
記住我




Oxidation-Reduction (Redox) reactions, including those involving reactive oxygen species (ROS), lie at the heart of nearly every physiologic stress response in illness and injury (1). These redox reactions reflect the overall oxidative stress of the system and involve the transfer of electrons between oxidants and reductants. The continuous measurement of these combined reactions in blood can be viewed as the ambient blood redox potential (RP). As RP is the balance of oxidants and reductants present, it is a measure of electron pressure in the system much in the same way as pH is the measure of proton pressure (balance of acid and base) in a system. A low RP (more negative) indicates a larger concentration of reductants in the system (gain of electrons and/or decrease in oxidation state) with a negative RP trend indicating ongoing systemic reduction (i.e., anti-oxidation). Conversely, a higher RP (more positive) indicates a larger concentration of oxidants (loss of electrons and/or increase in oxidation state), with a positive RP trend indicating accumulating oxidation.
Despite the seeming importance of the measure, RP is not widely understood and there are currently few means by which to make the measurement. As opposed to the measurement of pH, quantifying the ultimate balance of electron exchanges presents unique challenges due to biofouling and other factors that affect electrode performance, especially when testing RP in complex biologic fluids such as blood. Because of these challenges, little is known about the changes in RP during dynamic states of critical illness and injury yet understanding the changing redox state could provide vital clinical data. Various redox pairs such as lactate-pyruvate, and oxidative stress indicators such a thiobarbituric acid reactive substances (TBARS) have been studied in an attempt to measure oxidative stress as an indicator of RP (2). Yet, such individual markers do not reflect the system as a whole since there are a large number of redox pairs that contribute to overall ambient RP similar to the many acid–base pairs that contribute to pH. In addition, these markers have limitations, including an inability to fully assess systemic redox state, as they can be affected by the origin of the sample measured (e.g., chemical vs enzymatic) and by the analysis techniques utilized for the measurement (3).
In response to these challenges, and in an effort to make RP measurements more clinically feasible, our group has developed RP electrodes, previously described (4), that make RP measurements possible in whole blood and other biologic media at the bedside. Furthermore, we have begun evaluating changing RP states in clinically relevant scenarios, including a published study demonstrating that RP measurements reflect real-time changes in oxygen debt and oxygen extraction ratio in a swine traumatic shock model (5). While promising, we seek to further characterize RP and its pathophysiologic role as a direct measure of oxidative stress in clinically relevant studies.
To this end, the purpose of this study was to investigate correlations between RP and time-matched changes in concentrations of circulating blood and excreted urine metabolites. Time series blood and urine samples were obtained throughout progressive shock and subsequent resuscitation in a swine traumatic shock model to identify metabolite-RP associations. While previous work by Clendenen et al. and others have described overarching metabolic changes in swine models of both hemorrhagic and traumatic shock (6), we focused more specifically on redox homeostasis and oxidative stress and hypothesized that whole blood RP measurements correlate with changing concentrations of specific metabolites over time, and that the metabolites identified would be attributable to pathways involved in oxidative stress. In addition, we hypothesized that urinary RP would correlate with time-matched concentrations of redox-related urine metabolites, reflecting the presence of ongoing changes in systemic and renal oxidative stress due to progressive traumatic/hemorrhagic shock.
MATERIALS AND METHODSThis study was designed and performed in accordance with the principles outlined in the eighth edition of the Guide for the Care and Use of Laboratory Animals (7) and was approved by the University of Michigan's Institutional Animal Care and Use Committee (PRO00006553 – “Testing and Comparison of Resuscitative Strategies in a Swine Model of Traumatic Shock”).
Swine traumatic shock modelDetails of this swine model of traumatic shock have been previously described (5). This model of controlled hemorrhagic shock and tissue trauma is performed targeting oxygen debt as a primary endpoint for hemorrhage. Oxygen debt, in mL/kg, was obtained by calculating the sum of continuous measures of oxygen deficit obtained at a sampling rate of 200 time points/s throughout the experiment using indirect calorimetry. Oxygen deficit (mL/kg/min) was calculated as the difference between baseline and continuous oxygen consumption (VO2), after an initial drop in end-tidal partial pressure of carbon dioxide (PetCO2) of 4 mm Hg. This drop in PetCO2 was used as a criterion for critical oxygen delivery (DO2) to minimize variability around critical DO2 and prevent any overestimation of oxygen debt.
Animals were anesthetized, intubated, and ventilated to a PetCO2 between and 35 and 45 mm Hg. Prior to hemorrhage, animals underwent soft tissue injury of the bilateral hind-limb skeletal muscle as well as comminuted femur fracture using captive bolt discharge. Animals were then hemorrhaged by removing 45% to60% of their blood volume while maintaining a mean arterial pressure (MAP) of 30 to 35 mm Hg, ultimately targeting an oxygen debt of 80 mL/kg. Once 80 mL/kg of oxygen debt was achieved, animals were resuscitated with approximately 15 mL/kg of whole blood (shed blood) and, if needed, 20 mL/kg of normal saline to restore MAP to > 60 mm Hg. Animals received an FiO2 of 0.21 during both hemorrhage and resuscitation. Mixed venous (MV) blood samples from the pulmonary artery (PA) were collected into sodium heparin tubes (BD Vacutainer®, Franklin Lakes, NJ) at baseline, max O2 Debt (80/mL/kg), immediately post resuscitation, and 2 h postresuscitation. Urine samples were also obtained concurrently at these time points via foley catheter that was placed immediately prior to experimentation. Continuous data, including arterial blood pressure, central venous blood pressure, and ECG were recorded using the BIOPAC MP150 Data Acquisition system (BIOPAC Inc, Goleta, CA).
RP Data/sample collectionRP measurements were obtained immediately after sample collection utilizing nanoporous gold electrodes previously described (4). Approximately 4 mL of MV whole blood was obtained at each time point from the PA during the experiment and immediately collected into sodium heparin tubes for testing. Given the excellent inter-rater reliability of the nanoporous gold electrodes for measuring RP in whole blood in a similar model of swine traumatic shock previously reported (5), one aliquot of approximately 1 mL of MV blood was taken and immediately used for direct RP testing at the time of sample collection. In addition, 1 mL of urine was collected at each time point for RP testing as well. Direct measurement of RP was performed on all samples by placing the whole blood or urine sample in a glass cell containing both the nanoporous gold electrode (working electrode) and an Ag/AgCl reference electrode, and then measuring open circuit potential (OCP) of the blood or urine sample using a ParstatMC™ multi-potentiostat (Princeton Applied Research). All RP measurements were made in the large animal lab at the point-of-care using the electrodes and technique described, taking approximately 10 min on average for the RP value to result. RP measurements for blood and urine were then recorded; RP was not adjusted for pH.
Whole blood and urine sample collection and processing for metabolomicsWhole blood MV samples were collected from the PA catheter into sodium heparin blood collection tubes (BD Vacutainer®, Franklin Lakes, NJ), gently inverted, and immediately placed in an ice water bath. Within 15 min of collection, samples were aliquoted into sterile, screw top cryogenic tubes and immediately flash frozen and stored in liquid nitrogen. Prior to the day of assay, samples were transferred from liquid nitrogen and stored overnight (−80°C). Samples were then thawed on ice and subjected to a methanol-chloroform extraction as previously described to remove macromolecules (8). After lyophilization, samples were resuspended in 50 mM sodium phosphate buffer in D2O. Following the addition of an internal standard, DSS-d6 (4,4-dimethyl-4-silapentane-1-sulfonic acid), pH was measured and adjusted to 6.5 to 7.5 by dropwise addition of 0.1 mM deuterium chloride or sodium deuteroxide after which each sample was transferred to NMR tubes (5 mm, 528-PP-7; Wilmad Lab Glass; Vineland, NJ) for further analysis by 1H-NMR.
At each timepoint, urine was collected from a Foley catheter into a urine collection cup (BD Vacutainer®) and collection volume was recorded. Samples were immediately chilled at 4°C and protected from light and then transferred into a 15 mL conical tube (BD Vacutainer®) with added NaN3 within 3 h. After centrifugation (2,000 × g for 10 min at 4°C), the supernatant was carefully transferred into a separate 15 mL conical tube before being aliquoted into 2 mL screw top cryotubes which were subsequently stored at −80°C (9). On the day of assay, samples were thawed at room temperature and an internal standard (Chenomx internal standard (IS)-2; deuterated 4,4-dimethyl-4-silapentane-1-sulfonic acid (DSS-d6) and NaN3 in D2O, pH 7.0) totaling 10% of sample volume was added. Each sample pH was adjusted to pH 7.0 ± 0.5 using sodium deuteroxide and/or deuterium chloride, as necessary, before being placed into clean glass NMR tubes (Wilmad) for further analysis.
Metabolomic analysis by 1H-NMRSpectra for each whole blood and urine sample were acquired on a Varian (now Agilent, Inc, Santa Clara, CA) 11.74 Tesla (500 MHz) NMR spectrometer with a VNMRS console operated by host software VNMRJ 4.0 and equipped with a 5-mm Agilent “One-probe.” Thirty-two scans of the first increment of a 1 H,1 H-NOESY (commonly referred to as a 1D-NOESY or METNOESY) pulse sequence were used to create the NMR spectra as previously described (8). All spectra were completed at room temperature (295.45 ± 0.3 K) at the University of Michigan's Biochemical NMR Core Laboratory. The resulting spectra were analyzed using the Chenomx NMR Suite 8.0 (Chenomx, Inc.). The Processor Module of the software was utilized for phase shift and baseline correction, while the Profiler Module allowed for the identification and quantification of metabolites against an internal library of 312 compounds.
Statistical analysisAll statistical analyses were performed using Python 3.7.3. MV blood and urine RP were independently compared with time-matched metabolomic profiles obtained from each source over the protocol period across all swine studied. The response and predictor variable pairs were matched according to their sources (i.e., mixed venous RP measurements were compared to the mixed venous metabolite profile, and urine RP to the urine metabolite profile). Metabolite concentrations were reported for all mixed venous samples. However, there were some metabolite and RP measures not recorded in urine due to the swine becoming anuric during that time period. If this was the case, these metabolites were removed for the analysis. Furthermore, metabolites with >80% missingness across an animal studied were removed. The remaining missing values were imputed with half the minimum value of the metabolite analyzed rather than imputing them to zero, consistent with previously published data (10).
The metabolomic data were z-scaled to an N (0,1) distribution. Models were constructed for each source using LASSO regression (scikit-learn 0.20.3, α = 1), which uses L1 regularization to shrink regression coefficients to zero and functionally perform variable selection. LASSO models were constructed using leave-one-out cross validation, where the models were trained on 6 of the animals and tested on the 7th. To be selected as candidates in the subsequent analysis, metabolites must maintain a non-zero coefficient with consistent magnitude and direction across at least 6 of the 7 cross validations. As L1 regularization pushes many of the coefficients to zero, effectively removing them from the model, metabolites that remain across many cross validations indicate that their relationship to RP is consistent across each of the animals. Metabolites meeting these criteria were considered as meaningful candidates and were subjected to pathway mapping. To link metabolites to metabolic pathways, KEGG IDs of these metabolites were uploaded into Metscape (http://metscape.ncibi.org/).
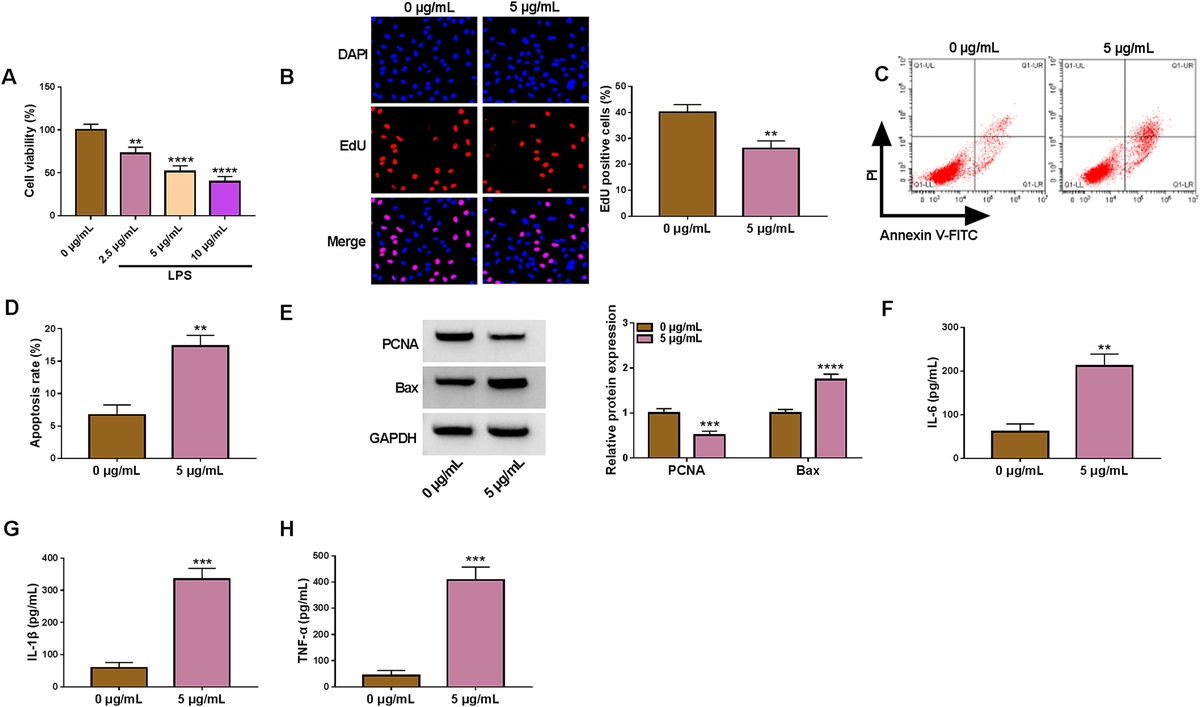
RESULTSA total of 34 metabolites in MV blood and 25 metabolites in urine specimens were detected and quantified (see Table 2a and b in supplemental file, https://links.lww.com/SHK/B450). Key relationships between RP and metabolites were found in both MV blood and urine samples across all animals studied. Coefficients from the LASSO regression across multiple cross validations show 23 metabolites that maintained a correlation with RP measures in MV blood and 20 in urine throughout the duration of the experiment in at least one swine (Figs. 1 and 2). As evidenced by the heat maps, LASSO regression identified the metabolites that maintained the magnitude and direction of correlation (whether positive or negative) across at least swine (Table 1A and B).
 Fig. 1:
Fig. 1: Heat map of all mixed venous (MV) metabolites that were found to have meaningful correlation with redox potential (RP) in at least one animal via LASSO regression throughout the swine traumatic shock model. Metabolites must have correlated with RP measures throughout the model and maintained magnitude and direction of coefficients across 6 or more swine (numbered 1 through 7 above) to be considered meaningful in our study. Right-sided Y-axis represents the range of correlation coefficients.
 Fig. 2:
Fig. 2: Heat map of all urine metabolites that were found to have a meaningful correlation with redox potential (RP) in at least one animal via LASSO regression throughout the swine traumatic shock model. Metabolites must have correlated with RP measures throughout the model and maintained magnitude and direction of coefficients across 6 or more swine (numbered 1 through 7 above) to be considered meaningful in our study. Right-sided Y-axis represents the range of correlation coefficients.
Table 1 - (A) MV Metabolites and (B) Urine Metabolites and their associated correlation coefficients with standard deviation (SD) Metabolite Coefficient Choline −6.27 (4.41) ATP −4.39 (1.48) Glycine 5.93 (2.35) ADP 1.84 (1.47) Glucose 15.96 (3.62) Formate −13.09 (4.83) Pyruvate 6.18 (1.56) Taurine −7.18 (3.01) Metabolite Coefficient Betaine 4.81 (2.41) Urea 4.14 (0.95) Glycine −2.97 (1.85) Taurine 10.32 (3.03) 3-Hydroxyisobutyrate −7.67 (3.41) N-phenylacetylglycine −14.52 (4.34) Hippurate 12.89 (3.76) Formate −5.89 (2.67)Each were found to have meaningful relationships with RP measures in at least 6 or more swine while maintaining magnitude and direction of coefficients.
In MV blood, a total of 8 metabolites met these criteria, with 4 metabolites increasing in concentration (positive coefficient) when RP measures increased, while 4 had a decreasing concentration (negative coefficient) when RP increased. Metabolites with positive coefficients were (coefficient [standard deviation]): ADP (1.84 [1.47]), glucose (15.96 [3.62]), glycine (5.93 [2.35]), and pyruvate (6.18 [1.56]). Metabolites with a decreasing concentration (negative coefficient) were: ATP (−4.39 [1.48]), choline (−6.27 [4.41]), formate (−13.09 [4.83]), and taurine (−7.18 [3.01]).
In urine, a total of 8 metabolites maintained magnitude and direction of correlation in 6 or more swine. Those with increasing concentration when RP measures increased were: betaine (4.81 [2.41]), urea (4.14 [0.95]), taurine (10.32 [3.03]), and hippurate (12.89 [3.76]). Those with decreasing concentration as RP increased were: glycine (−2.97 [1.85]), 3-hydroxyisobutyrate (−7.67 [3.41]), N-phenylacetylglycine (−14.52 [4.34]), and formate (−5.89 [2.67]).
Examination of the biochemical function of these metabolites revealed that the vast majority play a role in systemic responses to oxidative stress and/or reflect ongoing oxidative stress both systemically and locally (i.e., within the kidney). Pathway mapping of the metabolites generated two networks in which the glycolysis and gluconeogenesis pathway were common to both (Fig. 3).
 Fig. 3: Metabolic relationships among metabolites found to have meaningful correlations with RP in mixed venous blood. The metabolic networks of the MV LASSO regression model. The metabolites glucose (KEGG ID: C00221), pyruvate (KEGG ID: C00022), choline (KEGG ID: C00114), ADP (KEGG ID: C00008), ATP (KEGG ID: C00002), glycine (KEGG ID: C00037), formate (KEGG ID:C00058), and taurine (KEGG ID: C00245) were uploaded into Metscape (http://metscape.ncibi.org/). The compound-reaction-enzyme-gene function resulted in the generation of two networks in which the glycolysis and gluconeogenesis pathway was common to both. Because the glycolysis and gluconeogenesis pathway were present in both networks, three glucose metabolism by-products, 3-phospho-D-glyceroyl phosphate (KEGG ID: C00236), beta-D-fructose-6-phosphate (KEGG ID: C05345) and 2-phospho-D-glycerate (KEGG ID: C00631), designated as dark blue compounds, were added to the data set. This resulted in the connection of the two networks. Within this parent network (center), glucose, pyruvate, ADP, and ATP contribute to (A) the glycolysis and gluconeogenesis subnetwork and choline contributes to (B) the glycerophospholipid metabolism pathway. Glycine, serine, alanine, and threonine metabolism is represented by three sub-networks (C1-3) that include choline and pyruvate. Pyruvate also contributes to methionine and cysteine metabolism (D); ATP and ADP participate in purine metabolism (E); and glycine is central to lysine metabolism (F), porphyrin metabolism (G) and leukotriene metabolism (H). Formate is part of biopterin metabolism (I) and both glycine and taurine are involved in bile acid synthesis (J). Taurine also contributes to the methionine and cysteine metabolism subnetwork (K). Additional subnetworks are more broadly distributed in the parent network and are shown in Supplemental file Figure 4, https://links.lww.com/SHK/B450. In the illustration, the subnetworks are not spatially oriented. Compounds are represented by red hexagons (labeled metabolites are those that were uploaded into Metscape; the dark blue metabolites are those added to connect the initially generated two networks), reactions by grey squares, enzymes by green round corner squares and genes by blue circles.DISCUSSION
Fig. 3: Metabolic relationships among metabolites found to have meaningful correlations with RP in mixed venous blood. The metabolic networks of the MV LASSO regression model. The metabolites glucose (KEGG ID: C00221), pyruvate (KEGG ID: C00022), choline (KEGG ID: C00114), ADP (KEGG ID: C00008), ATP (KEGG ID: C00002), glycine (KEGG ID: C00037), formate (KEGG ID:C00058), and taurine (KEGG ID: C00245) were uploaded into Metscape (http://metscape.ncibi.org/). The compound-reaction-enzyme-gene function resulted in the generation of two networks in which the glycolysis and gluconeogenesis pathway was common to both. Because the glycolysis and gluconeogenesis pathway were present in both networks, three glucose metabolism by-products, 3-phospho-D-glyceroyl phosphate (KEGG ID: C00236), beta-D-fructose-6-phosphate (KEGG ID: C05345) and 2-phospho-D-glycerate (KEGG ID: C00631), designated as dark blue compounds, were added to the data set. This resulted in the connection of the two networks. Within this parent network (center), glucose, pyruvate, ADP, and ATP contribute to (A) the glycolysis and gluconeogenesis subnetwork and choline contributes to (B) the glycerophospholipid metabolism pathway. Glycine, serine, alanine, and threonine metabolism is represented by three sub-networks (C1-3) that include choline and pyruvate. Pyruvate also contributes to methionine and cysteine metabolism (D); ATP and ADP participate in purine metabolism (E); and glycine is central to lysine metabolism (F), porphyrin metabolism (G) and leukotriene metabolism (H). Formate is part of biopterin metabolism (I) and both glycine and taurine are involved in bile acid synthesis (J). Taurine also contributes to the methionine and cysteine metabolism subnetwork (K). Additional subnetworks are more broadly distributed in the parent network and are shown in Supplemental file Figure 4, https://links.lww.com/SHK/B450. In the illustration, the subnetworks are not spatially oriented. Compounds are represented by red hexagons (labeled metabolites are those that were uploaded into Metscape; the dark blue metabolites are those added to connect the initially generated two networks), reactions by grey squares, enzymes by green round corner squares and genes by blue circles.DISCUSSION
Redox reactions lie at the heart of physiologic processes in critical illness and injury. While metabolically active biologic systems rely on orderly conduction of electrons through interconnected redox reactions, alterations in redox balance (measured by redox potential, RP) have been linked to the generation of harmful free radicals (11), cellular apoptotic signaling (12), and the disruption of extracellular redox balance (11, 13), subsequently producing systemic redox shifts and oxidative stress. These redox shifts and oxidative stress also play a central role in driving the systemic inflammatory response in critical illness and injury and contribute to the development of multiple organ dysfunction (14, 15). In fact, mitochondrial ROS have been found to have the ability to drive the initiation of the inflammatory cascade via inflammasome-dependent and independent pathways, as well as through their interaction with Damage Associated- and Pathogen Associated-Molecular Pattern Molecules (DAMPS and PAMPS) (16). In addition, while restoring redox balance is of vital importance to improving the metabolic derangements in shock (17, 18), a strong association has also been demonstrated between individual markers of cellular redox balance and the onset of organ failure after injury, and these measurements are able to differentiate survivors from non-survivors during critical illness in one study (19). In the case of acute kidney injury (AKI), oxidative stress mediated pathways induce damage via reactive oxygen species (ROS) generation, nitric oxide depletion, DAMP generation, and microvascular dysfunction. Endogenous antioxidants and regulatory mechanisms are subsequently overwhelmed to the point that physiological homeostasis is disrupted, further provoking AKI (20). As with systemic redox measurements, techniques allowing for more accurate assessment of oxidative stress could improve the treatment of AKI in critical care settings as well.
Therefore, establishing straightforward, real-time measurements of redox balance that could be performed at or near the point of care could enable clinicians to quantify changing oxidative stress as it occurs, greatly improving our understanding of these changes in critical illness and injury. Our findings indicate that RP may be able to fill this role, as we were not only able to identify meaningful relationships between RP and changing metabolite concentrations, but also that these metabolites are active in systemic and local (e.g., renal) metabolic responses to oxidative stress, often serving important metabolic roles in antioxidant defense as well. Certainly, metabolites that serve to ensure energy generation and conservation during periods of shock and oxidative stress are central to systemic responses generated in these conditions, and while multiple metabolic pathways are involved, the two most essential overarching pathways involve glucose metabolism and mitochondrial activity/metabolism. It is no surprise that both are strongly linked to redox dependent regulation and responses, and are also well represented in our results. In fact, pathway analysis of RP-correlated metabolites generated two primary network pathways involving glycolysis and gluconeogenesis. These pathways were joined into one large parent network when 3 glucose metabolism by-products were included (Fig. 3). Secondary pathways related to this parent network, also associated with the metabolites listed in Table 1A and B, include those involving amino acid metabolism, porphyrin metabolism, leukotriene metabolism, biopterin metabolism, and bile acid synthesis.
Each of these secondary pathways are known to influence the redox state and oxidative stress of the system, both in a pro-oxidant manner, increasing systemic oxidation as in the case of porphyrin metabolism, leukotriene metabolism, and bile acid synthesis (21, 22), and by counteracting circulating oxidants by acting in an antioxidant capacity as with biopterin metabolism (23). Additional secondary subgroups are noted in the supplement (Figure 4, https://links.lww.com/SHK/B450) and involve the urea cycle and folate metabolism. However, in the setting of changing redox environments directly measured via RP, each individual metabolite has significant activity and responses inherent to its own metabolically active profile, whether related to the sub-pathways noted above, and/or due to inherent mechanisms of action in states of oxidative stress.
MV Metabolite/RP relationshipsLooking more closely at individual metabolites with correlation to RP measures in MV blood (Table 1A), glucose is noted to have one of the highest correlation coefficients (15.96 [3.62]), with its concentration increasing during periods of higher/increasing RP measures (increasing oxidation). This is understandable, as glucose levels are known to increase in response to trauma, sepsis, and critical illness/injury (24). However, this is especially significant given that during periods of acute oxidative stress, there is impairment in β-cell insulin secretion and concurrent decreases in glucose metabolism; the resulting hyperglycemia increases the production of free radical species by circulating leukocytes and other cells (1, 25). Furthermore, the ongoing presence of hyperglycemia has been associated with hepatic oxidative stress, antioxidant depletion, and the activation of systemic inflammatory responses (26), all of which are consistent with our findings that in the presence of a more oxidized state and higher/increasing RP measures there are also increasing glucose concentrations.
Formate and taurine exhibit the next highest coefficients in our study, and both are noted to decrease with higher/increasing RP and an increasingly oxidized environment. While these metabolites are associated with many active biological pathways, their biochemical activity points to why their concentrations fall in more oxidized settings. Formate contributes to anti-oxidative defense by acting as a reducing force to generate NADH and NADPH, while also undergoing cyclization from GTP to dihydroneopterin in periods of oxidative stress, a precursor for biopterin metabolism which has antioxidant effects. This mechanism is noted in Figure 3, subsection I (27). Furthermore, formate is utilized to recycle dihydrobiopterin (BH2) back to tetrahydrobiopterin (BH4) in order to improve overall antioxidant capacity (28). This has been demonstrated in clinical scenarios of oxidative stress as well, specifically in patients with critical limb ischemia who were noted to have reduced BH4, increased BH2, and reduced BH4/BH2 ratios (29). In each of these circumstances, formate is consumed to counteract oxidized environments, which is consistent with our data. Taurine appears to have an antioxidant role as well, although few studies evaluate taurine at physiologic concentrations. Its activity appears to be dependent upon its transport into cells by the TauT transporter, which is increased in periods of systemic inflammation and oxidative stress in order to replenish intracellular taurine and help inhibit ROS generation. This would decrease circulating taurine available as RP measures increase, which is consistent with our data (30).
Pyruvate and glycine also participate in oxidative stress responses and have positive correlations with RP measures in our study (increasing with increased oxidation). Pyruvate can function within the cell, yet its activity in the extracellular environment allows for the detoxification of ROS that also results in activation of acetate kinase, which in turn helps to maintain energy production in an oxygen independent manner (31). Pyruvate provides remarkable peroxide scavenging in oxidized environments as well, directly chelating H2O2 when increased extracellular concentrations of pyruvate are available, as reflected in our data (32). Glycine is more complicated as it is involved in multiple metabolic sub-pathways as illustrated in Figure 3. These include forming leukotrienes, as well as combining with succinate to make porphyrins, and with taurine to produce bile acids, each of which has pro-oxidant effects. However, given that our traumatic shock model induces ischemia with subsequent reperfusion over a relatively short period of time, these effects are likely limited in scope in comparison to the antioxidant effects of free glycine and its scavenging of oxygen free radicals (33). There is evidence this is the case based on experimental models as well, where glycine was found to have lower concentrations during periods of ischemia, yet when infused prior to reperfusion was able to reduce ischemia-reperfusion injury and reduce the oxidative burden (34). Again, this activity is reflected in our data as glycine concentrations fall when RP decreases during hemorrhagic shock and ischemia, and then increases as RP rises with increases in circulating oxidants during resuscitation and reperfusion.
Choline has also been shown to exert antioxidant effects both directly and indirectly, including reducing hepatocellular ROS and responding to oxidative stress by affecting the expression of PERK/Nrf-2 signaling pathways that regulate antioxidant responses, subsequently decreasing ROS in experimental studies (35). Although choline concentrations decreased as RP measures (and oxidation) increased in our study, choline very well may be consumed in these processes, while also undergoing conversion to acetylcholine. In addition, choline is easily converted to betaine in oxidized environments, and the amount of betaine excreted in the urine of swine in our model was found to increase with increasing oxidation across all cross-validations (36).
Finally, ATP and ADP were correlated to RP measures as well, but in opposite directions, with ATP concentrations decreasing with increasing oxidation while ADP levels increased. This is understandable given that ATP is converted to ADP when utilized as an energy source, and that tissue ATP levels are known to decline following shock and ischemia, as well as during periods of increased oxidative stress and within oxidized environments (37). Additionally, ADP decreases the production of ROS in oxidized states, and ADP has been shown to aid in preserving mitochondrial function, maintaining energy production in ischemia-reperfusion injury and other states of severe oxidative stress (38).
Urine metabolite/RP relationshipsThere is much less available literature regarding urine metabolite concentrations in the setting of oxidative stress, however there are some relationships that warrant recognition. This is especially true regarding those urinary metabolites in our study that demonstrated an increase in concentration when urinary RP decreased, and then declined in concentration with increasing oxidation/RP measures. These metabolites were glycine, N-Phenylacetylglycine (PAG), 3-Hydroxybutyrate (3HB), and formate. Glycine has been noted to be renal protective, specifically in renal tubules, and glycine levels have been shown to fall in the setting of increased renal oxidative stress (39), which may reduce urinary concentrations of glycine in increasingly oxidized states. However, the combined presence of glycine and PAG in urine has also been found to correlate with the reduced-to-oxidized glutathione ratio (GSH:GSSG), a redox couple that is sometimes used to assess the general state of oxidation vs reduction in a sample in which a low GSH:GSSG ratio represents oxidation. In the study, increasing glycine-PAG levels were correlated to higher GSH:GSSG ratios and may be predictive of response to therapeutic intervention (40). In our study, glycine and PAG are noted to decrease with increasing urinary RP/oxidization, which is consistent with the reported findings. Little data exists regarding 3HB in urine, but it has been noted to contribute to protection against oxidative damage in general (41), and one study evaluating the concentration of urinary enantiomers of 3HB in progressive kidney injury noted significantly lower levels of 3HB as kidney injury progressed, consistent with our data (42). Urinary formate is not well studied with regard to renal oxidative stress, however one study noted decreasing concentrations of urinary formate in primates with diabetic nephropathy vs both controls and other primates with diabetes but no nephropathy, suggesting formate levels may fall in urine when kidney injury and oxidative stress is present, as was the case in our study (43).
The remaining urinary metabolites were found to increase as RP measures and urinary oxidation increased. Again, there is not extensive literature on the activity of some of these metabolites in urine during states of oxidative stress, however some associations have been described. Urinary hippurate levels have also been found to strongly correlate with plasma antioxidant biomarkers, consistent with our results as well (44). As noted above, choline is easily oxidized to betaine, and this process may be one reason we see increasing betaine levels in the urine with increasing systemic oxidation and blood RP levels. Betaine also can inhibit NF-kB and has been found to reduce hepatic ROS, oxidative stress, and proinflammatory cytokines (45). There is little literature regarding urinary urea concentrations and oxidative stress, however it is well known that the degree of urea excretion relates to the degree of acute kidney injury (AKI) and the extent of ischemia/reperfusion injury-induced metabolic acidosis that is present (46). In addition to the antioxidant activity of circulating taurine, taurine has also been shown to exhibit a protective role in the setting of kidney injury by decreasing oxidative stress and improving mitochondrial dysfunction in renal tissue (47). In our model, urinary taurine increases as RP values and renal oxidation increases, which may reflect this activity.
Although multiple metabolites were found to have consistent relationships with RP measures throughout our model, both in MV blood and in urine, collectively these metabolites form an integrated picture of metabolic activity functioning during periods of oxidative stress, and one that correlates with RP measures taken at the point of care. While all metabolites do not demonstrate the same direction of correlation (i.e., some metabolites increase during increasing oxidation while others decrease), their behavior throughout the model can be understood by highlighting their known metabolic activity, and through data presented in multiple studies exploring the biochemical relationships of these metabolites in states of ischemia/reperfusion and oxidative stress. These data demonstrate that RP measures may not only provide an assessment of systemic oxidative stress in real time, but also an evaluation of local, organ specific injury as reflected in the urinary metabolite/RP relationships noted in this study. Furthermore, RP measures may not only provide diagnostic information regarding the state of oxidative stress, but may also play a role in guiding therapeutic intervention. In this way, these measures may serve as a unique marker for the use of antioxidant and/or anti-reductant therapies as well. Therefore, the ability to monitor both systemic and local RP may aid in optimizing the use of therapies that impact multiple clinical scenarios, from traumatic shock to septic shock, to organ-specific monitoring and treatment that targets acute kidney injury.
LIMITATIONSThis is a preliminary study of the relationship between RP measurements and the activity of multiple metabolites in whole blood and urine using a novel nanoporous electrode that is resistant to biofouling. As such, we acknowledge that our sample size was limited to seven swine. While important relationships between RP and the metabolome were uncovered using a cross-validated LASSO approach, obtaining additional data in a larger cohort of swine, and ultimately in a patient population, would allow us to further evaluate metabolite-RP relationships with more power. Although we utilized a well-established model of swine shock with controlled hemorrhage and shock progression, further studies are needed to evaluate RP measurements in traumatic shock and in other shock states, to further investigate RP in these models. This is particularly true for an assessment of the utility of urine metabolites which are often normalized to creatinine values which has limitations in models of critical illness. Since supplemental oxygen was not used, which could have had some effect on RP measures made during the resuscitation phase, this may limit clinical translation of our findings. Lastly, more population-based data will need to be collected to understand ranges of RP in patients with various conditions and in response to various therapies.
CONCLUSIONSIn this swine model of traumatic shock, real-time RP measures demonstrate meaningful relationships with metabolites attributable to metabolic pathways involved in systemic responses to oxidative stress, as well as those active through these responses. These results support RP measures as a feasible, biologically relevant marker of oxidative stress both systemically and at the local/organ level. As a direct measure of redox state, RP may be a useful biomarker to understand the role of RP in health and disease.
AcknowledgmentsThe authors would like to acknowledge the NIH, the Pediatric Critical Care and Trauma Scientist Development Program (PCCTSDP), the Michigan Center for Integrative Research in Critical Care (MCIRCC), and the University of Michigan's Fast Forward Medical Innovation Program (FFMI) for their financial support of this work, as well as the ongoing support of the Department of Pediatrics, the Division of Pediatric Critical Care Medicine, the Lurie Nanofabrication Facility, and MCIRCC at the University of Michigan.
REFERENCES 1. Dodson M, Darley-Usmar V, Zhang J. Cellular metabolic and autophagic pathways: traffic control by redox signaling. Free Radic Biol Med 63:207–221, 2013. 2. Gyurászová M, Kovalčíková A, Janšáková K, Šebeková K, Celec P, Tóthová Ľ. Markers of oxidative stress and antioxidant status in the plasma, urine and saliva of healthy mice. Physiol Res 67:921–934, 2018. 3. Tsikas D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Anal Biochem 524:13–30, 2016. 4. Patel J, Radhakrishnan L, Zhao B, Uppalapati B, Daniels RC, Ward K, Collinson M. Electrochemical properties of nanostructured porous gold electrodes in biofouling solutions. Anal Chem 85 (23):11610–11618, 2013. 5. Daniels RC, Jun H, Tiba H, McCracken B, Herrera-Fierro P, Collinson M, Ward K. Whole blood redox potential correlates with progressive accumulation of oxygen debt and acts as a marker of resuscitation in a swine hemorrhagic shock model. Shock 49 (3):345–351, 2018. 6. Clendenen N, Nunns GR, Moore EE, Reisz JA, Gonzalez E, Peltz E, Silliman CC, Fragoso M, Nemkov T, Wither M, et al. Hemorrhagic shock and tissue injury drive distinct plasma metabolome derangements in swine. J Trauma Acute Care Surg 83 (4):635–642, 2017. 7. National Research Council (U.S.). Committee for the Update of the Guide for the Care and Use of Laboratory Animals, Institute for Laboratory Animal Research (U.S.), National Academies Press (U.S.). Guide for the care and use of laboratory animals. In: 8th ed. Washington, DC: National Academies Press; 2011. 8. McHugh CE, Flott TL, Schooff CR, Smiley Z, Puskarich MA, Myers DD, Younger JG, Jones AE, Stringer KA. Rapid, reproducible, quantifiable NMR metabolomics: methanol and methanol: chloroform precipitation for removal of macromolecules in serum and whole blood. Metabolites 8 (4):93, 2018. 9. Emwas AH, Luchinat C, Turano P, Tenori L, Roy R, Salek RM, Ryan D, Merzaban JS, Kaddurah-Daouk R, Zeri AC, et al. Standardizing the experimental conditions for using urine in NMR-based metabolomic studies with a particular focus on diagnostic studies: a review. Metabolomics 11 (4):872–894, 2015. 10. Bijlsma S, Bobeldijk I, Verheij ER, Ramaker R, Kochhar S, Macdonald IA, van Ommen B, Smilde AK. Large-scale human metabolomics studies: a strategy for data (pre-) processing and validation. Anal Chem 78 (2):567–574, 2006. 11. Andrades ME, Morina A, Spasic S, Spasojevic I. Bench-to-bedside review: sepsis—from the redox point of view. Crit Care 15 (5):230, 2011. 12. Iijima T, Mishima T, Akagawa K, Iwao Y. Mitochondrial hyperpolarization after transient oxygen-glucose deprivation and subsequent apoptosis in cultured rat hippocampal neurons. Brain Res 993:140–145, 2003. 13. Merker MP, Bongard RD, Kettenhofen NJ, Okamoto Y, Dawson CA. Intracellular redox status affects transplasma membrane electron transport in pulmonary arterial endothelial cells. Am J Physiol Lung Cell Mol Physiol 282:L36–43, 2002. 14. Biesalski HK, McGregor GP. Antioxidant therapy in critical care—is the microcirculation the primary target? Crit Care Med 35: (9 Suppl): S577–S583, 2007. 15. Motoyama T, Okamoto K, Kukita I, Hamaguchi M, Kinoshita Y, Ogawa H. Possible role of increased oxidant stress in multiple organ failure after systemic inflammatory response syndrome. Crit Care Med 31:1048–1052, 2003. 16. Escobar J, Pereda J, Lopez-Rodas G, Sastre J. Redox signaling and histone acetylation in acute pancreatitis. Free Radic Biol Med 52:819–837, 2012. 17. Alonso de Vega JM, Diaz J, Serrano E, Carbonell LF. Oxidative stress in critically ill patients with systemic inflammatory response syndrome. Crit Care Med 30 (8):1782–1786, 2002. 18. Cowley HC, Bacon PJ, Goode HF, Webster NR, Jones JG, Menon DK. Plasma antioxidant potential in severe sepsis: a comparison of survivors and nonsurvivors. Crit Care Med 24 (7):1179–1183, 1996. 19. Ware LB, JP F, May AK, Roberts LJ. Plasma biomarkers of oxidant stress and development of organ failure in severe sepsis. Shock 36:12–17, 2011. 20. Pavlakou P, Liakopoulos V, Eleftheriadis T, Mitsis M, Dounousi E. Oxidative stress and acute kidney injury in critical illness: pathophysiologic mechanisms—biomarkers—interventions, and future perspectives. Oxid Med Cell Longev 2017:1–11, 2017. 21. Bomzon A, Holt S, Moore K. Bile acids, oxidative stress, and renal function in biliary obstruction. Semin Nephrol 17 (6):549–562, 1997. 22. Dvash E, Har-Tal M, Barak S, Meir O, Rubinstein M. Leukotriene C4 is the major trigger of stress-induced oxidative DNA damage. Nat Commun 6 (1):10112, 2015. 23. Rebelo J, Auerbach G, Bader G, Bracher A, Nar H, Hosl C, Schramek N, Kaiser J, Bacher A, Huber R, et al. Biosynthesis of pteridines. Reaction mechanism of GTP cyclohydrolase I. J Mol Biol 326 (2):503–516, 2003. 24. Sperry JL, Frankel HL, Nathens AB, O’Keefe GE, Cuschieri J, Moore EE, Maier R, Minei JP. Characterization of persistent hyperglycemia: what does it mean postinjury? J Trauma 66 (4):1076–1082, 2009. 25. Mohanty P, Hamouda W, Garg R, Aljada A, Ghanim H, Dandona P. Glucose challenge stimulates reactive oxygen species (ROS) generation by leucocytes. J Clin Endocrinol Metab 85 (8):2970–2973, 2000. 26. Ling PR, Smith RJ, Bistrian BR. Acute effects of hyperglycemia and hyperinsulinemia on hepatic oxidative stress and the systemic inflammatory response in rats. Crit Care Med 35 (2):555–560, 2007. 27. Thomas SC, Alhasawi A, Auger C, Omri A, Appanna VD. The role of formate in combatting oxidative stress. Antonie Van Leeuwenhoek 109 (2):263–271, 2016. 28. Crabtree MJ, Channon KM. Synthesis and recycling of tetrahydrobiopterin in endothelial function and vascular disease. Nitric Oxide 25 (2):81–88, 2011. 29. Ismaeel A, Papoutsi E, Miserlis D, Lavado R, Haynatzki G, Casale GP, Bohannon WT, Smith RS, Eidson JL, Brumberg R, et al. The nitric oxide system in peripheral artery disease: connection with oxidative stress and biopterins. Antioxidants 9 (7):590, 2020.
留言 (0)