
記住我




Finding food is essential for animal survival. Thus, a diverse and adapted set of behaviors have evolved to allow animals to effectively forage in their niche environment. This adaptation to the environment, including its risks and its available resources, applies to invertebrates as well. This includes naive hawkmoths preferring the smell of flower tubes with an optimal length for their proboscis and feeding while in flight to avoid ambush predators.1 Female mosquito approaches unsuspecting hosts by first identifying plumes of exhaled CO2, then honing in on human skin odorants and visual cues.2 Ants track the sun and count their steps during foraging expeditions to find their way home again3, 4 and a species of hermit crab can forage across terrestrial, marine and freshwater environments,5 showing remarkable diversity.
Broadly, foraging involves a balance between exploration and exploitation, where exploration is the search for profitable food sources and exploitation is the localized search and feeding in a known food patch.6 Despite the variety of foraging strategies spanning diverse habitats, feeding apparatus and mobilities, the basic principles of foraging behavior between invertebrates can be well-fitted by the same class of models.7 A key similarity is for example state changes between a fast exploration mode (roaming) and a slow, local exploitation mode (dwelling). This strategy is effective at maximizing food encounters and is present in both Caenorhabditis elegans as well as in Drosophila melanogaster.8-10
While multiple cues are integrated when navigating toward food, a key sense used in navigation across phyla is the sense of smell. Invertebrates from the crawling worm to the flying fruit fly all rely on odors to locate their next meal over long distances11-14 and the mechanisms of invertebrate olfaction in general are well understood,15 especially in mosquitos and flies. Recent work has developed multiscale connections spanning from the structure of the key odor receptor family Orco,16 to behavioral and neural data linking navigation and the coding of odor stimuli17 and finally to the connectome and projections of the antennal lobe, which is the key brain area for sensory integration in insects.18, 19 In contrast, how odor affects food intake among invertebrates is vastly understudied. Anticipatory feeding behaviors, such as motor20-22 and digestive priming,23-26 prepare the animal for rapid ingestion and reduce the time spent exposed to predators,22, 27, 28 for example. One can therefore argue that odor cues serve a second and equally vital function during foraging by preparing the animal for efficient food exploitation (odor priming), which deserves greater attention in the fields of invertebrate foraging and olfaction.
To gain a complete understanding of both exploration and exploitation in foraging requires simultaneous measurements of animal motion, food intake and odor concentration. Although it would be desirable to perform such measurements in natural environments with realistic odors, currently studies are only fully tractable using laboratory animals. Nevertheless, invertebrate model organisms are particularly suited for investigating the role of smell in exploitation because of their small size, early maturation, large brood sizes and the plethora of genetic tools and databases that are already established.29-32 Food intake can be well monitored and accurate assays can be designed to mimic increasingly more realistic odors and resource distributions,33-35 hence invertebrate models can help fill the gaps in our understanding of foraging behavior.
In this review, we cover the role of odors in invertebrate foraging, with a focus on data from model invertebrates like D. melanogaster and C. elegans. We first briefly summarize how odor is used as a cue in exploration before presenting evidence for odor's role in exploitation. Moreover, we will discuss current models of foraging behavior and how they can better reflect organismal energy balance by explicitly accounting for odor priming. Lastly, we suggest experiments based on recent advances in technologies and assays quantifying invertebrate behavior in order to better understand the role of odor sensation in both the exploration and exploitation phases of foraging.
2 EXPLORATION—ODOR AS A CUE 2.1 Smell is the key sensory modality for food detectionWhile invertebrates can access multiple sensory modalities, the odor remains one of the most important cues for foraging.11-13, 36 Unsurprisingly, in Drosophila larvae and C. elegans, sensing food odors confers a fitness advantage by allowing animals to detect scattered food sources or through odor-dependent alterations in the organismal metabolism that result in prolonged lifespans.37 Furthermore, odor-sensory cues are vital for the efficient localization of food sources in specific niches. When naive hawkmoths from two species of the same subfamily were presented with odor and a visual stimulus in a wind tunnel, nocturnal Deilephila elpenor placed more importance on the odor, while the diurnal Macroglossum stellatarum strongly preferred the visual stimulus.38 This is in line with the hawk moth subfamily's nocturnal ancestry, where their keen sense of smell allows them to locate food in the dark when visual stimuli such as flower shape and color can be less reliably distinguished. Consequently, hawkmoths are set apart from visual, light-dependent foragers like M. stellatarum by partitioning themselves temporally into a different niche, thereby reducing competition and improving resource acquisition.
Odors can also travel farther than sightlines allow in dense terrain, and odor plumes can stretch up to tens of meters.39 In particular, long-range foragers such as bumble bees, moths and certain marine invertebrates rely on odor cues to guide their movements.39-43 For example, the bumblebee relies solely on odor rather than incorporating visual cues when locating small flowers. Visual cues only replace odor in cases of large flowers with a low probability of odor encounter.44 Evidence suggests female mosquitos begin host localization by first scenting fluctuating CO2 levels downstream of a human as far as 10 m away. After following the CO2 plume within a 1 or 2-m proximity of the target, they may then leave the plume and only rely only on human skin odorants and visual cues for close approach and landing.2 Although army ants are known to have exceptionally poor vision, as is the case with Eciton hamatum, they are still able to locate the few species of ants that make up their specialist diets by detecting and discriminating among the many odors present in their neotropical habitat.45 Smell is therefore a key mid- to long-range sense in diverse habitats.
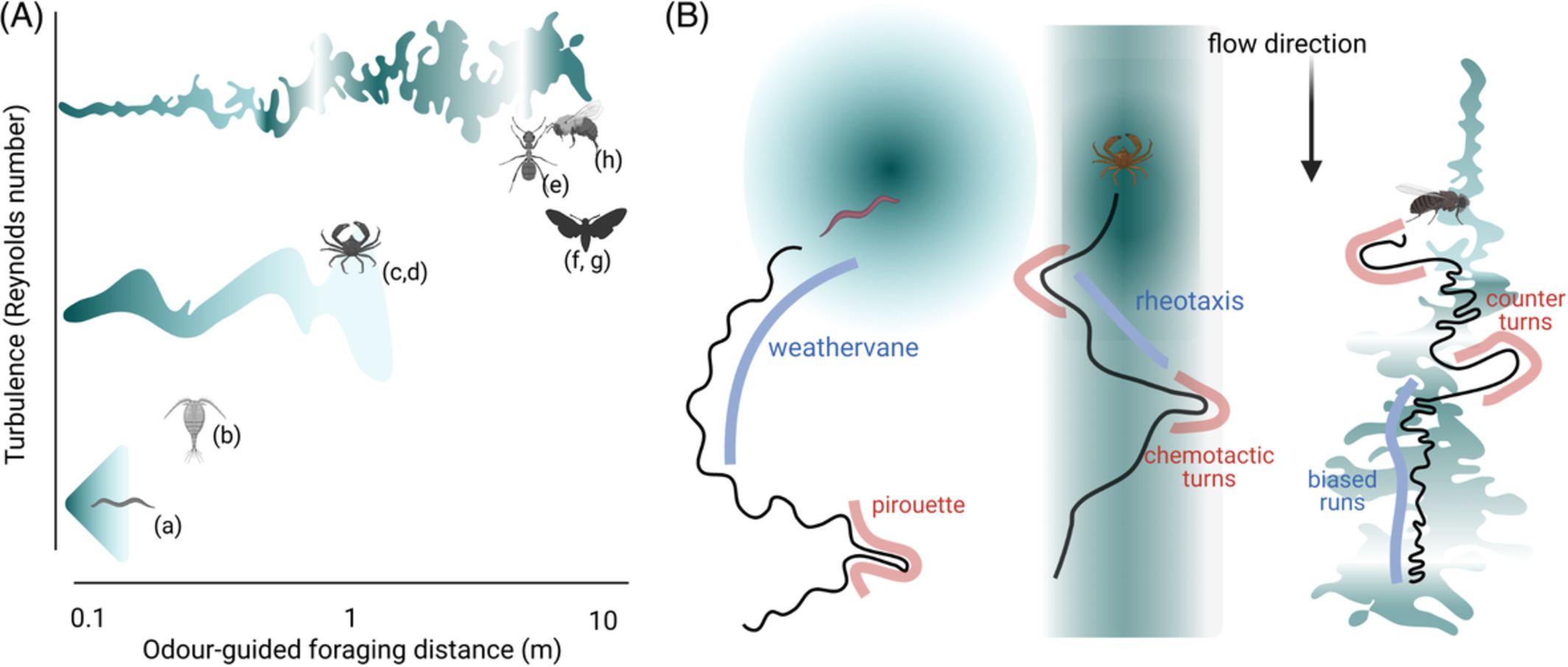
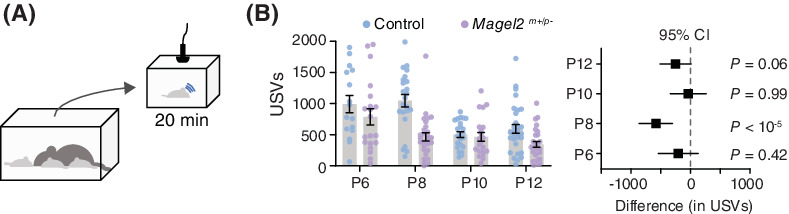
2.2 Common principles of odor-guided navigation at different scalesEffective navigation in search of food depends on the specifics of the environment, and particularly what form the odor cue is expected to take. In environments with laminar flow, odorants form stable, long gradients that allow for smooth gradient tracking such as chemotaxis (following a chemical signal).46-49 Moderate wind or water flows result in longer odorant tubules that still show well-defined gradients, but must be integrated into the animal's tracking through rheo- or anemotaxis (movement following the water- or wind direction42, 43, 50). With larger wind or water flow speeds, odor plumes become turbulent, therefore requiring complex and rapidly adapting search strategies to effectively navigate (Figure 1A).39-41, 51, 52 Locomotion plays an integral part in how animals experience these odor plumes. Depending on the animals' size and speed of locomotion, they will sense vastly different time-varying odor stimuli as they move relative to the odor. Consequently, the animal tracks from these three navigational regimes will show distinct statistics of behavioral parameters such as run length, turn direction and frequency and directional bias (Figure 1B).
 Typical odor plumes and navigational strategies for invertebrates. (A) Typical plumes and odor-guided foraging lengths encountered by roundworms (a),46, 47 copepods (b),49 blue crabs (c)42 or green crabs (d),43 desert ants (e),51, 52 gypsy or hawk moths (f, g)41 and bumble bees (h).39, 40 Odors transition from smooth gradients to turbulent odor plumes with increasing travel distances. (B) Navigational strategies of three species encountering odor plumes of different types. C. elegans,46-48 blue crabs50 and walking D. melanogaster.53 Irrespective of the strategy followed, navigation can be divided in runs (light blue) and turns (pink). Odor concentration is shown in dark green
Typical odor plumes and navigational strategies for invertebrates. (A) Typical plumes and odor-guided foraging lengths encountered by roundworms (a),46, 47 copepods (b),49 blue crabs (c)42 or green crabs (d),43 desert ants (e),51, 52 gypsy or hawk moths (f, g)41 and bumble bees (h).39, 40 Odors transition from smooth gradients to turbulent odor plumes with increasing travel distances. (B) Navigational strategies of three species encountering odor plumes of different types. C. elegans,46-48 blue crabs50 and walking D. melanogaster.53 Irrespective of the strategy followed, navigation can be divided in runs (light blue) and turns (pink). Odor concentration is shown in dark green
For example, the roundworm C. elegans in the laboratory mostly experiences laminar flow or static environments with smooth gradients. When C. elegans navigates to a food patch, it will use random reorientations called pirouettes and smooth curved trajectories as it reorients toward the highest odor concentration (Figure 1B), reacting to the smooth increases in odor concentration as it approaches the food.46-48 In contrast, blue crabs encounter smooth odors on a background of active flow, thus leading to navigation upstream along “tubules,” with chemotactic (chemical-induced) turns at the edge of the odor column.42, 50 Surprisingly, a similar “counter-turning” behavior at the edges of an odor stripe has also been shown in worms embedded in a flow environment, despite crabs being orders of magnitude larger in size and typically experiencing much larger flow rates.54
Airborne insects like moths or D. melanogaster experience a very different environment, which is dominated by air flow that will disrupt any smooth gradients and result in turbulent plumes that are neither continuous, nor form a stable, smooth gradient of chemicals (Figure 1A). The navigational strategy of walking flies is thus composed of runs biased toward the (expected) odor source, and counter-turns when the plume is lost55 (Figure 1B). Interestingly, a similar strategy is employed when aquatic arthropods encounter turbulent plumes.56 Without smooth odorant gradients simple strategies that work in laminar environments fail, for example, comparing the current and preceding odor concentrations to determine which direction is closer to the source. Given their greater speed, the sensory experience is even more complex for flying insects, as their own wingbeat can cause changes in the odor plumes.57 Such environments require adapted sensory strategies, which often integrate both the odors, as well as the wind speed and the animal's own wingbeat frequency to determine an optimal flight direction.58-60 Further investigation is needed for predatory feeding, where both the food source (prey) and the predator are moving. In this case, the landscape of odors and locomotion is even more complex, which is beyond the scope of this review.
It is surprising that similarities in odor tracking strategies were identified despite the different environmental characteristics, scale and type of locomotion.61 At its core, any navigational strategy consists of runs in the estimated direction of the odor source, and re-orientations when the odor is lost or reduced. Extracting statistics from animal tracks during odor tracking and comparing these to stochastic processes such as biased random walks, continuous-time random walks or Lévy flights has elucidated the mechanisms involved in generating these trajectories.48, 62 In turn, these stochastic models make predictions about the required neuronal activity underlying these navigation behaviors.63, 64 Overall, odor tracking in navigation has been extensively studied across species and is thus relatively well understood.7
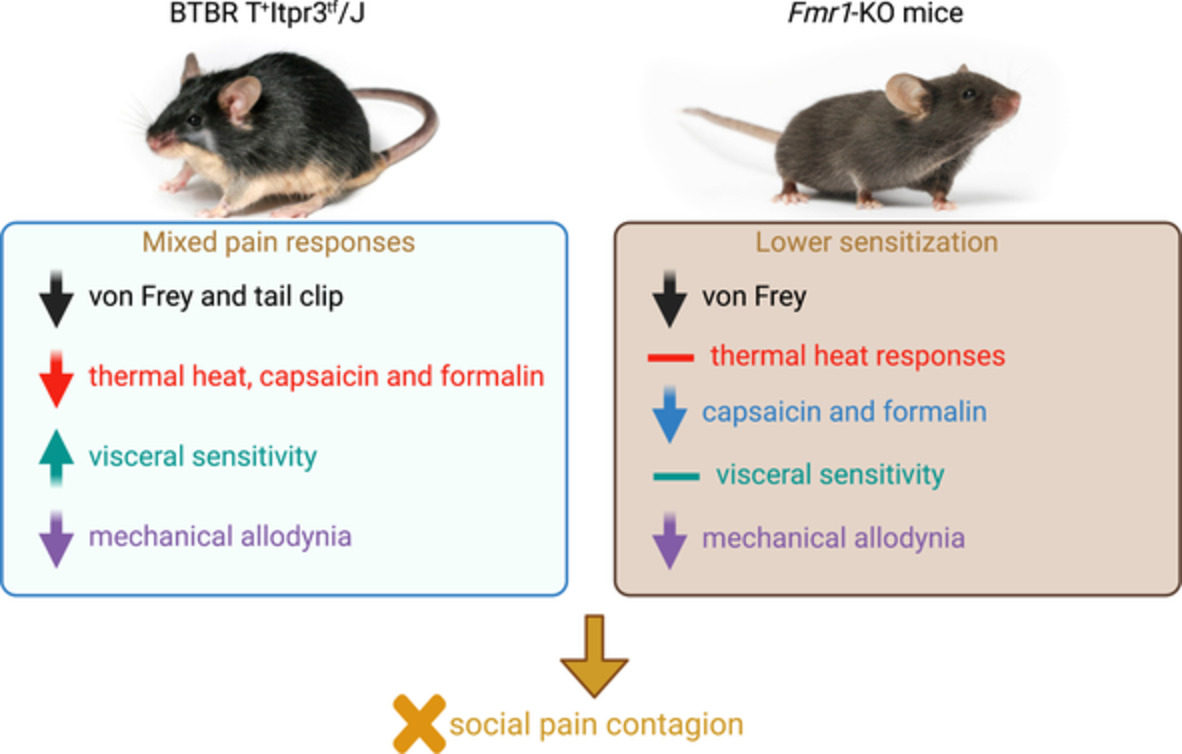
3 EXPLOITATION—ODOR AS A PRIMER 3.1 Odors help appraise the value of food sourcesOdor not only indicates the direction toward a food source but also its quality and type, leading to the more effective exploitation of high-value food or food filling a nutritional gap, when the animal is provided with a choice (Figure 2A). Besides fulfilling energy requirements, food choices additionally satisfy demands for specific nutrients.65, 66 For example, terrestrial gastropods selectively choose plant species and fresher leaves to feed on through olfaction.67 Drosophila adapt foraging choices to amino-acid needs based on odors, such as females replenishing their reserves after mating by taking more sips of amino acid-rich yeast compared with sucrose, while virgin flies are uninterested in yeast.68 More than 70 years ago, Lindauer showed that the addition of scent to a known unscented food source led to an initial decrease in honeybee waggle dances compared with controls, where the number of waggle dances was used as a proxy for food attractiveness.69 Ants will interrupt feeding if the scent of the food contradicts their prior experiences, even if the energy value of the food, as perceived by its sweetness, is exactly the same.70 Drosophila will prioritize food odor over the innately aversive odor CO2 when feeding on fermented fruit.14 Odor is, therefore, an integral part of food evaluation across multiple species, even to the extent of dominating multisensory inputs and spurring counter-intuitive behavior.

Advantages of odor sensation during exploitation. (A) Besides determining distance to food, odor plumes allow for distinctions between food type, quality and size and allow animals to navigate based on nutritional needs and other internal drivers. (B) Anticipatory feeding behaviors induced by food odor, including the motor- and digestive priming, allow for improved animal survival, fitness and reproduction through efficient food capture and ingestion
Because of its prominence and tractability as a genetic model organism, olfaction and food sensing has been extensively studied in C. elegans. In the worm, both the neural circuits as well as the mechanisms of odor-guided food selection are well understood. Worms display preferences for certain bacterial food sources over others28, 71, 72 and identify them based on their attractive odorants.73 When provided with bacteria found in their natural habitat, worms chose food with higher nutritional value, which then led to an extended lifespan.28, 71 Attractive smells further facilitate C. elegans feeding behavior in the presence of food via increased pharyngeal pumping, while repellent smells suppress it.74 In worms, the neurons primarily responsible for food sensing have been identified, as well as the navigational circuits that are active during foraging.9, 75 A neural “flip-flop” circuit has been proposed linking odor-sensory neurons AWC and AWB, as well as the neuropeptides they release, to the recognition and subsequent generation of food odor preference.76 When contradictory odors are encountered, this flip-flop motif performs a nonlinear computation of the sensory inputs that leads to stable behaviors in noisy environments where multiple conflicting or fluctuating odors are present. Moreover, AWC is part of the circuit that drives local food-searching behavior,9, 77-79 thus providing the neural circuit that connects odor sensation and the corresponding changes in locomotor behavior. C. elegans is therefore a highly useful model organism for studying the neural circuits linking odor and food exploitation.
Internal states such as hunger, mating drive or sleep drive similarly alter the perception of food value.68, 80-82 Internal states are often set by neuromodulators and neuropeptides, which signal gross changes in the balance between behavioral and sensory priorities.83, 84 For example, increased value can be allocated to odor sensation during times of hunger.85, 86 When placed on a single fly treadmill, hungry Drosophila will doggedly pursue a food odor even when no reward is forthcoming, meaning the promise of food driven by smell is enough to outweigh negative experiences.87 Furthermore, hungry Drosophila will pick up attractive, low odor concentrations of vinegar more acutely while reducing the neural activation caused by aversive high vinegar concentrations.88 Similarly, when C. elegans are starved they risk death to obtain a meal based on the food smell emanating across a dangerous desiccating hyperosmotic barrier.89 Odor sensation can thus affect food intake by altering the valuation of the food depending on current and dynamically changing organismal priorities.
3.2 Anticipatory digestive and motor behaviors lead to efficient feedingChanges in metabolism as a result of food odor detection can cause more efficient uses of resources by priming the body for digestion (Figure 2B). Efficient digestion leads to beneficial effects such as greater longevity and fertility, thereby positively impacting the animal's fitness.20, 22, 90 However, in dietary-restricted worms and flies, ablating odor-sensory neurons leads to an increase in life-span,91, 92 which in worms is mediated by an octopamine signal to the gut.93 While it is possible that this odor-mediated change in lifespan reflects a necessary tradeoff between earlier reproduction when conditions are favorable, and faster aging,94 the behavioral and ecological function of odor-mediated inhibition of longevity for the survival of the animals has not yet been established.
The cephalic phase response, or the preparation of the gastrointestinal tract for optimal food processing, has been described in rodents95-97 and humans, although the evidence from human trials is conflicting and debated.98-100 In model invertebrates, odors have been found to change lipid catabolism in peripheral fat storage tissues,20, 22, 90, 91 proteostasis21 and reproduction rates via germline proliferation.94 In C. elegans for example, the activation of odor-sensory neuron AWC by the food-related odor 2-butanone causes a cascade of metabolic reprogramming of fat-related pathways, leading to more efficient exploitation of energy sources without directly changing feeding behavior.20 Moreover, odor-regulated microRNAs inhibit AWC, stimulating proteostasis and prolonging longevity.21 The odor-sensory neuron AWB is responsible for detecting the presence of preferred dietary odors and adjusting germline proliferation accordingly, favoring increased reproduction rates and an early onset of reproductive aging to counteract increasing germline mutation rates with age.94 Other species, such as scavenging deep-sea amphipods, anticipate food intake by increasing their initial oxygen consumption when exposed to bait odor. The amphipods then switch from an energy-conserving state to an active one, allowing them to migrate quickly to the meal to exploit it and leave quickly to avoid predators.22 Digestive priming thus links odor sensing to exploitation by directly affecting and adapting metabolism to the available food sources (Figure 2B).
The second mechanism connecting odor sensation to increased food intake primes the animal by directly affecting the motor programs controlling biting or swallowing (Figure 2B). In C. elegans, the presence of attractive odors directly affects the feeding rate via the sensory flip-flop circuit described above74 and a similar effect can be observed in Drosophila larvae.101 Starved C. elegans upregulate their feeding rate even in the absence of food, possibly to enhance ingestion when food becomes available again.27 This upregulation might allow the animal to immediately detect even small food sources and exploit them without delay, as worms also employ feeding as a way of sampling their surroundings.102 By preemptively increasing the rate of food intake animals can reduce the time spent in the potentially dangerous and vulnerable state of feeding. Beyond these examples in model organisms, few experimental studies have shown direct evidence of motor priming, likely because of the difficulty in detecting food intake in unrestrained animals. Yet, motor priming will directly affect measured foraging parameters such as time spent in a food patch and will therefore need to be considered. Overall, olfaction can additionally improve food intake and resource utilization by inducing digestive or motor preparation.
4 MODELS OF FORAGING BEHAVIOR IN DIFFERENT DISCIPLINESAs a complex behavior, foraging encompasses many behavioral aspects that can be modeled, ranging from statistical models underlying the process of navigation, to inference of internal value models and the neural basis of decision-making. A typical foraging sequence comprises five distinct behavioral phases: random search (without cues), cued navigation toward a food source, encounter, feeding and leaving (Figure 3A). Here, we focus on models that aim to implicitly or explicitly codify the animal decisions that affect the timing and duration of foraging phases. At most transitions between foraging phases, animals can decide to enter the next phase, continue the current phase or repeat the previous phase (Figure 3A, black arrowheads). Thus, most foraging models fundamentally try to model animal decisions: habitat choice, patch choice, choice of diet and patch leaving are all decisions the foraging animal makes repeatedly.103, 104 Similarly, food choice, feeding duration in a patch and patch leaving are three decisions that have been tested against data for many foraging species.105

Foraging decisions are altered when a forager is primed by odor sensation. (A) Foraging encompasses multiple sub-behaviors of which a forager can perform all or a subset. The animal starts with a random search pattern, and once an odor is encountered, odor navigation begins. Upon food encounter, feeding begins. Eventually, the forager leaves the current food patch. There are many decisions involved (black arrowheads): choice of a patch, taste and choice of leaving. (B) Anticipatory behaviors can alter energy intake, for example by shortening ingestion and digestion delays or by allowing a higher metabolic efficiency. (C) Possible effects of odor priming on a marginal value theory forager: The forager leaves a patch when the food intake drops below the average expected rate (top, dashed line is the average expected rate, gray is time in a patch). Given the same time spent in a food patch, both efficient digestion (middle) and motor priming (bottom) lead to more energy intake compared with an unprimed classical forager, either by faster metabolism or reducing ingestion delays, respectively
In the classical paper by Charnov,106 the now well-known marginal value theorem (MVT) was introduced. Economical in nature, the theorem connects the travel time between patches of food with the optimal time to leave a patch, given that the forager aims to maximize the rate of food intake relative to the energy loss because of motion, that is, its energy balance. The theorem's value lies in clear predictions for measurable foraging parameters such as the optimal patch residence time. However, the original model has been criticized for lack of experimental support since data showed a consistent bias toward the forager remaining in the patch longer than predicted by the MVT.105 Subsequent modifications, therefore, included arbitrary food distributions,107 time-limitations for foraging because of other behaviors108 and underlying behavioral states.105
A different class of models from cognitive neuroscience use decision theory to explicitly describe the process of deciding, rather than its optimal economical outcome.105 While MVT assumes the forager has perfect knowledge of the available patches, decision theory models assume the forager has to decide based on unknown rewards, which it needs to measure during exploration of its environment.109 For example, an evidence-integrating drift–diffusion model has been used to predict feeding decisions in C. elegans when the concentration of food is uncertain.102
More complex hierarchical decision models have extended these basic choice models to integrate internal states such as hunger, mating drive or satiety and motivational states.110
Foraging can also be modeled by considering the information that is available to the animal for its decision. These approaches explicitly account for and consider how animals could optimally collect information from fluctuating or noisy cues. The proposed strategy for navigation using optimal information gathering was termed “infotaxis.”111 A noisy cue, such as a scent in a turbulent flow, could lead an animal to a food source. Infotaxis considers that the animal can either navigate toward the likely odor source or gather more information, depending on how accurate its current estimate of the source location is. The elegant algorithm is tractable for simple biological implementation. To date, however, there is limited experimental evidence for infotaxis, in part because of the difficulty in measuring information acquisition and processing in foraging animals. A study in C. elegans has shown some evidence that worms might implement infotaxis after being removed from a food source.78 For patch leaving, a model has now integrated noisy information about the environment and food rewards with evidence accumulation from decision theory.112 To date, models describing all aspects of foraging from search to patch leaving at a level of detail that includes cues, evidence and information, remain elusive.
5 INTEGRATING SMELL INTO MODELS OF FORAGING AND FUTURE EXPERIMENTAL DIRECTIONSCurrent foraging models account for odor cues when modeling exploration, or the navigation toward a food source. We suggest integrating anticipatory behaviors into existing foraging frameworks because of their impact on the dynamics of energy intake and changes to organismal risk assessment. To add the effects of priming into foraging models requires only small changes to existing models. We will here discuss how this adaptation could be achieved for the MVT model, as it is easy to illustrate how the effects of odor priming might affect the energy balance of the animal, which MVT aims to maximize.
Accounting for smell in the valuation of a food patch would mean adding an additional term to the cost function that describes food preference because of odor. Such preferences have been measured successfully for a large number of volatile and soluble compounds in binary or multi-choice assays.113-115 Therefore, adding this aspect would not necessarily require free parameters, but rather adding a term that is, sign and strength are already established (Table 1).
TABLE 1. Food-related odors and their behavioral effects on invertebrate animals Species Odor Natural source Behavioral effect Reference C. elegans Isoamylalcohol (IAA) Bacteria73 Increase in pirouettes 116 Diacetyl (DIA) Rotten fruit, lactic acid bacteria117 Attraction toward source smell 117 2-Butanone Pathogenic bacterium Serratia marcescens73 Attraction toward smell source 73 D. melanogaster larvae Ethyl butyrate Fruit, for example, physalis, cactus fig, honeymelon115 Attraction toward smell source 115 Ethyl acetate Fruit, for example, passionfruit115 Attraction toward smell source 115 Methyl butyrate Byproduct of microorganisms in soil and rotten vegetation118 Increased in pirouettes 119 D. melanogaster adults Mthyl acetate Fruit, for example, passionfruit115 Increase in walking speed 120 Benzaldehyde Almonds, fruits121
留言 (0)