationship between the ductal and lobular components of 20 IDLC was analyzed, and clonal relatedness was found in 80% of cases. One important characteristic of our study is its design. We selected as IDLC only those cases in which the lobular component had a complete loss of E-cadherin expression. Recently, 2 large BC clinical trials have indicated suboptimal concordance between BC subtypes determined by local versus central pathology, with the histopathologic diagnosis of ILC ranging from 60% to 66% based on morphology alone.16 This is more frequently due the fact that E-cadherin-positive IBC-NST with trabecular growth pattern are frequently diagnosed as ILC. For this reason, some authors have proposed the use of morphomolecular criteria for the diagnosis of ILC.3 Accordingly, ILC should be diagnosed in tumors with a typical morphology (classic or variants) of ILC with complete loss of E-cadherin expression. In tumors with a characteristic morphology that have E-cadherin expression, additional criteria should be used, such as an abnormal pattern of expression of E-cadherin (cytoplasmic, fragmented), abnormal expression of catenins (especially cytoplasmic p120), evidence of pathogenic CDH1 mutations or the association with in situ lobular carcinoma in multiple lobules within the tumor. Although this proposal has not still been adopted, some studies have reported that subtyping of BC as ILC achieves an almost perfect agreement with a predefined reference standard, if the assessment is supported by E-cadherin immunohistochemistry.16
The criteria to define ILC influence the diagnosis of IDLC. Without the inclusion of E-cadherin loss in the lobular component as a criterion, the diagnosis of IDLC describes a growth pattern that can include different situations. By applying the criteria of loss of E-cadherin in the lobular component, a more homogeneous population of tumors was selected and the bias of including ductal carcinomas with areas of lobular-like growth pattern or lobular carcinomas with tubular elements was avoided. Other studies analyzing the molecular features of IDLC have not taken into account these criteria.5,7 For example, McCart Reed et al7 reported that just 17.6% of mixed BC (9/51) had lost E-cadherin expression, with most showing aberrant staining.
Our results indicated that in most tumors, the lobular component arose from the ductal component after E-cadherin loss. Our results confirmed and expanded the observations of a previous study by McCart Reed et al7 which molecularly analyzed fewer invasive mixed BC (n=8). This suggestion is supported by: (1) 80% of tumors in our series shared a common driver in both the ductal and lobular component; (2) CDH1 mutations or homozygous CDH1 loss were only present in the lobular component, suggesting mutation acquisition during progression in ductal tumors with E-cadherin LOH, as demonstrated by FISH. It appears improbable that, even in the absence of CDH1 mutation, a tumor with E-cadherin loss, reverted this feature and acquire both intercellular adhesion and a “ductal” phenotype.
Since most tumors were Luminal, PIK3CA was the most common mutated gene, and PIK3CA mutation the most common shared molecular alteration between both components. Moreover, other common drivers of BC, such as amplification of CCND1 or TP53 mutations, were also shared by ductal and lobular areas.
There was no evidence of clonality between the ductal and lobular components in only 4 tumors in this series. Although the panel used in this study included the most commonly mutated genes in BC, we cannot rule out the existence of alterations in other genes infrequently mutated in BC and/or the presence of gene amplifications or deletions in genes other than CCND1, MYC, MDM4, and FGFR1.
In addition to common drivers, we also observed unique mutations of each component. Of special importance were those actionable mutations according to SCAT criteria (PIK3CA, AKT1, and ERBB2).17 In 3 patients (T1, T3, and T20), a PIK3CA mutation was present only in the ductal component, whereas in T7 it was only present in the lobular component. Of note, T_5 had different actionable mutations in both components: ERBB2 in the ductal and AKT1 in the lobular. These results suggested that molecular heterogeneity in IDLC is not only limited to CDH1 and that both components should be molecularly characterized in advanced tumors if targeted therapies are considered.
Another interesting observation in our study is that a relatively large percentage of tumors showed loss of E-cadherin expression not associated with CDH1 mutations. Using the present Next-Generation Sequencing panel, we observed in previous studies by our group11 and in our routine practice that the percentage of ILC with E-cadherin loss that carry CDH1 mutations was about 80%. In this series, only 45% of the tumors carried CDH1 mutations in their lobular component. Although some studies analyzing ILC have reported a similar frequency of CDH1 mutations in ILC, these studies did not select cases based on E-cadherin loss.18 Our results suggested that mechanisms other than CDH1 mutations are frequently implicated in E-cadherin loss in IDLC.
We investigated by FISH whether homozygous CDH1 loss could be an alternative mechanism to gene mutation. However, we did not detect this alteration in any IDLC without a CDH1 mutation, despite the loss of a CDH1 copy being a constant event in all 14 tumors analyzed. Interestingly, we detected chromosome 16 monosomy in 2 tumors with CDH1 mutations, suggesting loss of the chromosome carrying the wild type allele. In an additional case homozygous CDH1 loss was observed in the lobular component.
An alternative mechanism to structural CDH1 alterations for E-cadherin loss could be CDH1 promoter hypermethylation. This alteration, which was not studied in this series, has been reported in several studies.19,20 However, this mechanism has been recently questioned and discrepancies between studies have been attributed to differences in the techniques used.5
It has been suggested that some ILC without CDH1 alterations could develop because of alterations of catenin genes.18,21 In our panel, we analyzed the complete sequences of CTNNA1, CTNNB1, JUN, and CTNND1, the genes coding for α, β, and γ-catenin and p120, respectively. However, we did not detect any pathogenic alteration. Although we cannot exclude chromosome alterations involving these loci, our results appear to exclude catenin genetic alterations as important contributors to IDLC.
Limitations of our study included the relatively low number of cases analyzed and the number of genes analyzed. However, our results suggested that most IDLC derived from IBC-NST in which a population of cells lost E-cadherin and acquired a lobular phenotype. In addition, the frequency of CDH1 mutation in IDLC in this series (45%) appears to be lower than in conventional ILC, suggesting the implication of alternative mechanisms of E-cadherin loss. Moreover, because of the molecular heterogeneity between ductal and lobular areas, molecular characterization should be performed on both components if targeted therapies are considered.
REFERENCES 1. WHO Classification of Tumours [Editorial Board]. Organisation mondiale de la santé. Breast Tumours, 5th ed. Geneva: OMS; 2019. 2. McCart Reed AE, Kalinowski L, Simpson PT, et al. Invasive lobular carcinoma of the breast: the increasing importance of this special subtype. Breast Cancer Res. 2021;23:6. doi:10.1186/s13058-020-01384-6 3. Christgen M, Cserni G, Floris G, et al. Lobular breast cancer: histomorphology and different concepts of a special spectrum of tumors. Cancers. 2021;13:3695. doi:10.3390/cancers13153695 4. Metzger-Filho O, Ferreira AR, Jeselsohn R, et al. Mixed invasive ductal and lobular carcinoma of the breast: prognosis and the importance of histologic grade. The Oncologist. 2019;24:e441–e449. 5. Ciriello G, Gatza ML, Beck AH, et al. Comprehensive molecular portraits of invasive lobular breast cancer. Cell. 2015;163:506–519. 6. Tazaki E, Shishido‐Hara Y, Mizutani N, et al. Histopathological and clonal study of combined lobular and ductal carcinoma of the breast. Pathol Int. 2013;63:297–304. 7. McCart Reed AE, Kutasovic JR, Nones K, et al. Mixed ductal-lobular carcinomas: evidence for progression from ductal to lobular morphology: mixed ductal-lobular tumours constitute a clonal entity. J Pathol. 2018;244:460–468. 8. Wagner PL, Kitabayashi N, Chen Y-T, et al. Clonal relationship between closely approximated low-grade ductal and lobular lesions in the breast: a molecular study of 10 cases. Am J Clin Pathol. 2009;132:871–876. 9. Wolff AC, Hammond MEH, Allison KH, et al. Human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists Clinical Practice Guideline Focused Update. Arch Pathol Lab Med. 2018;142:1364–1382. 10. Cheang MCU, Chia SK, Voduc D, et al. Ki67 index, HER2 status, and prognosis of patients with Luminal B breast cancer. J Natl Cancer Inst. 2009;101:736–750. 11. Rosa-Rosa J, Caniego-Casas T, Leskela S, et al. High frequency of ERBB2 activating mutations in invasive lobular breast carcinoma with pleomorphic features. Cancers. 2019;11:74. doi:10.3390/cancers11010074 12. Rosa-Rosa JM, Caniego-Casas T, Leskela S, et al. Modified SureSelectQXT target enrichment protocol for illumina multiplexed sequencing of FFPE samples. Biol Proced Online. 2018;20:19. doi:10.1186/s12575-018-0084-7 13. NovoAlign | Novocraft. Available at: http://www.novocraft.com/products/novoalign/. Accessed March 31, 2022. 14. McLaren W, Gil L, Hunt SE, et al. The ensembl variant effect predictor. Genome Biol. 2016;17:122. doi:10.1186/s13059-016-0974-4 15. González-Martínez S, Pizarro D, Pérez-Mies B, et al. Differences in the molecular profile between primary breast carcinomas and their cutaneous metastases. Cancers. 2022;14:1151. doi:10.3390/cancers14051151 16. Christgen M, Kandt LD, Antonopoulos W, et al. Inter‐observer agreement for the histological diagnosis of invasive lobular breast carcinoma. J Pathol CR. 2022;8:191–205. 17. Condorelli R, Mosele F, Verret B, et al. Genomic alterations in breast cancer: level of evidence for actionability according to ESMO Scale for Clinical Actionability of molecular Targets (ESCAT). Ann Oncol. 2019;30:365–373. 18. McCart Reed AE, Foong S, Kutasovic JR, et al. The genomic landscape of lobular breast cancer. Cancers. 2021;13:1950. doi:10.3390/cancers13081950 19. Sarrió D, Moreno-Bueno G, Hardisson D, et al. Epigenetic and genetic alterations of APC and CDH1 genes in lobular breast cancer: relationships with abnormal E-cadherin and catenin expression and microsatellite instability: E-Cadherin, Catenins and APC. Int J Cancer. 2003;106:208–215. 20. Roessler J, Ammerpohl O, Gutwein J, et al. The CpG island methylator phenotype in breast cancer is associated with the lobular subtype. Epigenomics. 2015;7:187–199. 21. de Groot JS, Ratze MA, van Amersfoort M, et al. αE-catenin is a candidate tumor suppressor for the development of E-cadherin-expressing lobular-type breast cancer: αE-catenin is a candidate driver of invasive lobular carcinoma. J Pathol. 2018;245:456–467.
留言 (0)