
記住我




Our social worlds emerge from, and persist within, our sensory and perceptual experiences. One of the great challenges facing the study of behavior and ethology has been to understand the sensory and perceptual world in which an organism lives. An organism's perceptual space (i.e., the umwelt)1 refers to the environmental surroundings to which an animal attends for survival and reproduction. All stimuli, social or otherwise, are filtered by an organism's sensory and perceptual systems. A “social umwelt” allows individuals of any particular species to respond appropriately to the natural diversity of cues and signals they encounter during social interactions with conspecifics. Social interactions occur within, and because of, these shared perceptual spaces. Through repeated social interactions, shared perceptual spaces provide the fundamental basis for the formation and expression of social relationships.2, 3In this way, the behavioral and neurobiological mechanisms of sociality and social bonding are inextricably linked with perception. However, approaches in both research and clinical settings conceptually, methodologically, and diagnostically delineate behavioral and physiological mechanisms as either social or sensory. This modular perspective both reflects and reinforces the significant gaps in our knowledge of how these processes co-occur and emerge.
Across species, patterns of social behavior reflect and exploit species-specific differences in perceptual spaces. Many species have exceptional sensory capabilities in certain domains, creating specific 'channels' of communication. For example, many species operate within perceptual spaces that extend beyond the human experience or human perception, such as ultrasonic vocalizations,4, 5 fine auditory temporal processing,6, 7 ultra-violet and infrared color vision,8-10 and chemosensory perception by the vomeronasal organ.11, 12 Differences in animal behavior reflect these differences in perceptual abilities, and behavior is often specialized within each modality (e.g., echolocation and hunting in bats).13 Key differences across species in social behavior (including parenting, courtship, and mate choice) reflect these sensory differences (e.g., vision for mate choice in primates14-16 or chemosensation and social recognition in rodents).17, 18 The co-evolution of social and sensory systems has been studied in over 70 species, including numerous invertebrates, fish, amphibians, birds, primates, and even whales.19, 20 The majority of research has focused on co-evolution of social behavior with visual or auditory systems,8, 19, 21, 22 but there is also evidence of co-evolution for social behaviors with vibratory and chemosensory systems.19, 23-25 Such comparisons across species highlight the potential magnitude of sensory differences, the specializations of key 'primary' modalities (e.g., visual, auditory, or chemosensory), and the embeddedness of social behavior within these sensory systems. Identifying species differences in 'primary' channels of communication is an important first step toward developing a unified framework to understand the reciprocal impacts of sensory and social processes.
1.1 An integrated sensory-social framework of pair bondingDeveloping frameworks that reflect the extent to which social and sensory systems are interconnected has the potential to greatly expand our understanding of social bonding. Although it has long been known that communication and perception are essential to social bonding, research at the interface of these processes has been restricted to a few key questions – largely outside of broader pair bonding research. For example, there is extensive research on female mate choice and social recognition. However, there is very little research on how patterns of dynamic communication vary across pairs or how these patterns of communication are shaped over time by pair bonding. Furthermore, even though the original mammalian pair bonding circuit26 includes the olfactory bulb, whether differences in sensory processing relate to differences in pair bonding remain unknown. These gaps in our knowledge are not specific to communication. In fact, we know surprisingly little about the mechanisms and consequences underlying individual differences in social bonding across species. We also currently have a poor understanding of how social bonds change over time27-31 or how such dynamic relationships produce enduring impacts on individuals' brain and behavior.32, 33 Such gaps in our understanding have implications for both basic and applied research. The majority of pair bonding research still conceptualizes pair bonding as a dichotomous state (individuals being either bonded or not bonded) rather than considering it as a continuous spectrum of individual differences in bond strength. One approach to deepening our understanding of social bonding broadly is to rely on an integrated social-sensory framework of bonding. Such a framework would emphasize the more mundane and dynamic aspects of the social interactions that culminate to form and reflect bonds.
In this review, we leverage what is known about sensory processing and pair bonding in two common study systems. We discuss the prairie vole (Microtus ochrogaster) and zebra finch (Taenopygia guttata) systems to explore such an integrated social-sensory framework of pair bonding. For each study system, we (1) describe the unique aspects of ethology/ecology for that species, (2) situate this knowledge within the broader research programs in each relevant field, (3) highlight the role of communication and perception in pair bonding, (4) review candidate neural circuits underlying the integration of communication, perception, and pair bonding, and (5) emphasize the extensive potential for neuromodulation of signal processing based on social experience. Prairie voles and zebra finches offer very different insights into how to develop and apply an integrated social-sensory framework to pair bonding. We conclude by discussing future directions that will extend this integration across behavioral, neurobiological, and genomic levels of analysis.
2 PRAIRIE VOLESMicrotus voles have become an established system for studying mammalian social structures, reproductive decision-making, and social behavior. In particular, prairie voles (M. ochrogaster) are best known for their impact on understanding the behavioral and neurobiological mechanisms associated with pair bonding and social monogamy.28, 29, 34-36 The greater vole study system is particularly powerful because closely related species differ in their levels of social organization, mating systems, and propensity to form social bonds, even though they often experience similar ecological pressures in nature.37-40 Montane and meadow voles (M. montanus and M. pennsylvanicus, respectively) are non-monogamous and do not exhibit biparental rearing of offspring.41 In contrast, the prairie vole is well known for its tendencies to form long-term monogamous pair bonds and to exhibit biparental family structure.29, 37, 42, 43 Comparative work between the monogamous and non-monogamous species has revealed that aspects of neural phenotype account for the different social organizations and mating systems these species demonstrate.18, 44
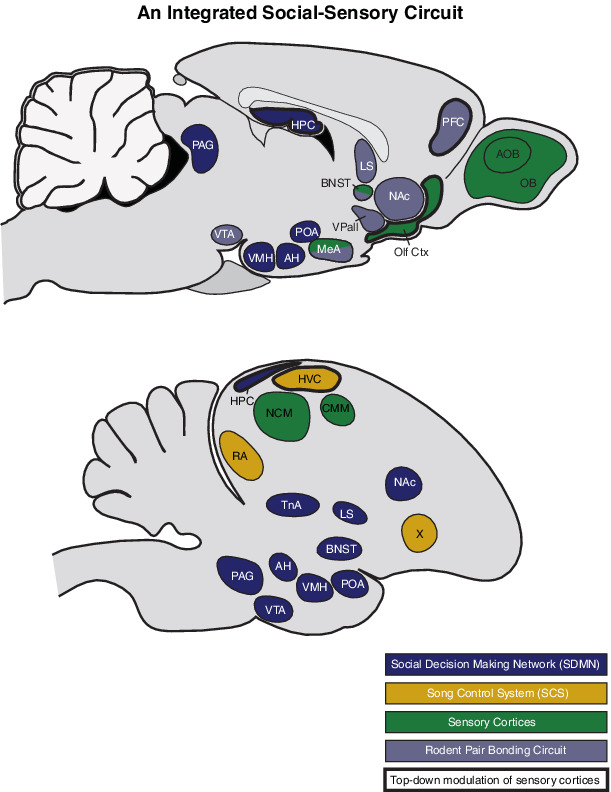
Pair bonding in prairie voles is very well documented, and robust behavioral assays have enabled the examination of social and neuroendocrine factors that influence the formation of pair bonds,42 which has led to an increasingly well-developed neurobiological model for mammalian pair bonding (e.g., see References 18, 26, 45, 46). As shown in Box 1, the neural structures that have been closely associated with prairie vole pair bonding include a combination of reward nuclei47 and structures that modulate social behavior.48, 49 This so-called 'pair bonding neural circuit' represents a subset of a larger network of interconnected forebrain structures (predominately limbic) that are collectively recognized for their general role in modulating social behavior and decision-making.50, 51 This 'social decision-making network' (SDMN) is an ancient network of highly homologous brain areas that has proven valuable in identifying neurobiological mechanisms involved in the regulation of social behavior across vertebrates. Arguably, the decision to form a pair bond falls safely within social decision-making, and it is reasonable to consider the pair bonding neural circuit as a sub-unit of the SDMN from both a behavioral and neuroanatomical point of view.

Defining the brain networks that support social communication and behavior has tremendous value for elucidating brain function and supporting broad translational applications for human mental health and well-being. The foundations and trajectories of prairie vole and zebra finch neurobiological research are quite different, but highly complementary. Together these two well-studied species offer complementary perspectives on what an integrated social-sensory communication circuit might look like. We believe that an integrated network including the well-described network associated with the control of social behavior, known as the social decision-making network (SDMN), in addition to neural circuitry responsible for chemosensory and auditory processing within the rodent and avian brains provide a platform for envisioning such an integrated social-sensory communication circuit.
The SDMN (denoted with blue) is a large network of interconnected forebrain (primarily limbic) and midbrain structures that are collectively recognized for their general role in modulating social behavior and decision-making.51 The mammalian pair bonding circuit (denoted with gray) largely represents a subset of the SDMN nodes, and is comprised of olfactory processing areas of the brain, reward nuclei,47 and structures that modulate social behavior.48, 49 Additionally, the avian song control system (SCS; denoted with yellow) represents a well-documented set of specialized brain regions that are central for the production and perception of vocal communication among avian species. The full pair bonding circuit and SCS are described below for context. Key sensory cortices are highlighted in green (related to auditory processing in zebra finches and chemosensation in voles).
Rodent pair bonding circuit: At its core, the prairie vole pair bonding neural circuit combines reward with facilitated learning of social identity (based on olfactory information) to enhance the social valence of a particular conspecific, resulting in pair bond formation.60 According to this model, mating behavior stimulates the ventral tegmental area (VTA) to activate dopaminergic reward systems in the prefrontal cortex (PFC) and the nucleus accumbens (NAc), while simultaneously stimulating the release of the neuromodulatory peptides oxytocin and vasotocin (OT and VT respectively, see)59 within the PFC, NAc and ventral pallidum (VPall).36 Concurrent with the release of OT and VT, chemosensory information originating from a mate is detected by sensory neurons in the main olfactory and vomeronasal sensory epithelia, initially processed in the olfactory bulb (OB) and accessory olfactory bulb (AOB), then projected to a number of (mostly cortical) structures within the olfactory pathway (olfactory cortex, Olf Ctx) before being integrated in the medial amygdala (MeA).73 In response, the MeA and the bed nucleus of the stria terminalis (BNST) simulate release of VT in the lateral septum (LS). The presence of OT in the MeA and VT in the LS facilitate chemosensory learning of the specific chemical signature of each partner's respective mate.26, 60 This concurrent activation of the reward system with facilitated sensory learning results in a conditioned partner preference (i.e., a pair bond). The prairie vole pair bonding circuit highlights a clear circuit that integrates sensory-social information (OB/AOB ->Olf -> MeA). However, as discussed throughout the paper, the extent to which this circuit is involved after initial pair bonding, and/or the extent to which neural circuits shape perception and social bonding (including top-down modulation of sensory processing) remains largely unexplored.
Avian song control system (SCS): The SCS is a series of discrete brain nuclei involved in the production and learning of vocalizations. The SCS has traditionally been studied with respect to male song. This SCS neural circuit includes two primary pathways. The first is the motor pathway comprised of the HVC (proper name), which projects to the robust nucleus of the arcopallium (RA). The RA, in turn, projects to tracheosyringeal portion of the hypoglossal nucleus (nXIIts), which is involved in motor projection in the syrinx. This pathway is required for normal vocal (predominately song) production throughout life. The motor pathway also intersects with the auditory regions of the avian pallium, which is analogous to the mammalian auditory cortex. These auditory regions consist of the caudomedial nidopallium (NCM) and caudomedial mesopallium (CMM) via reciprocal projections through the nucleus interfacialis (NIf). HVC also projects to the second primary pathway in the SCS - the anterior forebrain pathway. In this pathway, HVC projects to Area X (homologous to mammalian basal ganglia and a strong target of midbrain dopamine projections), the medial nucleus of the dorsolateral thalamus (DLM), and the lateral magnocellular nucleus of the anterior neostriatum (LMAN; a frontal cortex-like nucleus). The two pathways again interact via projections from LMAN to RA.125, 131, 219 In this diagram, we highlight the nuclei from the SCS that have been directly and broadly implicated in acoustic communication (HVC, RA, and Area X). There are numerous candidate pathways that putatively integrate information between the SDMN and SCS – here we highlight VTA-Area X162, 163 and mPOA -> PAG -> HVC.164, 165, 169
For both species, we include the interconnected social behavior circuit and key dopaminergic projections to this circuit (together making the SDMN). These include (1) the hypothalamic structures - ventromedial hypothalamus (VMH), anterior hypothalamus (AH), and the preoptic area (POA), and (2) reward circuitry - periaqueductal gray (PAG), and the medial amygdala (MeA) in mammals or the nucleus taenia (TnA) in birds. Beyond initial pair bonding, the broader SDMN is likely involved in social decision-making more broadly and may be crucial for regulating pair-directed behavior and social cognition throughout the lifetime. Further research focused on the functional connectivity between the SDMN and communication circuits promises to reveal a more comprehensive view of the neural mechanisms subserving social behavior.
Finally, we highlight key brain regions that function in top-down neuromodulation of sensory processing (denoted with bold outline). In voles, although the PFC is accounted for in the pair bonding circuit, other regions such as HPC and Olf Ctx are not. These ‘top-down’ areas that are often not included in efforts to understand pair bonding merit closer attention if we are to achieve a more complete understanding of the neural processing of this important social behavior (e.g., see Reference 220)
Although prairie voles typically form strong and enduring pair bonds, there remains extensive individual variation in the extent and expression of social bonding52 – a phenomenon that has received relatively less attention. Notably, prairie voles exhibit alternative mating tactics in nature, in some cases either remaining single or forming pairs and mating outside the pair opportunistically.29 The alternative reproductive strategies among vole species and alternative mating tactics observed within species demonstrates that profound variability exists within the social structures of voles. Thus, this system allows deep exploration of the mechanisms underlying reproductive decision-making and social bonding. Here, we discuss mechanisms of social communication among prairie voles in the context of pair bonding to illustrate the dynamic relationship between sensory systems and social cognition in mammals.
2.1 Chemical communication and pair bondingIt is well established that chemical communication plays an essential role in many aspects of social communication and reproduction in rodents.53, 54 The chemosensory system is crucial for bonding because the expression of a pair bond requires the ability to recognize and remember the specific identity of a mate.55 In prairie voles, like most rodents, social recognition depends on chemosensation of an individual's chemical profile. Thus, the chemosensory pathway provides the fundamental information necessary for the neural circuitry of pair bonding.26 Notably, prairie voles do not pair bond without an intact olfactory bulb42, 56 or a functional vomeronasal organ.57, 58 The presence of oxytocin (OT) in the medial amygdala (MeA) and vasotocin (VT, see)59 in the lateral septum (LS) facilitate chemosensory learning of the specific chemical signature of each partner's respective mate.26, 60 This concurrent activation of reward systems with facilitated sensory learning results in a conditioned partner preference, that is, a pair bond.
The relationship between sensory input and behavior is commonly conceptualized as unidirectional (i.e., sensory input leads to behavioral output). However, this perspective falls far short of accurately describing the dynamic nature by which sensory information influences, and in turn is influenced by, the ongoing social and sensory experiences associated with social bonding. Although being bonded requires social recognition of one's mate,55 the very ability to discriminate any particular chemical profile within the extremely complex chemical environment in which an animal lives is dependent upon continual refinement of a dynamic system of signal processing and chemosensory learning.61 Furthermore, it is likely that chemosensory communication between pair bonded partners is important beyond initial partner preference formation. Across rodents, chemical signals are known to encode a staggeringly diverse array of behaviorally and ecologically relevant information. Chemical signaling is known to facilitate detection and evaluation of reproductively available potential mates to maximize fecundity,62, 63 detection and avoidance of sick or parasitized conspecifics,64, 65 evaluation of age,66 quality of diet,67 and levels of stress or hunger in nearby conspecifics,68, 69 as well as detection and avoidance of predators or other dangerous environmental factors.70-72 Clearly, there is the potential for a tremendous amount of behaviorally relevant information to be encoded in chemosignals beyond sex and individual identity. The importance of chemical communication between pair bonded partners, beyond social recognition and partner preference formation, remains largely unexplored.
2.2 Neurobiological regulation of chemosensation during pair bondingAs shown in Box 1, the olfactory bulb (OB) is included in the mammalian pair bonding circuit, and it has long been recognized that information from chemical signals is important for social bonding. Chemosensory information originating from a mate is detected by sensory neurons in the main olfactory and vomeronasal sensory epithelia. This information is initially processed in the olfactory bulb and accessory olfactory bulb, then projected to a number of (mostly cortical) structures within the olfactory pathway before being integrated in the MeA.73 Ultimately, the diversity of individual chemical ligands that can be detected by chemosensory systems is based on the genetic repertoire of receptor genes in the genome. The functional repertoire of receptor proteins present in the sensory epithelium is adaptively established by control of gene expression74 and experience-based survival of sensory neurons75 resulting in functionally optimized sensory receptor profiles. However, this description does not accurately portray the dynamic nature of chemosensory processing. Decoding raw chemosensory input is an extremely complex task requiring multiple mechanisms working in concert to filter unwanted information and increase signal-to-noise ratios to discriminate functionally relevant signals embedded in a complex background.76-78 To accomplish this task, a wide array of functionally diverse neuromodulatory mechanisms is employed at multiple levels within the chemosensory pathway.
A great many genes are involved in chemosensory processing and producing behavioral responses to chemical stimuli. Thus, variation in genetics and mechanisms governing gene expression have the potential to greatly impact the function of this system. To cite just one illustrative example, consider the oxytocin receptor (OTR), which is encoded by a single functional gene (otr). Recall that OT is necessary and sufficient for pair bonding in prairie voles.26 Unsurprisingly, the presence and density of the OTR protein has a large impact on vole social behavior. OTR expression within the mesolimbic pathway is required for pair bonding.18 Additionally, region-specific densities of OTR predict social organization between monogamous and non-monogamous voles species,44 and manipulation of OTR density via pharmacological or genetic means alters pair bonding.79-82 In addition to mediating social behavior, OT and OTR play a key role in neuromodulation and early signal processing within the olfactory pathway.77 Furthermore, OT-OTR binding in the rat OB enhances the signal-to-noise ratio during olfactory information processing resulting in improved olfactory specificity and discrimination of social signals.77 Behaviorally, this results in increased exploration of social chemical signals during interactions with conspecifics and enhanced social recognition.77 Thus, the release of OT and the presence and density of OTR accomplish several functions, including asserting critical control over pair bonding, and modulating the OB to enhance sensory information processing and social recognition.
The OB itself contains intrinsic neuromodulatory mechanisms and memory circuitry capable of modifying sensory information based on prior experience. Thus, the OB is able to perform a surprising amount of signal processing prior to sending the information to other areas of the brain.76 Mitral cells and periglomerular cells in the OB receive input directly from olfactory sensory neurons and process the signal before relaying information to the piriform cortex and other higher order processing areas.76, 83-85 As sensory input is processed through the OB, inhibitory granule cells act to modulate, dampen, and filter the output from mitral cells to increase the signal to noise ratio. Olfactory learning is accomplished as sensory information guides the development of granule cells, such that only specific mitral cells are inhibited thereby resulting in cleaner signals representing learned odorant profiles.76, 85 These intrinsic memory circuits are maintained and expanded upon as adult neurogenesis incorporates new granule cells into the OB, which may subsequently be arranged to include additional odorant profiles.85
In addition to intrinsic mechanisms, the OB receives a great deal of extrinsic top-down neuromodulatory input from a diverse range of brain regions involved in memory formation such as the hippocampus (HPC, via the lateral entorhinal cortex), anterior olfactory nucleus, piriform cortex, and prefrontal cortex (PFC).86, 87 Cortical projections from these regions primarily activate a subset of granule cells within the OB, which act to further modulate and filter sensory information based on learned odorant profiles.88
Memory circuits also exist further along the chemosensory pathway including in the MeA, bed nucleus of the stria terminalis (BNST), LS, nucleus accumbens (NAc), and PFC, which further modulate sensory information while simultaneously functioning to produce appropriate behavioral responses to social chemical signals.26, 89, 90 Although the olfactory pathway is affected by multiple neuromodulatory mechanisms, the extent to which these mechanisms shape and are impacted by emerging pair bonds remains unclear.
2.3 Behavioral evidence for enduring effects of pair bonding on social recognitionBehavioral evidence provided by prairie voles supports the hypothesis that social bonding produces lasting impacts on chemosensory processing and social cognition. Blocker and Ophir (2015) showed that paired male voles exhibit social recognition of individual females based on chemical profiles, whereas single voles (i.e., un-paired) do not discriminate between individual females.91 Paired voles presumably benefit from enhanced individual recognition of females, as pair bonding inherently requires partners to recognize and remember the individual to which they are bonded. Likewise, single voles might benefit from indiscriminate mating, which does not require social recognition of individual females. These findings suggest that differences in sensory processing among single and paired voles affect social recognition of potential mates, although the specific neural mechanism underlying this shift in social cognition is still unknown.
Given how important chemosensory information is for prairie vole social behavior, the fact that social recognition is fundamental to pair bonding is easily taken for granted such that it is assumed to simply be a static foundational component of a pair bond. However, behavioral studies call into question this assumption, and more readily support the hypothesis that the ability to recognize a conspecific is dynamic, depending on social and environmental context. This is evident when examining social recognition abilities of single male voles.92 Zheng et al., (2013) demonstrated that the interpretation of social chemical signals by prairie voles depend on the social and environmental context in which they were sensed. In this study, single male voles demonstrated social recognition using a habituation-dishabituation paradigm to assess discrimination between individual males. Zheng et al. (2013) found that male's social recognition of other males was most pronounced in a familiar environment, and weakest in a novel environment absent of social cues. Consistent with Blocker and Ophir (2015), single males did not discriminate between females, but they did discriminate between males, indicating contextual valence and/or motivation to attend to social cues in a sex-dependent manner. This result is even more peculiar considering that chemical profiles of females are known to convey sufficient information to allow social recognition and individual discrimination.91 Thus, key differences in processing and interpreting chemosensory information not only exist, but they are particularly sensitive to social context. The emphasis on male, but not female recognition among single males suggests the existence of a mechanism that favors individual recognition of potential neighboring competitors while also favoring indiscriminate mate selection. Moreover, such a mechanism can be particularly beneficial for male voles that do not have a pair-mate.29 Although the specific neuromodulatory mechanisms underlying these observations have not yet been elucidated, OT and VT are well known to influence the recognition process.93 This is one example of how dynamic chemosensory perception shapes social behavior phenotypes, and these behavioral findings constitute evidence that adaptive sensory modulation impacts reproductive decision-making in prairie voles.
2.4 Enduring effects of pair bonding on chemosensory processingWhat neurobiological mechanisms might account for the contextual effect of social bonding on social recognition? Mating, exposure to chemical social signals, and formation of pair bonds are often associated with long-term mechanisms of neuroplasticity in key structures within the chemosensory processing pathway and in the SDMN. The well-characterized “Bruce effect”, in which pregnancy is spontaneously aborted after exposure to chemical cues from an unfamiliar male,94 provides an intriguing example. Specifically, the Bruce effect is normally avoided by chemosensory imprinting driven by adult neurogenesis in the accessory olfactory bulb (AOB) occurring in response to continual exposure to familiar male chemical cues.95 Hence, this illustrates the powerful effect neuroplasticity has on physiology and behavior. Indeed, adult neurogenesis provides a mechanism of neural plasticity allowing adaptive optimization of neural circuitry to meet current social or environmental demands.96, 97 In prairie voles and another socially monogamous rodent (Mus spicilegus), adult neurogenesis is observed in the MeA following formation of a pair bond.98, 99 Further evidence from prairie voles demonstrates that mating and exposure to social chemical signals stimulates neurogenesis in the subventricular zone (SVZ), dentate gyrus (DG) of the hippocampus, and the rostral migratory stream (RMS), which produce neurons ultimately destined for the OB100 and AOB.83 In contrast, social isolation reduces neurogenesis and survival of neurons in the MeA, medial preoptic area of the hypothalamus (mPOA), and the DG and ventromedial HPC.101 It remains unclear what functions this impressive neuroplasticity may serve beyond initial pair bonding.
Neuroplasticity within these circuits may also exist at the level of gene regulatory networks, which reflect and result in changes in neuronal functioning. Pair bonding is associated with robust changes to gene expression in the NAc of prairie voles. Notably, in addition to its critical role in pair bond formation, the NAc is highly interconnected with the olfactory tubercle, which receives input directly from the OB,102 providing yet another opportunity for sensory integration with the processing of social behavior. Patterns of gene expression in the NAc are significantly altered during early bond formation occurring within the first 24 h of cohabitation and continue to change during the first several weeks following formation of a bond.79 The subset of differentially expressed genes identified during early bond formation only partially overlap with those identified three weeks after formation of a bond, suggesting that separate transcriptional mechanisms have roles within early pair bond formation and later during bond maintenance. The transcriptional profiles associated with pair bond maintenance are stable from two to at least six weeks after bond formation, but these profiles erode within four weeks if partners are separated, suggesting that transcriptional mechanisms underlying pair bond maintenance persist as long as the partners remain paired.103 Gene expression affecting pair bond formation may also be altered on a more permanent basis via epigenetic modifications within regulatory regions of the genome. To illustrate this, we again refer to the earlier example involving OT and OTR interactions. Epigenetic modifications within the prairie vole OTR gene promoter region within cells of the NAc increase OTR density, and thereby increase sensitivity to OT and facilitate pair bond formation in both males and females.79, 80 Although pair bonding can have both profound transient and enduring impacts on pair bonding circuitry, it remains unknown whether and to what extent this occurs within the OB and other chemosensory processing circuitry.
2.4.1 Next stepsThe importance of chemosensation in rodents has long been known, and the OB is included in the mammalian pair bonding circuit. Nonetheless, there are striking gaps in our basic knowledge about the role of chemosensory communication beyond social recognition for pair bonded individuals. Moreover, as highlighted above, the transduction and interpretation of chemosensory information is not as straightforward as it may seem. Sensory information is modified by multiple functionally disparate neuromodulatory mechanisms acting in concert to adaptively optimize the social interpretations of chemical signals in intricate ways that we are just beginning to understand. Each of these diverse mechanisms are individually capable of modulating sensory processing. In a living animal, however, most of these mechanisms simultaneously and continuously modify neural activity.84 This dynamic nature of chemosensory processing means that the neural representation of a particular chemical profile is highly plastic and adaptive, and it is dependent on multiple factors such as prior experience and learning, environmental and social context, and current physiological state.88 This system of feedback adaptively and effectively allows discrimination of minute, but often critical, differences among chemical signals driving social decision-making. The extent to which pair bonding shapes sensory processing within the OB and chemosensory circuity remains unknown.
3 ZEBRA FINCHESZebra finches are small songbirds native to the arid and semi-arid regions of Australia.104 The natural ecology and ethology of the zebra finch makes them an excellent system for studying sociality and communication. Zebra finches are highly gregarious, form life-long sexually monogamous pair bonds, divide parental care equally between partners, and breed opportunistically – one of many adaptations to evolving within extreme and unpredictable climate and environmental conditions.104-108 Zebra finches are also one of the most well-studied avian species in the lab or field.109, 110 Zebra finches are a predominant study system for a range of biological disciplines including genetics/genomics, physiology, behavior, ecology and neuroscience.109, 110 Indeed, the second avian genome sequenced was the zebra finch.111
Monogamy is overwhelmingly the most common mating system in birds, exhibited by ~90% of species.112-115 Yet even relative to other small songbirds, zebra finches form remarkably strong pair bonds. Zebra finch partners actively maintain their bonds throughout life, even during long non-breeding periods, and extra-pair courtship is very rare.104, 106, 116 Zebra finches are one of the most well studied avian species with respect to monogamy and pair bonding.106, 110, 116-
留言 (0)