
記住我


Alzheimer's disease (AD) is a progressive neurodegenerative disease that accounts for about 80% of dementia cases [1]. Pathological hallmarks of AD include the formation of extracellular amyloid β (Aβ) protein plaques and intracellular neurofibrillary tangles (NFT) composed of hyperphosphorylated tau (pTau) protein [2]. The deposition of Aβ plaques is an early event in AD pathogenesis, which induces subsequent inflammation, accumulation of NFTs, and ultimately neuronal dysfunction and cell death [3]. Genetic risk factors for AD development are principally involved in Aβ processing and clearance – most notably apolipoprotein ε4 allele (APOE4) – and are expressed by CNS-resident immune cells [4]. Additionally, an estimated 21%–42% of lifetime risk on AD is attributed to environmental factors [5]. Recent studies demonstrated that Aβ can function as an antimicrobial peptide that protects neurons from bacterial and viral infections [6, 7], supporting the hypothesis that microbial infections – especially herpes simplex virus 1 (HSV-1) – could play a role in the pathogenesis of AD.
Most adults worldwide are infected with the human neurotropic alphaherpesvirus herpes simplex virus 1 (HSV-1). Latent HSV-1 predominantly resides in the somata of pseudounipolar sensory trigeminal ganglion (TG) neurons, which innervate both the orofacial epithelia and the brainstem [8]. Periodic HSV-1 reactivation results in virus spread to the oral mucosa and the central nervous system (CNS). Whereas oral HSV-1 shedding can be both asymptomatic and symptomatic (cold sores), symptomatic HSV-1 infection of the CNS is extremely rare and associated with severe morbidity and mortality (herpes simplex encephalitis; HSE) [9, 10]. Multiple studies have detected HSV-1 DNA in the brain of elderly individuals with and without AD [11-15]. In the brain of AD patients, viral DNA was found to preferentially colocalize with Aβ plaques [16]. The presence of HSV-1 in brain could contribute to the pathogenesis of AD by inducing the accumulation of intracellular Aβ, extracellular Aβ aggregates, and nuclear accumulation and hyperphosphorylation of tau protein, as observed in cultures of mouse primary cortical neurons, human neuroblastoma cells, and human iPSC-derived neuron models [17-24]. In mice, these AD-related pathological changes were found to progressively accumulate with repeated HSV-1 infection and to correlate with cognitive impairment [25]. However, humans are the only natural host of HSV-1. Unlike mice, lytic HSV-1 infection of the human CNS is extremely rare, in part, because human cortical neurons are intrinsically resistant to HSV-1 infection [26]. It remains to be determined whether lytic or latent HSV-1 infection of human neurons in vivo is directly associated with aberrant Aβ and pTau expression. Therefore, the aim of this study was to investigate the relationship between AD-related neuropathological changes and HSV infection in the TG of latently HSV-1 infected individuals and the brain of individuals with lytic HSV infection of the brain (HSE).
2 METHODS 2.1 Human clinical specimensTemporal cortex samples from AD patients and paired human TG and plasma samples from healthy controls, AD patients, and patients with other neurological diseases were obtained from the Netherlands Brain Bank (Netherlands Institute for Neuroscience; Amsterdam, the Netherlands). All donors had provided written informed consent for brain autopsy and the use of material and clinical information for research purposes. All study procedures were performed in compliance with relevant laws in the Netherlands and in accordance with the ethical standards of the Declaration of Helsinki. Institutional guidelines were approved by the local ethical committee (VU University Medical Center, Amsterdam, project number 2009/148). AD brain tissues and some of the TG samples were fixed in neutral buffered formalin and embedded in paraffin (FFPE) for histological analysis. Additional TG samples were used to generate single-cell suspensions or snap-frozen for RNA extraction [27]. FFPE brain samples from patients with acute traumatic brain injury (TBI), HSE patients, and a VZV encephalitis patient were obtained for diagnostic purposes and provided by the BioBanks of the Erasmus MC (TBI patients) and Amsterdam University Medical Center (HSE and VZV encephalitis). According to the institutional “Opt-Out” system, which is defined by the National “Code of Good Conduct” [Dutch: Code Goed Gebruik, May 2011], these surplus human brain tissues were available for the current study.
2.2 Nucleic acid extraction, quantitative TaqMan real-time PCR (qPCR), and APOE genotypingOne-tenth of a TG single-cell suspension was used for DNA extraction using the QIAamp DNA Kit (Qiagen). RNA was extracted from homogenized TG using TRIzol (Thermo Fisher Scientific) and the RNeasy Mini Kit including on-column DNase I treatment (Qiagen), as described [28]. cDNA synthesis was performed using 5 µg total RNA and superscript IV with oligo(dT)12–18 primers (Thermo Fisher Scientific). TaqMan qPCR was performed in duplicate on the 7500 Real-Time PCR system using 2X PCR Universal Master Mix (Applied Biosystems) and primer/probe pairs specific for HSV-1 US4, HSV-1 LAT, VZV ORF38, and the human single-copy gene hydroxymethylbilane synthase [28, 29] (Table S1). APOE genotyping was performed using allele ε2-, ε3-, and ε4-specific primer/probe combinations and TaqMan qPCR, as described [30] (Table S1). Results were confirmed by PCR amplification of the APOE gene region containing allele-differentiating SNPs rs429358 and rs7412 using PfuUltra II Fusion High-Fidelity DNA Polymerase (Agilent), followed by Sanger sequencing using the BigDye 3.1 Cycle Sequencing Kit on the 3130XL Genetic Analyzer (Applied Biosystems) [30].
2.3 Immunohistochemical and immunofluorescent stainingFFPE tissue sections were subjected to heat-induced antigen retrieval using citrate buffer or Trilogy™ (Cell Marque). Immunohistochemical (IHC) staining was performed using the following primary antibodies: monoclonal mouse anti-β-Amyloid (clone 4G8, BioLegend), anti-phosphorylated-Tau (Ser202/Thr205) (AT8, Invitrogen), and anti-HSV-1 infected cell protein 8 (ICP8) (10A3, Cell Marque). Staining was visualized using biotinylated polyclonal rabbit anti-mouse Ig (Dako) and goat anti-rabbit Ig (Dako), followed by horseradish peroxidase-conjugated streptavidin (Dako) and 3-amino-9-ethylcarbazole substrate. Images were obtained with an Olympus BX51 microscope or by scanning the slides using the Hamamatsu NanoZoomer 2.0 HT.
Immunofluorescent (IF) staining was performed using the following primary antibodies: polyclonal rabbit anti-HSV-1 (cross-reactive with HSV-2; Agilent), anti-Iba1 (Wako), or anti-GFAP (Dako), polyclonal chicken anti-GFAP (Abcam), monoclonal mouse anti-NeuN IgG1 (A60, Sigma-Aldrich), anti-MBP (1.B.645, Santa Cruz Biotechnology), anti-Iba1 (GT10312, Invitrogen), anti-β-Amyloid (4G8, BioLegend), anti-phosphorylated-Tau (Ser202/Thr205) (AT8, Invitrogen), anti-CD45 (2B11 + PD7/26, Dako), and monoclonal rat anti-CD3 (CD3-12, Abcam). Alexa Fluor® 488 (AF488)-, AF594-, and AF647-conjugated polyclonal goat anti-rabbit IgG, anti-mouse IgG1, and IgG2b, goat anti-chicken IgY and goat anti-rat IgG secondary antibodies (Invitrogen) were used. Nuclei were stained with Hoechst 33342 Solution and mounted with ProLong™ Diamond Antifade Mountant (Thermo Fisher Scientific). Images were obtained using a Zeiss LSM700 confocal microscope.
2.4 In situ hybridizationTissue sections were stained for HSV-1 and VZV RNA by in situ hybridization (ISH), using the RNAscope® 2.5 HD Kit-RED and probes HSV-1-LAT (Cat No. 315651) and VZV-Pool (Cat No. 400701) (Advanced Cell Diagnostics). Slides were counterstained with hematoxylin (Sigma) and mounted with EcoMount (Biocare Medical).
2.5 Statistical analysesAll statistical analyses were performed using GraphPad Prism 8.0.2 (GraphPad Software Inc).
3 RESULTS 3.1 HSV-1 DNA load and LAT expression in the TG of AD patients and controlsPrevious studies suggest that HSV-1 infection or reactivation, as measured by plasma HSV-1 IgG and IgM levels, could be a risk factor for AD development [31]. Clinical HSV-1 reactivation frequency correlates with both APOE4 carriage (humans) and latent viral DNA load (mice) [32-35]. To investigate whether latent HSV-1 DNA load was associated with AD development or APOE genotype, we performed qPCR on human TG that were infected with HSV-1 and obtained from AD patients and controls. Additionally, we performed qPCR to detect the closely related varicella-zoster virus (VZV), because most human TG is co-infected by HSV-1 and VZV [10]. HSV-1 and VZV DNA loads were similar between the TG obtained from AD patients and controls (Figure 1A). Further, APOE genotyping of all analyzed TG specimens demonstrated comparable HSV-1 and VZV DNA levels in APOE4 carriers and non-carriers (Figure 1B and Figure S1), despite the reported effect of APOE4 on promoting HSV neurovirulence [36, 37]. We performed RT-qPCR and ISH to quantify expression of the HSV-1 latency-associated transcripts (LAT) in human TG. Similar to HSV-1 DNA loads, the prevalence of LAT-positive neurons and abundance of LAT RNA were comparable between AD patients and controls (Figure 1C).

HSV-1 infection is not associated with aberrant Aβ or pTau expression in latently infected human trigeminal ganglia (TG) neurons. (A, B) HSV-1- and VZV-specific qPCR was performed on DNA extracted from the TG of Alzheimer's disease (AD) patients and control subjects, stratified on disease status (A; 20 controls and 10 AD patients) or APOE4 allele carrier status (B; HSV-1: 22 APOE4-negative and 8 APOE4-positive individuals; VZV: 24 APOE4-negative and 13 APOE4-positive individuals). Horizontal line: median. (C) Detection of LAT RNA by RT-qPCR (7 AD patients and 7 controls) and ISH (11 controls and 5 AD patients; 3 sections per donor analyzed) in human TG from AD patients and control subjects. (D) Sequential TG sections from AD (n = 4) and control (n = 6) subjects were stained for amyloid β protein, HSV-1 latency-associated transcript (LAT) RNA and phosphorylated Tau protein (pTau; Ser202/Thr205) by immunohistochemistry (IHC) and RNA in situ hybridization (ISH). Arrowheads indicate LAT-positive neurons and asterisks indicate lipofuscin granules. Scale bar: 50 µm
3.2 No expression of Aβ and pTau in latently HSV-infected TG neurons of AD patients and controlsMost individuals acquire HSV-1 infection in their childhood, followed by a lifelong latent infection and frequent (asymptomatic) virus reactivation [38, 39]. Given that HSV-1 replication leads to the accumulation of Aβ and increased pTau expression in cultured neurons and in the brains of HSV-infected mice [17, 20, 21, 25, 40], we analyzed whether latent HSV-1 infection was associated with Aβ or pTau expression in TG neurons of AD patients and controls (Table S2). Analysis of consecutive TG sections from HSV IgG seropositive (n = 3 AD; n = 5 controls) and HSV IgG seronegative (n = 2 AD) donors did not demonstrate Aβ, Aβ plaques, or pTau staining in latently HSV-1-infected TG neurons of either AD patients or controls (Figure 1D). Similarly, we did not observe aberrant Aβ or pTau staining in neurons not infected by HSV-1.
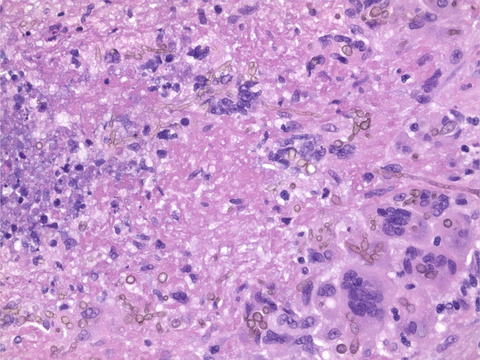
3.3 Expression of Aβ and pTau in brain of HSE patientsAs we did not observe aberrant Aβ or pTau expression in HSV-infected peripheral TG neurons, we hypothesized that HSV-induced AD-related pathological changes could be (1) restricted to CNS neurons, or (2) reversible and therefore limited to lytic (productive) virus infection. To test these hypotheses, we acquired rare post-mortem brain specimens from five HSE patients (Table 1), four cases of HSV-1 encephalitis and one case of neonatal HSV-2 encephalitis. Brain sections from all patients contained lytically HSV-infected cells, as evidenced by the detection of intranuclear eosinophilic (Cowdry type A) inclusion bodies, HSV-1 LAT RNA (in all HSV-1 HSE patients) and HSV ICP8 protein (Figure 2A). In all patients, the majority of HSV-infected ICP8POS cells were identified as NeuNPOS neurons, with lower frequencies of infected non-neuronal cells, mainly Iba1POS microglia (Figure 2B,C). Thus, brain sections from HSE patients provide a snapshot of the appearance of lytically HSV-infected CNS neurons in vivo.
TABLE 1. HSE patient characteristics and Alzheimer's disease-related pathology Patient Agea Genderb Causative virus Brain regions analyzed Aβ intraneuronalc Aβ plaques NFT 1 – – HSV-1 Cortex No No No 2 79 F HSV-1 Hippocampus/cortex Yes Yes Yes Amygdala/entorhinal Cortex Yes Yes Yes Cortex Yes Yes Yes Insula Yes Yes Yes 3 0d F HSV-2 Cortex No No No 4 81 F HSV-1 Putamen No Yes No 5 76 M HSV-1 Hippocampus/entorhinal cortex No Yes Yes Abbreviation: NFT, neurofibrillary tangle. Lytic HSV infection of neurons in the brain of herpes simplex virus encephalitis (HSE) patients. (A) Brain sections from HSE patients (Table 1) were stained with hematoxylin and eosin, or stained for HSV-1 LAT RNA by ISH and HSV-1 ICP8 protein by IHC. Boxes indicate the area shown at higher magnification in the inset. Scale bar: 50 µm. (B, C) Brain sections of HSE patients were immunofluorescently stained for (B) HSV-1 protein (green), neurons (NeuN; white) and oligodendrocytes (MBP; red), as well as (C) HSV-1 protein (green), microglia (Iba1; white), and astrocytes (GFAP; red). Nuclei were stained with Hoechst-33342 (blue). Representative images are shown for patient #2 (amygdala/entorhinal cortex; Table 1). Scale bar: 20 µm
Lytic HSV infection of neurons in the brain of herpes simplex virus encephalitis (HSE) patients. (A) Brain sections from HSE patients (Table 1) were stained with hematoxylin and eosin, or stained for HSV-1 LAT RNA by ISH and HSV-1 ICP8 protein by IHC. Boxes indicate the area shown at higher magnification in the inset. Scale bar: 50 µm. (B, C) Brain sections of HSE patients were immunofluorescently stained for (B) HSV-1 protein (green), neurons (NeuN; white) and oligodendrocytes (MBP; red), as well as (C) HSV-1 protein (green), microglia (Iba1; white), and astrocytes (GFAP; red). Nuclei were stained with Hoechst-33342 (blue). Representative images are shown for patient #2 (amygdala/entorhinal cortex; Table 1). Scale bar: 20 µm
We then investigated whether lytic HSV infection was associated with increased intracellular production of Aβ or pTau, or the deposition of extracellular Aβ plaques in brain sections from HSE patients (Table 1). As a control, we included brain sections from patients with acute traumatic brain injury (TBI, n = 3; Table S3) and AD patients (n = 3; Table S4). TBI-induced neuronal damage is associated with increased production of Aβ, resulting in its accumulation in axonal spheroids as well as plaques in about 30% of patients [41]. Indeed, we observed axonal spheroids in all three TBI patients and increased expression of intracellular Aβ protein and (diffuse) extracellular Aβ plaques in two TBI patients (Figure 3A). NFTs and pTau staining was not observed in TBI patients (Figure 3A). Abundant intracellular Aβ, extracellular Aβ plaques, and NFT were observed in all three AD patients (Figure 3A). By contrast, brain sections from two of five HSE patients showed neither Aβ protein/plaques nor pTau protein/NFTs (Table 1). Aβ plaques were observed in one of five HSE patients (patient #2), whereas intracellular accumulations of Aβ protein were detected in three of five HSE patients. NFTs were detected in two of five HSE patients (patients #2 and #5) (Figure 3A and Table 1).
 Lytic HSV infection is not consistently associated with aberrant Aβ or pTau expression in the brain of herpes simplex virus encephalitis (HSE) patients. (A) Brain sections from Alzheimer's disease (AD; patient 1, Table S4), trauma brain injury (TBI; patient 2, Table S3), and HSE patients (Table 1) were stained for Aβ and pTau (Ser202/Thr205) by IHC. Filled and open arrowheads indicate mature senile and diffuse Aβ plaques, respectively. Filled black arrows indicate intracellular Aβ accumulations. Open arrows indicate neurofibrillary tangles. Boxes indicate the area shown at higher magnification in the inset. Scale bar: 50 µm. (B) Consecutive brain sections from HSE patients were stained for Aβ and pTau (Ser202/Thr205) protein by IHC and HSV-1 RNA by ISH. Boxes indicate the area shown at a higher magnification. Scale bars indicate 500 µm (panels 1), 100 µm (panels 2), and 50 µm (panels 3)
Lytic HSV infection is not consistently associated with aberrant Aβ or pTau expression in the brain of herpes simplex virus encephalitis (HSE) patients. (A) Brain sections from Alzheimer's disease (AD; patient 1, Table S4), trauma brain injury (TBI; patient 2, Table S3), and HSE patients (Table 1) were stained for Aβ and pTau (Ser202/Thr205) by IHC. Filled and open arrowheads indicate mature senile and diffuse Aβ plaques, respectively. Filled black arrows indicate intracellular Aβ accumulations. Open arrows indicate neurofibrillary tangles. Boxes indicate the area shown at higher magnification in the inset. Scale bar: 50 µm. (B) Consecutive brain sections from HSE patients were stained for Aβ and pTau (Ser202/Thr205) protein by IHC and HSV-1 RNA by ISH. Boxes indicate the area shown at a higher magnification. Scale bars indicate 500 µm (panels 1), 100 µm (panels 2), and 50 µm (panels 3)
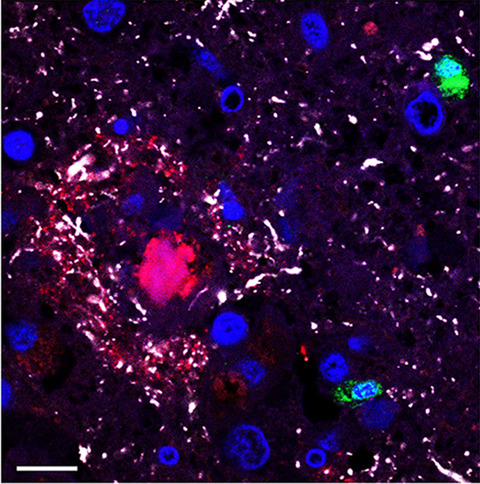
Subsequently, we determined the spatial relationship between HSV-infected cells and AD-related pathological changes. Although HSV-infected cells were occasionally observed in close proximity to NFT and/or Aβ plaques, majority of these AD-related pathological changes were not associated with sites of HSV replication in the brains of HSE patients #2 and #5 (Figure 3B). Similarly, intracellular accumulations of Aβ protein were widely spread through brain sections of HSV-1 HSE patients and not restricted to areas with HSV-1 infection (Figure 3B and Figure S2). IF staining was performed to investigate potential colocalization between HSV-1 infected cells and plaques/NFT and to assess the intracellular expression of Aβ and pTau in more detail. Again, occasional HSV-infected cells were observed adjacent to Aβ plaques and NFT, but HSV antigen never colocalized with Aβ plaques nor NFT (Figure 4A). In HSE patients #2, #4, and #5, abundant intracellular Aβ protein staining was detected not only in most HSV-infected neurons but also in non-infected neurons (Figure 4B). Nuclear pTau staining was observed in some HSV-infected cells in HSE patients containing NFTs (Figure 4C). However, we did not observe increased Aβ or pTau expression in HSV-infected neurons in HSE patients #1 and #3 (Figure 4D).
 HSV-infected cells do not consistently express aberrant Aβ or pTau in the brain of herpes simplex encephalitis (HSE) patients. Brain sections from HSE patients were immunofluorescently stained for HSV protein (green), Aβ (red), and pTau (Ser202/Thr205; white). Nuclei were stained with Hoechst-33342 (blue). Images are shown for HSE patients #2 (A left panel), #5 (A right panel, and C), #4 (B), and #3 (D) (Table 1). White boxes indicate the areas shown at higher magnification. Scale bars indicate 10 µm (A: Amyloid β plaques; and C) and 20 µm (A right panel, B and D)
HSV-infected cells do not consistently express aberrant Aβ or pTau in the brain of herpes simplex encephalitis (HSE) patients. Brain sections from HSE patients were immunofluorescently stained for HSV protein (green), Aβ (red), and pTau (Ser202/Thr205; white). Nuclei were stained with Hoechst-33342 (blue). Images are shown for HSE patients #2 (A left panel), #5 (A right panel, and C), #4 (B), and #3 (D) (Table 1). White boxes indicate the areas shown at higher magnification. Scale bars indicate 10 µm (A: Amyloid β plaques; and C) and 20 µm (A right panel, B and D)
Thereafter, we investigated whether the widespread increased expression of intracellular Aβ protein without diffuse Aβ plaques in the brain of some HSE patients was specific to HSV infection. For this, we studied Aβ and pTau expression in brain sections from a VZV encephalitis patient. The patient was a 72-year-old male who had no known history of dementia and died 15 days after the onset of disease. Abundant VZV RNA-positive cells were detected (Figure S3A), as well as VZV antigen-positive cells, indicative of lytic VZV infection. Aβ plaques and NFT, as well as intracellular Aβ protein, were observed in hippocampal sections, whereas only widespread prominent intracellular Aβ protein depositions were observed in medulla oblongata sections (Figure S3B). Similar to the HSE brains, we did not observe an association between the spatial relationship of virus-infected cells and AD-related pathological changes (Figure S3C). Overall, these results suggest that lytic α-herpesvirus, especially HSV, infection of human CNS neurons is not directly associated with aberrant Aβ or pTau protein expression.
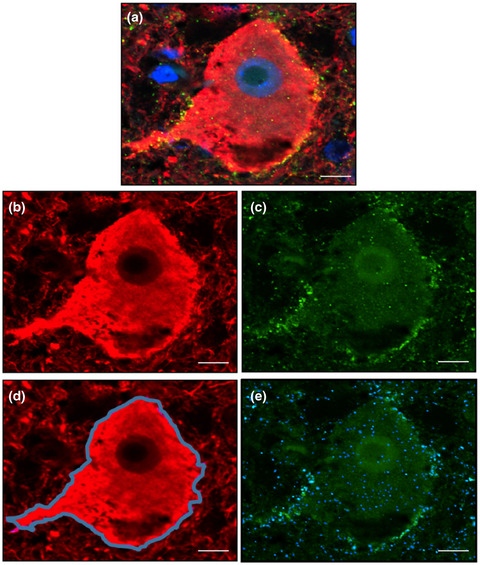
3.4 Expression of Aβ and pTau in brain of an AD patient with concurrent HSEA previous study reported that HSV-1 DNA colocalizes with Aβ plaques in brain of AD patients [16]. To investigate whether pre-existing AD pathology impacts the relationship between HSV-infected cells and Aβ and pTau expression, we obtained rare brain tissue specimens from an AD patient presenting with HSE. The patient was a 63-year-old female diagnosed with AD who developed HSE and died from septic shock resulting from HSE and aspiration pneumonia 1.5 wk after hospitalization. Analysis of six cortical brain regions revealed intermediate AD-related pathological changes, that is, amyloid Thal phase 5/5 and Braak NFT stage 4/6 (A3B2), consistent with dementia. HSV antigen-positive cells were abundantly detected in insular and temporal cortical tissue sections and less prominent in parietal and occipital cortex sections (Figure 5A). Importantly, HSV-1 infected cells were often found in close proximity to NFT and especially Aβ plaques (Figure 5B). However, AD-related pathological changes were not different between regions with and without HSV-1 infected cells. HSV-1 infected cells were situated around but did not colocalize with Aβ plaques and NFTs (Figure 5C). Similar to the other HSE patients, increased expression of intracellular Aβ protein was present in most HSV-infected cells, as well as non-infected cells (Figure 5D). Thus, AD and HSV infection may affect the same brain regions, but we did not observe enhanced AD-related pathology proximate to HSV infection.

Detection of HSV-infected cells in close proximity to Aβ plaques in the brain of an Alzheimer's disease patient with herpes simplex encephalitis. (A) Brain section stained for HSV protein (brown) by immunohistochemistry (IHC). Scale bar indicates 50 µm. (B) Consecutive brain sections were stained for Aβ (4G8) and pTau (Ser202, Thr205) by IHC or stained for HSV-1 RNA by in situ hybridization (ISH). Boxes indicate areas shown at higher magnification. Asterisks indicate the same blood vessel in consecutive sections. Scale bars indicate 500 µm (low magnification) and 100 µm (high magnification). (C, D) Brain sections were immunofluorescently stained for HSV protein (green), Aβ (red), and pTau (Ser202/Thr205; white) protein. Nuclei were stained with Hoechst-33342 (blue). Scale bar: 20 µm
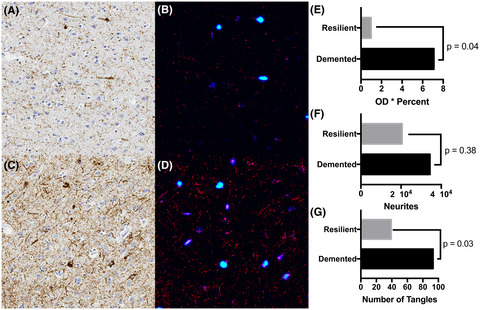
3.5 Neuroinflammation in HSE patients with and without AD-related pathologyBecause neuroinflammation is a major contributor to neurodegeneration in AD [42], we hypothesized that inflammatory cells rather than HSV replication itself could be associated with specific AD-related pathological changes. Brain tissue sections of all HSE patients, including the AD patient with HSE, indeed demonstrated widespread inflammation and gliosis in comparison to AD patients without viral encephalitis (Figure 6A and Table S4). HSE patients and the AD patient with concurrent HSE showed prominent perivascular cuffs, mainly composed of CD45POS CD3NEG Iba1NEG mononuclear cells – most likely macrophages or histiocytes – and lower numbers of CD45POS CD3POS T-cells (Figure 6B). Astrogliosis was observed in the brain parenchyma of all HSE patients, irrespective of Aβ or pTau expression pattern. This was also observed in the AD patient with HSE, but not as extensively in the AD patients without HSE (Figure 6C). Notably, while astrocytes colocalized with Aβ plaques of all AD patients and HSE patient #2, more intense GFAP staining was observed in astrocytes entangling Aβ plaques in the AD patient with HSE (Figure S4). Additionally, extensive microgliosis was observed in the brain parenchyma of the HSE patients and AD patient with HSE, but not AD patients without HSE (Figure 6D). Both amoeboid (Figure 6D) and ramified microglia (Figure S5) were observed throughout the tissue sections and in association with Aβ plaques and/or pTau in the AD patient with concurrent HSE and the HSE patients #2 and #5 (Table 1). Microglia were mostly amoeboid in HSE patients #1 and #3, whereas predominantly ramified microglia were present in HSE patient #4 (Figure 6D). Thus, although lytic HSV infection of the CNS induced significant neuroinflammation, the overall pattern of inflammation and gliosis was not associated with specific Aβ or pTau expression patterns.
 Neuroinflammation is similar in herpes simplex encephalitis (HSE) patients with and without Aβ plaques and/or NFT. Brain sections from Alzheimer's disease (AD) patients (n = 3, Table S4), HSE patients (n = 4, Table 1), and an AD with HSE patient (n = 1) were stained with (A) hematoxylin and eosin staining or IF stained for: (B) CD3 (green) and GFAP (white) (left panel) or Iba1 (green) and CD45 (white) (right panel); (C) Aβ (red) and GFAP (white); or (D) Aβ (red), Iba1 (green), and CD45 (white). Nuclei were stained with Hoechst-33342 (blue). Images are shown for the AD patient with HSE (B–D), AD patient #1 (C, D; Table S4) and indicated HSE patients. Boxes indicate the areas shown at higher magnification. Scale bar: 50 µm
4 DISCUSSION
Neuroinflammation is similar in herpes simplex encephalitis (HSE) patients with and without Aβ plaques and/or NFT. Brain sections from Alzheimer's disease (AD) patients (n = 3, Table S4), HSE patients (n = 4, Table 1), and an AD with HSE patient (n = 1) were stained with (A) hematoxylin and eosin staining or IF stained for: (B) CD3 (green) and GFAP (white) (left panel) or Iba1 (green) and CD45 (white) (right panel); (C) Aβ (red) and GFAP (white); or (D) Aβ (red), Iba1 (green), and CD45 (white). Nuclei were stained with Hoechst-33342 (blue). Images are shown for the AD patient with HSE (B–D), AD patient #1 (C, D; Table S4) and indicated HSE patients. Boxes indicate the areas shown at higher magnification. Scale bar: 50 µm
4 DISCUSSION
Cumulative evidence has led to the hypothesis that the endemic human neurotropic virus, HSV-1, could play a role in the pathogenesis of AD [17-19, 21, 22, 25, 43]. Here, we used clinical specimens from HSV-infected individuals to investigate the relationship between HSV infection and Aβ and pTau expression within human neurons in TG and brain. We report that latent HSV infection is not associated with Aβ or pTau expression in TG neurons located in the PNS. Similarly, lytic HSV infection in the CNS was not consistently associated with increased expression of intracellular Aβ or pTau proteins nor with the deposition of Aβ plaques and NFT. Analysis of unique material from an AD patient with concurrent HSE showed that the same brain regions are often affected by HSV and AD-related pathology, but did not reveal exacerbated Aβ or pTau production proximate to regions containing virus-infected cells.
Previous studies showed that increased anti-HSV IgM plasma levels and possibly anti-HSV IgG avidity can be a measurement of HSV reactivation that correlates with AD development [31, 44]. One proposed explanation is that genetic risk factors associated with AD development, especially APOE4 and paired immunoglobulin-like type 2 receptor alpha (PILRA), can directly impact lytic HSV-1 infection. The APOE4 allele is associated with increased frequency of HSV-1 DNA detection in the brain of AD patients and symptomatic HSV-1 reactivation (cold sores) [45, 46]. By contrast, asymptomatic oral HSV-1 shedding was not affected by APOE genotype [41], suggesting that APOE4 does not directly influence virus reactivation but may play a role in peripheral control of HSV infection. In this study, we did not observe differences in latent HSV-1 DNA load nor LAT expression in the TG of AD patients compared to control subjects without AD, or in APOE4 carriers compared to non-carriers. These data support the hypothesis that latent HSV-1 infection is not directly associated with increased risk of AD development.
Detailed analysis of rare human HSE brain specimens demonstrated that lytic HSV infection was not associated with increased levels of intraneuronal Aβ or Aβ plaque deposition in human neurons in vivo. Although we observed accumulation of intraneuronal Aβ protein in three HSE and one VZV encephalitis patient, Aβ depositions were widely present in both virus-infected and non-infected neurons and tissue sections. These findings are consistent with the ubiquitous and progressive accumulation of intraneuronal Aβ in the absence of extracellular Aβ plaques and NFT that is observed with aging [47]. Similarly, we only observed Aβ plaques in one HSE patient and locally (hippocampus, but not medulla oblongata) in one VZV encephalitis patient. The elderly age of these two patients, lack of an association between sites of virus replication and Aβ plaques, and the presence of both senile plaques and NFT suggests that these pathological changes were most likely part of ongoing development of AD, rather than induced by HSV infection.
HSV particles bind to Aβ protein and catalyze Aβ42 oligomerization, leading to Aβ aggregates that physically entrap virus particles [7, 48]. Consistent with the idea of HSV initiating the deposition of Aβ plaques in the CNS, a previous study used in situ PCR to detect HSV-1 DNA in the brain of AD patients and controls, and reported that HSV-1 preferentially colocalizes with Aβ plaques in AD patients [12]. Here, we did not detect viral antigen nor RNA within Aβ plaques in the brain of elderly individuals with HSV-1 or VZV encephalitis. Although HSV-infected cells were often found in close proximity to Aβ plaques and/or NFT in the brain of the AD patient with HSE, AD-related pathology was not restricted to regions with HSV-infected cells. We also did not observe HSV antigen or RNA in Aβ plaques in this patient. HSV-1 infection rapidly induces Aβ plaque deposition in the brain of genetically susceptible AD mouse models, but repeated HSV-1 reactivation and lytic virus replication in brain results in the progressive accumulation of both senile plaques and NFT [25]. While HSE is extremely rare in humans, asymptomatic HSV reactivation and viral spread to the CNS, as measured by the presence of viral DNA, occur frequently [9, 10, 16, 45]. Our data suggest that lytic HSV infection does not directly induce the formation of Aβ plaques in the human brain. This is possible because of the extensive neuronal cell death observed in HSE patients [49], which may limit the time available for neurons to produce substantial Aβ aggregates. Instead, repeated exposure to abortive HSV infections may be required to induce persistent Aβ plaques or could enhance Aβ oligomerization in existing plaques.
HSV infection of diverse human neuron models in vitro leads to the hyperphosphorylation of tau protein, including Ser202/Thr205, by cellular cyclin-dependent kinases and relocation of pTau to the nucleus [17-19]. We report that HSV-infected neurons in the brain of human HSE patients occasionally expressed nuclear pTau but only in individuals presenting with NFT. However, NFTs were not restricted to areas containing HSV-infected cells in HSE patients and the AD patient with HSE. These findings are in agreement with prior studies in 3xTg-AD mice (prone to develop NFT [50]), in which the degree and residues involved in tau phosphorylation in response to HSV infection varied between brain regions, and progressively increased with repeated viral reactivation events [25]. The heterogeneity of neurons in vivo – including the expression of specific cyclin-dependent kinases – could determine their differential susceptibility to HSV-induced aberrant tau phosphorylation and cellular localization.
Specific subsets of microglia and astrocytes colocalize with Aβ plaques in the brain of AD patients [51-53] and are thought to be involved in both early and advanced stages of AD pathogenesis [54, 55]. Lytic HSV infection induces robust innate immune responses in the brain, involving both microglia and astrocytes, which not only control ongoing virus replication but also causes immunopathology [56]. Prominent microgliosis and astrogliosis were present in all HSE patients, but we did not observe overall differences in glia cell morphology or abundance in patients with or without Aβ plaques and/or NFT. Interestingly, GFAP staining tended to be more abundant in Aβ plaque-associated astrocytes in brain sections from the patient with combined AD and HSE, compared to patients with either AD or HSE (Figure S4). Although these observations suggest that HSV infection may influence reactive astrocyte function in the brain of AD patients, more detailed analyses in more patients or experimental animal models are warranted.
In conclusion, we demonstrate that latent and lytic HSV infection of human neurons in TG and CNS is not consistently associated with aberrant Aβ or pTau expression. Human in vitro neuron cultures and murine AD models highlight potential mechanisms by which HSV infection could contribute to the initiation or perpetuation of AD. However, our data suggest that the human CNS could be more resilient to HSV-induced AD-related neuropathology than previously anticipated. Future studies comparing the effects of HSV infection on the healthy aging brain and AD brain may provide valuable insight into mechanisms by which HSV may affect AD pathogenesis.
ACKNOWLEDGMENTSWe thank Dr. Georges Verjans for the critical discussion of the data and Tamana Khemai-Mehraban for technical assistance. Research reported in this publication was in part supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under Award Number R01AI151290 (W.J.D.O.). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. This study was funded in part by a Human Disease Model Award 2020 (Erasmus MC).
CONFLICT OF INTERESTThe authors have no conflicts of interest to declare.
AUTHOR CONTRIBUTIONSWJDO conceived and designed the study. DNT and ATCMB conducted the experiments. DNT, ATCMB, VvD, RMV and WJDO analyzed and interpreted the data. EA and RMV contributed patient samples. DNT and WJDO wrote the manuscript.
留言 (0)