
記住我

Drosophila melanogaster, a model organism that plays an illustrious role in many branches of biological research, has contributed greatly to our understanding of complex pathological processes, including neurodegeneration, obesity, diabetes, and aging related diseases (Baker & Thummel, 2007; Demontis et al., 2013; Hirth, 2010). In cancer research, the Drosophila model has been crucial to the discovery of genes and pathways that play oncogenic or tumor suppressor roles, thanks to their shared functions in the development of multicellular organisms (Brumby & Richardson, 2005; Parvy et al., 2018; Sonoshita & Cagan, 2017; Stratton, 2011). One such gene is lethal (2) giant larvae (lgl), first discovered in the Drosophila model (Gateff, 1978), and later found to be part of a group of cell-polarity genes that also include scribble (scrib) and discs large (dlg). Mutations of these genes, collectively known as neoplastic tumor suppressor genes, lead to loss of apicobasal polarity in epithelial cells and neoplastic overgrowth in the larval brain and imaginal discs (Bilder et al., 2000a; Humbert et al., 2008). The human scrib homolog was later found to assume a tumor suppressor role in breast, liver, skin, and lung cancers (Elsum et al., 2012; Moreno-Bueno et al., 2008), highlighting the evolutionarily conserved function of this class of genes in tumor suppression.
In recent years, owing to the large arsenal of powerful and versatile tools available for genetic manipulation, a growing number of tumor models have been established in various Drosophila tissues to address fundamental questions of cancer biology. Among these tools, the flippase recombinase (FLP)/flippase recognition targets (FRT) based mitotic recombination technique, which can generate mosaics that induce tumorigenic mutations in clones, is particularly useful, as it resembles the process of cancer initiation (Potter et al., 2000). When this technique is combined with other tools such as the Gal4/Upstream Activation Sequence (UAS), RNA interference (RNAi), and Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/Cas9 techniques, cancer-related genes can be manipulated in a strict spatial- and temporal-pattern for exploring mechanisms in tumor initiation, growth, metastasis, and cell–cell interactions within a tumor microenvironment (Chatterjee & Deng, 2019; Gonzalez, 2013; Herranz et al., 2016; Johnson & Halder, 2014; Sonoshita & Cagan, 2017; Stefanatos & Vidal, 2011). In this review, we focus on tumor models in various Drosophila tissues with human cancer hallmarks (Figure 1) (Hanahan & Weinberg, 2011), and explore consequences of tumor growth, including cachexia-like wasting and tumor–host interactions. Specific attention will be given to tumor models that are capable of transplanted growth and metastasis (Figures 2 and 3).

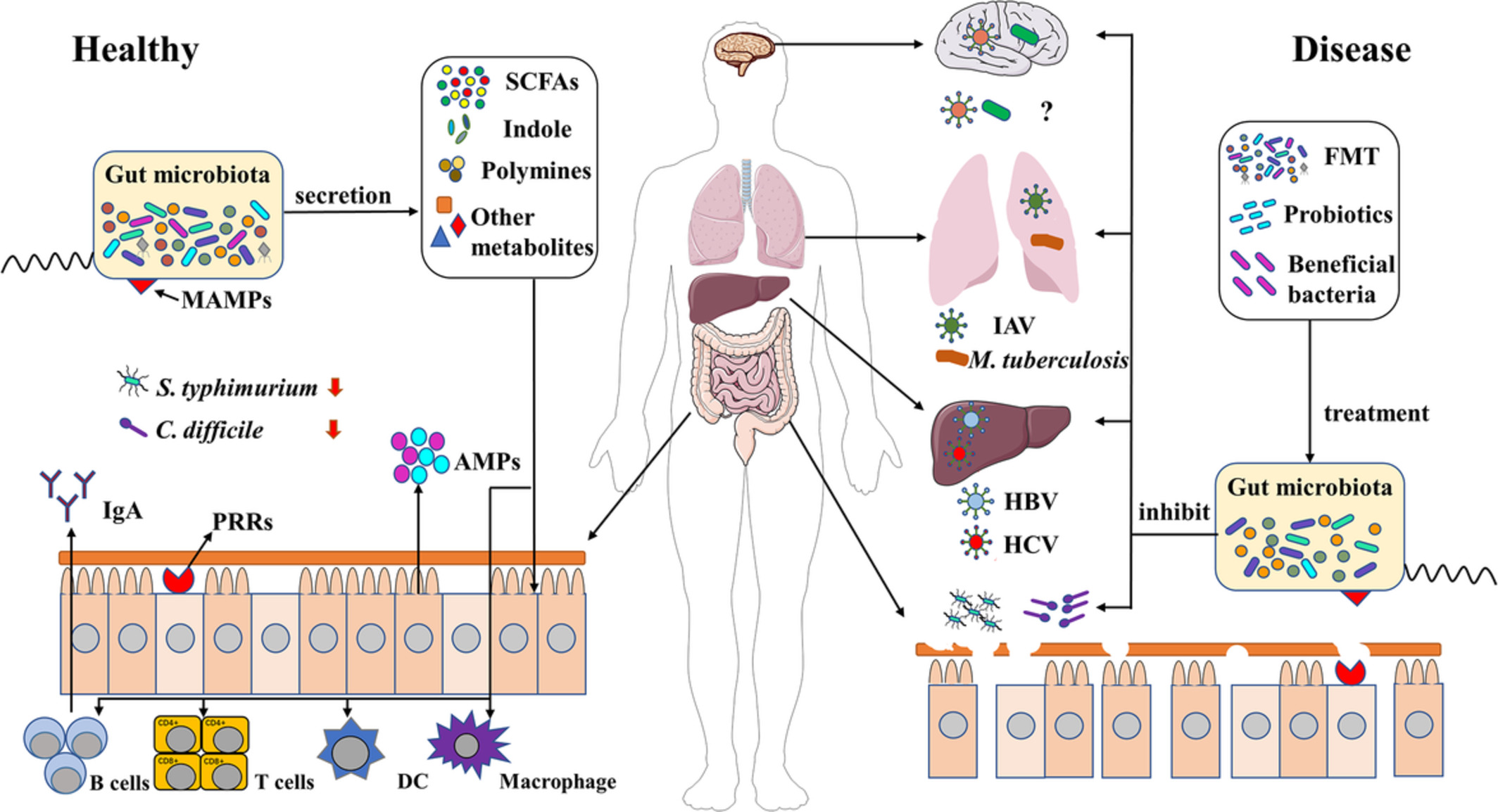
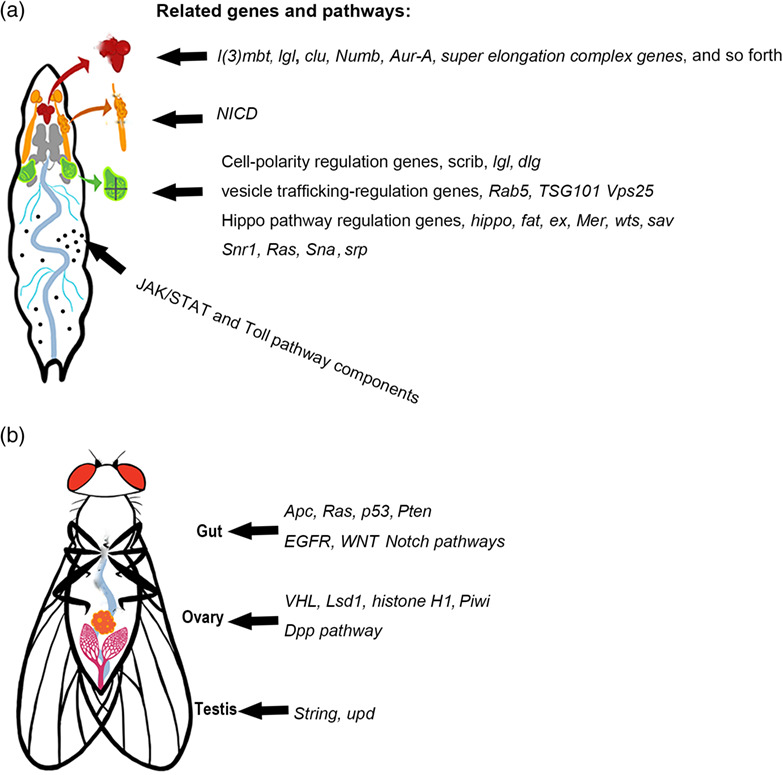
Tumor models and related genes and pathways in different larval and adult tissues. (a) Larval tumor models in the brain, salivary gland imaginal ring, imaginal discs and hemocytes. (b) Adult tumor models in the gut, ovary, and testis

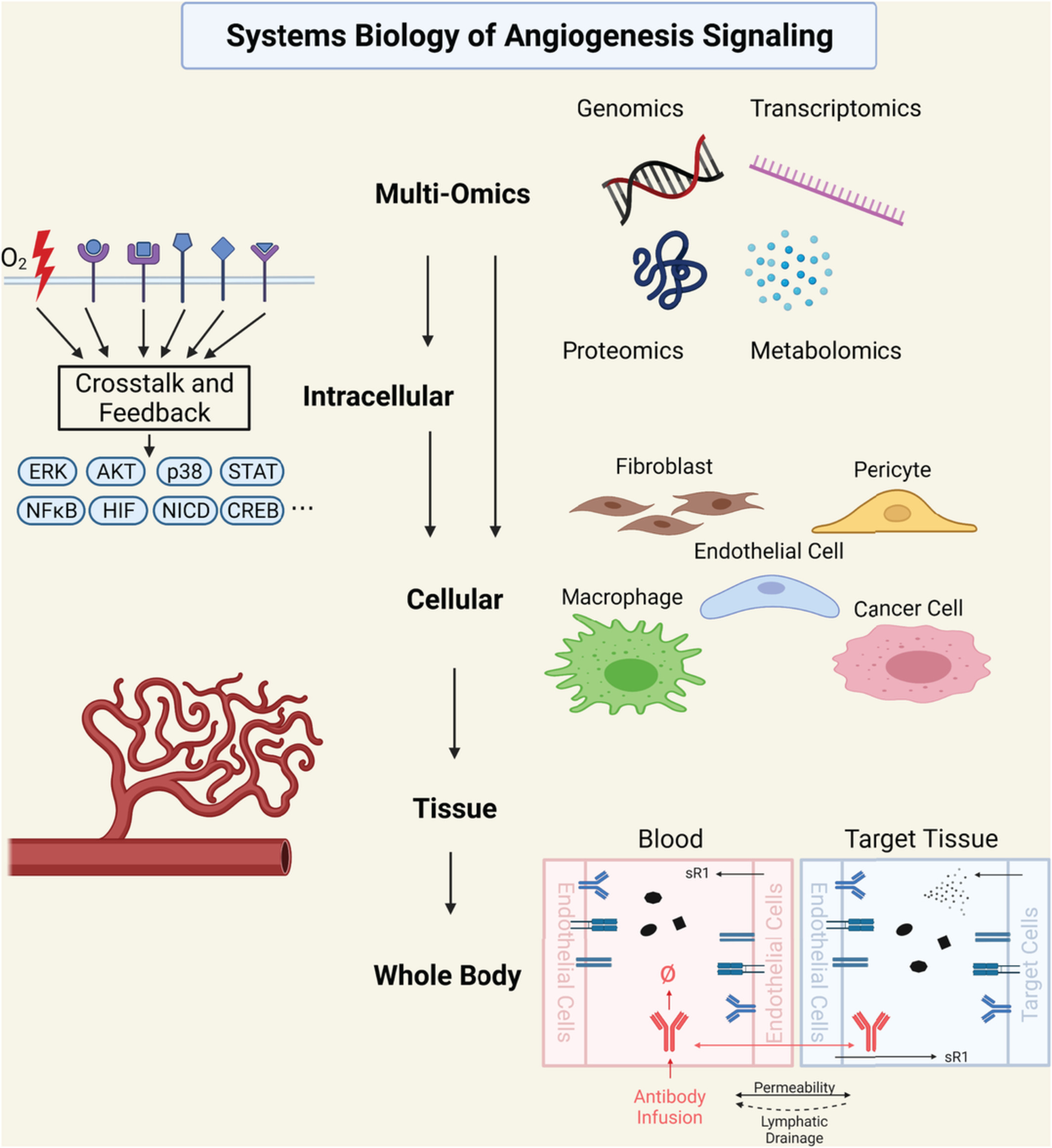
Tumor transplantation (xenograt) studies in Drosophila. First, primary tumors are induced in various tissues. They are then harvested and sectioned into small pieces. Next, the tumor pieces are injected into the abdomen of a host adult fly. The transplanted tumor can be harvested and replanted into new host flies for continued tumor growth

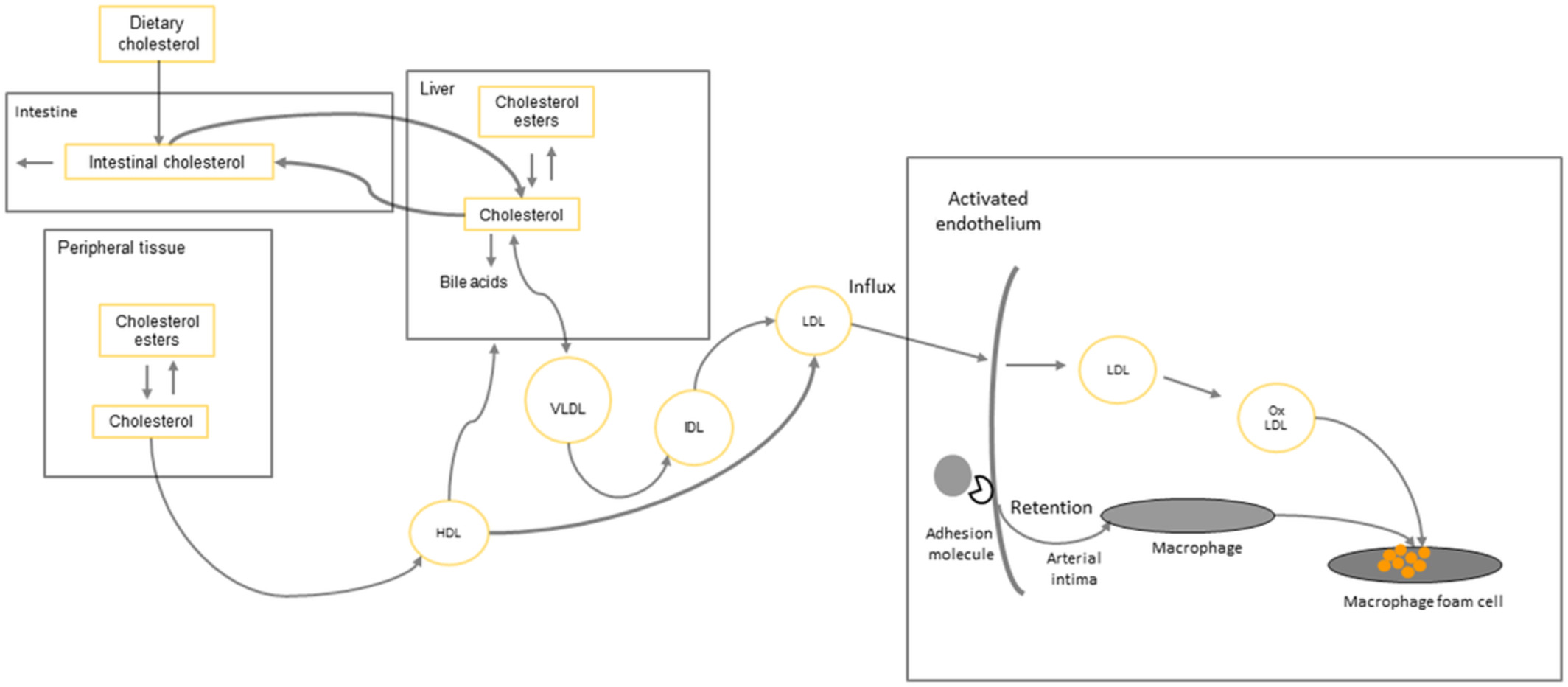
Metastasis in Drosophila tumor models. (a) The larval tumor cells metastasize to the gut, fat body, and trachea. (b) Tumor cells from the transplanted tumor metastasize to the gut, ovary, muscle, and brain. (c) The ISC tumor cells induced in the adult gut metastasize to the ovary and other tissues. (Arrows indicate metastasis directions)
2 TUMOR MODELS IN DIFFERENT TISSUES 2.1 The imaginal tissue 2.1.1 Imaginal discsThe imaginal discs, which are structured epithelial sacs found in the larvae, have been popular tissues for tumorigenesis studies in Drosophila (Elsum et al., 2012; Herranz et al., 2016; Mirzoyan et al., 2019). During metamorphosis, these discs undergo major morphogenetic changes precipitated by intrinsic hormonal pathways and transform into exoskeletal structures including the head, thorax, and appendages (Blair, 2009). In total, there are 19 discs, each of which is formed from precursor cells that originate in the ectoderm of the embryo. Disc cells have a high rate of proliferation in the wondering larval stage, and the largest disc the wing disc can grow to a final size of approximately 50,000 cells (Aldaz & Escudero, 2010; Johnston et al., 1999).
The disc epithelial cells exhibit the archetypal apical-basal polarity. Their growth and proliferation are regulated by conserved signaling pathways. Deregulation of the growth signaling and disruption of cell polarity are two major causes leading to tumor formation in Drosophila epithelial tissues. The cell-polarity genes scrib, lgl, and dlg and their associated neoplastic growth phenotypes have been well characterized using the disc models (Herranz et al., 2016; Humbert et al., 2008). Another class of genes that may lead to neoplastic overgrowths when mutated in imaginal discs include regulators of vesicle and membrane trafficking, for example, Tumor susceptibility gene 101 (TSG101), Vacuolar protein sorting 25 (Vps25), and Rab5. Not surprisingly, their mutants exhibit simultaneous cell-polarity loss and activation of growth-promoting signaling pathways (Gilbert et al., 2009; Herz et al., 2006; Thomas & Strutt, 2014). In this section, we focus mainly on recent developments using the imaginal-disc models to understand how different regulatory factors contribute to tumorigenesis in Drosophila.
The disc model has been used to determine the relationship between oncogenic mutations and tissue microenvironment during tumor initiation. It has long been known that oncogenic or tumor suppressor mutations do not always lead to neoplasms. For example, the Rb mutation, while commonly detected in many tissues, leads to tumorigenesis exclusively in the eye and bone (Ferres-Marco et al., 2006). With the advent of high throughput sequencing technology, cancer driving mutations have been found to be a lot more prevalent in healthy individuals than previously thought, raising questions about the exact role of these oncogenic mutations in cancer initiation (Martincorena et al., 2018). In this regard, the Drosophila wing disc model elucidates the role of tissue microenvironment in determining whether cells bearing oncogenic mutations can develop into a tumor. Applying mosaic generating tools, loss of lgl or scrib in different regions of the wing disc resulted in different consequences on tumorigenesis. Tumors grow specifically in the hinge region but not the pouch region of the wing disc (Tamori et al., 2016). A key difference between these tissue microenvironments is that JAK/STAT signaling is more potent in the hinge region than in the pouch. Another difference is the organization of the epithelium, which translates to discrepancies in the extrusion direction of pro-tumor cells. These findings have led to the concept of tissue tumor hotspot that describes the role of microenvironment in tumorigenesis (Tamori & Deng, 2017).
Similar to the wound healing process in humans, tissue regeneration in Drosophila involves genes and pathways that also regulate cell proliferation and growth. One such pathway is the Salvador-Warts-Hippo axis (Grusche et al., 2011), which is crucial in organ size control (Zhao et al., 2011). Yorkie (Yki), the Drosophila version of Yes-associated ptorin 1 (Yap), is a key transcriptional co-activator that serves a pivtal role in this axis (Sun & Irvine, 2011). By receiving upstream signals from atypical cadherin (Daschsoous and Fat) and myosin, Yki becomes the rate-limiting activator in Drosophila tissue regeneration. Yki is activated by wing disc damages via the Jun N-terminal kinase (JNK) pathway (Sun & Irvine, 2011). Thus, it is not surprising that a disregulated Yki can result in massive compensatory proliferation and organomegaly. It turns out that certain tumors can take advantage of this pathway during their dysplastic changes (Katsukawa et al., 2018). These studies help define the relationship between tissue regeneration and tumorigenesis.
The disc model has also been used to determine how genes involved in the epidermal–mesenchymal transition (EMT) promote tumor formation, similar to their human counterparts (Moreno-Bueno et al., 2008). snail (sna) and serpent (srp) are two EMT inducers in Drosophila. srp overexpression in wing discs leads to neoplastic transformation. The derived tumor grows many times larger after transplantation into a new host fly, and can continue to grow for many generations following repeated transplantation (Campbell et al., 2018). The underlying mechanism of srp-overexpression induced tumorigenesis involves activation of both Yki and Ras signaling. In contrast, overexpression of sna does not induce neoplastic growth, because sna activates only Yki but not Ras in this tissue (Campbell et al., 2018), suggesting that neoplastic growth involves multifaceted integration of signaling factors.
Epigenetic regulation is crucial in developmental plasticity and has been shown increased importance in tumorigenesis (Gonda & Ramsay, 2015). Most epigenetic regulators are evolutionarily conserved and are commonly mutated in human cancers (Piunti & Shilatifard, 2016). In Drosophila, the Polycomb group (PcG) and the Trithorax group (TrxG) chromatin proteins are major epigenetic regulators that function complementarily to maintain the repressed and active gene expression states, respectively (Geisler & Paro, 2015). A mutation of Polyhomeotic (Ph), a conserved member of the Polycomb repressive complex 1 (PRC1) in eye-disc cells, results in neoplastic growth (Y. R. Jiang et al., 2018). Loss of Ph leads to activation of knirps (kni), an orphan nuclear hormone receptor and embryonic transcription factor, and reprogramming of cellular identity toward an embryonic state. Overexpression of kni could also lead to tumor formation in the larval eye disc. Both loss of Ph and kni overexpression share features such as ectopic activation of the JAK/STAT pathway and a block of differentiation (Torres et al., 2018).
In addition, PcG-mediated histone modifications regulate chromatin conformation within the irradiation-responsive enhancer region (IRER), an intergenic sequence that mediates the expression of surrounding proapoptotic genes, reaper, hid, and grim following ionizing irradiation (Y. Zhang et al., 2008). dMyc-induced apoptosis or hyperplasia is dependent on the epigenetic status of IRER. In flies with IRER deleted or in a closed epigenetic status, dMyc-overexpression induced apoptosis is blocked, whereas hyperplastic growth takes place, suggesting the important role of the epigenetic status of IRER in cellular decisions following oncogene-induced stress (C. Zhang et al., 2015).
Epigenetic regulators also include chromatin remodelers such as the Switch/sucrose nonfermentable (SWI/SNF) chromatin-remodeling complex, including brahma (brm) or Snf5-related 1 (Snr1). The brm and Snr1 homologs are frequently mutated in human cancers (Reisman et al., 2009). Knockdown of brm or Snr1 along with simultaneous overexpression of Yki using apterous-Gal4 (apGal4) leads to tumor growth in the dorsal compartment of the wing disc. SWI/SNF performs functions similarly to those of the Hippo pathway in suppressing Yki activity, which can induce tumor-like growth (S. L. Song et al., 2017). In a separate study, Snr1 was shown to prevent tumorigenesis by maintaining an endosomal trafficking-mediated signaling cascade (Xie et al., 2017). Removal of Snr1 resulted in neoplastic overgrowth in imaginal discs, whereas the depletion of any other members of the SWI/SNF complex failed to induce similar phenotypes. These findings are consistent with the fact that SMARCB1, the Snr1 homolog in humans, is the only gene of the SWI/SNF complex responsible for malignant rhabdoid tumors (MRTs). It appears that the aberrant regulation of multiple signaling pathways, including Notch, JNK, and JAK/STAT, is responsible for tumor progression upon Snr1 depletion. The tumor suppressor function of Snr1 is largely dependent on its subcellular localization to the cytoplasm, rather than to the nucleus (Xie et al., 2017). Because MRTs are not of epithelial origin, the modeling of these pediatric tumors in Drosophila imaginal discs suggests the versatility of this system in understanding basic functions of tumor suppressor genes in normal development and their potential effects on tumorigenesis.
2.1.2 Imaginal ringsThe imaginal ring is another imaginal tissue precursory to adult organogenesis in insects. Unlike the imaginal discs that form the exoskeletal structures in the adult, the imaginal rings eventually develop into the adult gastrointestinal organs, including the foregut (Fuss et al., 2004), hindgut (Tian et al., 2016), and salivary glands (Haberman et al., 2003). Notch signaling is activated in all three imaginal rings from the middle embryonic stage to the early pupal stage, and positively controls imaginal-ring cell proliferation during the third larval instar (Haberman et al., 2003; S. A. Yang & Deng, 2018). Overexpression of an active form of Notch, the Notch intracellular domain (NICD), induces overproliferation in these imaginal rings, similar to the hyperplasia growth phenotype induced by NICD in many imaginal discs (Djiane et al., 2013; S. A. Yang & Deng, 2018).
Interestingly, continuous overexpression of NICD in the salivary gland imaginal ring triggered tumor formation (S. A. Yang et al., 2019). These tumors form at the posterior end of the imaginal ring, which borders the polyploid giant-salivary-gland cells. This boundary region resembles the so-called “transition zones” known for their susceptibility to metaplasia, a premalignant lesion in mammalian epithelial tissues (Mcnairn & Guasch, 2011). Notch hyperactivation is sufficient to induce tumorigenesis at the salivary gland transition zone, but not in other imaginal ring cells. The underlying mechanism for tumor induction in this specific location is the endogenous high-level of JAK/STAT signaling and Matrix Metalloprotease 1 (MMP1) activity, induced by the JNK pathway. These tumors grow steadily in the host abdomen after transplantation and can reach the size about 30-times of the original tumor. Some tumor cells disseminate from the transplanted tumor and migrate to the host ovarian muscle sheath, oviduct, gut, and Malpighian tubule (S. A. Yang et al., 2019). This tumor model offers an opportunity to explore the effect of tissue microenvironment on tumor initiation and how host–tumor interactions affect tumor growth and metastasis.
2.2 The central nervous systemThe Drosophila central nervous system (CNS) consists of neurons organized into lineages, each of which descending from a single neuroblast specialized from a progenitor cell (Spindler & Hartenstein, 2010). The first round of neuronal divisions is performed by about 100 primordial neuroblasts, derived from the ectodermal layer around embryonic stage 10 (Younossi-Hartenstein et al., 1996). The neuroblast undergoes asymmetric division to produce a daughter neuroblast and a ganglion mother cell (GMC); the latter then generates two individual neurons via an additional division (Urbach & Technau, 2003). Although all cells in a proneural cluster have the potential, only one cell in each cluster is designated to become a neuroblast in CNS.
Cell fate specifications in the neural cell lineage involve cell–cell signaling mediated by the Notch pathway. The neuroblast expresses high levels of the Notch ligand Delta, thus activating Notch in the surrounding cells, inhibiting their differentiation into neuroblasts. In the larval stage, the neuroblast undergoes stereotypic, self-renewing divisions to produce INPs (immature neural progenitors), which, upon maturation, undergo a few more rounds of asymmetric, self-renewing divisions to give rise to ganglion mother cells (GMCs) that subsequently generate postmitotic neurons or glia (Bello et al., 2008). The disruption of cell-polarity and Notch pathway genes can potentially lead to tumorigenesis in the larval brain (Froldi et al., 2019; Goh et al., 2013; Y. Song & Lu, 2011).
Unidirectional Notch signaling that involves direct cell–cell contact between signal-sending and signal-receiving cells is both necessary and sufficient to promote neuroblast self-renewal (Bowman et al., 2008). At each neuroblast division, differentiation-promoting determinants are asymmetrically segregated. For example, the Notch antagonist Numb shows high levels of expression in immature INPs to suppress Notch signaling. Thus, the Notch pathway effector HES (hairy and E[spl]) genes are highly expressed in neuroblasts but not in immature INPs (Zacharioudaki et al., 2012). numb mutant INPs fail to complete maturation but instead revert back into the neuroblast fate and result in tumorigenesis in the larval brain (H. Wang et al., 2006).
Hyperactivation of Notch induced by changes in different Notch regulatory mechanisms leads to immature INP dedifferentiation and tumorigenesis, that is, the overproliferation of neuroblasts. One such regulation is achieved by the super elongation complex (SEC) which directly interacts with the Notch transcription activation complex, including dCSL (Drosophila homolog of CSL, also named Suppressor of Hairless (Su(H)). dCSL interacts tightly with the SEC subunit AF4/FMR2 family-3 (dAFF-3). The inactivation of SEC leads to neuroblast loss, whereas its forced activation results in neural progenitor dedifferentiation and tumorigenesis. Overexpression of the SEC subunits such as eleven-nineteen lysine-rich leukemia (dELL) and dAFF induces the expression of hairy and E(spl)) genes (dHES), leading to a progenitor-derived tumor. These larval brain tumors can be sectioned and transplanted into the abdomen of young female adult hosts (Figure 2). Notably, only the tumor cells co-expressing dELL and dAFF or dELL, dAFF and dCSL can grow into large tumor masses and metastasize to the eye and thorax after xenograft of the tumor tissue into the adult abdomen. In contrast, the larval brain tumor tissue derived from dCSL-overexpression alone fails to metastasize in the adult host abdomen (Liu et al., 2017).
The cell polarity genes lgl and atypical Protein Kinase C (aPKC) have also been investigated extensively for their roles in neuroblast tumorigenesis (Homem & Knoblich, 2012). This is mainly due to their involvement in neuroblast asymmetric cell division, which often gives rise to an identical neuroblast as well as a differentiated daughter cell. This process begins with polarity establishment within the neuroectoderm during the embryonic stage, which is accomplished by the Bazooka (Baz) protein. The apically localized Baz directs the assembly of multiple downstream scaffolding complexes, which then coordinate enzymatic activities involved in asymmetric divisions (Gómez-López et al., 2014). Next, the mitotic spindle is oriented around the cellular scaffolds by the mushroom body-defective (Mud) protein. (Izumi et al., 2006). The final step in asymmetric division is cell fate determination, an intricate process directed by the Baz/Par6/aPKC complex and dependent on actin and myosin functions (Kraut et al., 1996).
Disruption of normally precisely controlled aPKC activity in neuroblasts can trigger tumor formation in larval brain. The aPKC/Baz/Par6 complex phosphorylates Numb and Miranda (Mira) to exclude them from the apical cortex, establishing the basal localization of cell fate determinants (C. Y. Lee et al., 2006). In lgl null mutants, Numb and Mira localization is defective, which leads to neuroblast overproliferation due to dysfunctional spindle formation (Atwood & Prehoda, 2009). The Drosophila ortholog of clustered mitochondria homolog (CLUH), clueless (clu), has been found to promote lgl-induced tumorigenesis in the larval brain through regulating aPKC activity (Goh et al., 2013). Clu preferentially binds to the aPKC/Bazooka/Par6 complex and stabilizes aPKC levels.
A Drosophila larval CNS tumor model induced by the lethal (3) malignant brain tumor (l(3)mbt) mutation revealed the role of germline genes on tumorigenesis (Janic et al., 2010). The germline genes such as nanos (nos), vasa (vas), P-element induced wimpy testis (piwi), and aubergine (aub) are upregulated in these tumor cells to induce a soma-to-germline transformation, which then help the larval brain tumor cells undergo self-renewal and hyperproliferation. Inactivation of these germline genes successfully suppressed tumor growth. Furthermore, blocking piwi, vas or aub in combination with the l(3)mbt mutation inhibited the transplanted tumor growth. It has been shown in various human malignancies, including melanoma and several types of carcinomas, that germline genes including Melanoma antigen-1 (MAGEA1), MAGE-A2, ADAM metallopeptidase domain 2 (ADAM2), and cancer/testis antigen 1B (CTAG1B) become aberrantly activated (Akers et al., 2010). These genes, suspected to contribute to oncogenic traits such as immortality, invasiveness, and hypomethylation (Simpson et al., 2005), are being pursued as biomarkers and as targets for therapeutic cancer vaccines (Old, 2008). The similarities between the fly model and human cancers in germline gene upregulation present a promising new direction for future Drosophila cancer research (Gonzalez, 2013).
2.3 The adult intestinal systemThe Drosophila adult intestine is comprised of an ectoderm-derived foregut and hindgut, and an endoderm-derived midgut. The adult midgut is the functional equivalent of the mammalian small intestine where food digestion and nutrient absorption occur. The midgut epithelium is monolayered and contains self-renewing intestinal stem cells (ISC), the only dividing cell type in the midgut. After asymmetric division, an ISC produces a renewed ISC and either an enteroblast (EB) or a pre-enteroendocrine cell. EBs originate from the mitotic cell cycle and differentiate into absorptive enterocytes (ECs). Pre-enteroendocrine cells differentiate into mature enteroendocrine cells (EEs) (Miguel-Aliaga et al., 2018). This feature of a single-layer epithelium with divisional capabilities limited to ISCs makes the adult midgut an ideal model to explore tumorigenesis from stem cells (Singh et al., 2019).
In normal development, ISC division and differentiation are regulated by a combination of signaling pathways (H. Jiang et al., 2016). Their deregulation could lead to ISC overproliferation or loss of differentiation, which are major contributors to tissue overgrowth and tumor formation in the midgut model. Loss of Notch (N) has been shown to block differentiation, thus inducing an accumulation of ISCs (Micchelli & Perrimon, 2006; Ohlstein & Spradling, 2006). Along with loss of N, stress-induced divisions and an autocrine EGFR ligand Spitz are independently required for tumor initiation and early tumor growth in the ISC model (Patel et al., 2015). The stress-dependent cytokines Upd2 and Upd3 are induced to propel tumor growth without secondary mutations. In addition, APC mutation-driven intestinal adenoma has been shown to compete with surrounding cells by inducing apoptosis in the latter (Suijkerbuijk et al., 2016). Competition between the tumor cells and the surrounding enterocytes is mediated by JNK and Yki. APC−/− mutant cells upregulate both JNK signaling and Yorkie activity, which in turn promotes tumor growth. The ISC model can thus be used to explore the relationship between tumor and the host microenvironment.
The Drosophila intestine has been used to model colorectal cancer (CRC), a multifactorial neoplasm involving genetic, environmental, and lifestyle risk factors (Brenner et al., 2014; Rawla et al., 2019). In humans, 40% of all cases of CRC involve mutations in the KRAS gene, while 70% show loss of function of APC. The combination of APC loss and KRAS mutation have been reported to cause an increase in adenoma size, number, and invasiveness (Cordero & Sansom, 2012; Janssens et al., 2006). In Drosophila, the activation of Wg/Wnt by loss of APC or the activation of Ras signaling in the midgut promotes ISC proliferation and causes tissue hyperplasia (Biteau & Jasper, 2011; H. Jiang et al., 2011; W. C. Lee et al., 2009; Martorell et al., 2014; C. Wang et al., 2013). Importantly, the combination of APC loss and Ras activation in the ISC model generates tumors with hallmarks of the human CRC (Martorell et al., 2014; C. Wang et al., 2013). These APC-Ras intestinal tumors are normally confined to the midgut, but the expression of the epithelial–mesenchymal transition (EMT) master inducer snail (sna) leads to the dissemination of tumor cells and macrometastases, which show parallels to human metastases (Campbell et al., 2019). sna activates a partial EMT in tumor cells, which then undergo collective cell migration and seed polyclonal metastases in multiple distant locations, including the ovary, thorax, and head, providing an excellent model for exploring metastatic behavior.
The adult hindgut, which has also been used to model colon cancer, is composed of large cuboidal epithelial cells confined to an anterior narrow segment called the hindgut proliferation zone (HPZ). Within the HPZ, self-renewal of ISCs, as well as subsequent proliferation and differentiation, are controlled by Wingless and Hedgehog (Hh) (Takashima et al., 2008). In the hindgut model, introducing a combination of multiple genes, including RasG12V and knockdowns of p53, APC, Pten, and dSmad4, can recapitulate key multigenic features of human colon cancer (Bangi et al., 2016). These features include the proliferation, disruption of the epithelial architecture, evasion of apoptosis and senescence, epithelial–mesenchymal transition (EMT), migration and cell dissemination to distant sites. Utilizing this multigenic model, the Drosophila hindgut has been used as a platform to investigate drug–tumor interaction and to screen various antineoplastic medications.
2.4 The reproductive tissue 2.4.1 Ovarian germline stem cellsWithin the female reproductive system, cells that have been reported to undergo tumorigenesis include the germline stem cells (GSCs) and the somatic follicle cells in the adult ovary. The ovaries are made of ovarioles, tube-like structures consisting of developing egg chambers and the anteriorly located germarium, where the GSCs are located. Asymmetric division of the GSC generates a daughter stem cell that retains contact with the somatic cap cells. The posterior daughter cell, however, is dissociated from the cap cells to differentiate into a cystoblast, which continues to undergo incomplete cytokinesis to form a 16-cell cyst. One of the cyst cells further differentiates into an oocyte, whereas the remaining 15 cells become polyploid nurse cells. The egg chamber, the developmental unit of oogenesis, is formed as the somatically derived follicle cells migrate to wrap the 16-cell cyst in the germarium (Greenspan et al., 2015; Hudson & Cooley, 2014).
Ovarian germline tumors in Drosophila are usually formed when GSC differentiation is blocked and continued divisions occur. Bag-of-Marbles (Bam), a member of a deubiquitinase complex to stabilize Cyclin A, is a key differentiation-promoting protein for cell-fate switching from a self-renewing stem cell to a differentiation-competent cystoblast. Bam is normally expressed in cystoblasts. Premature expression of Bam leads to GSC loss, whereas its suppression results in continued divisions of the GSC, producing GSC tumors. Disruption of bam, which can be achieved by the loss of Bam interaction partners in the cystoblast, is responsible for the majority of GSC tumor phenotypes reported so far (Iovino et al., 2009; Ohlstein & McKearin, 1997). Benign gonial cell neoplasm (Bgcn), Sex-lethal (Sxl), Mei-P26, and Twin are all factors related to RNA processing of Bam. Individual loss of any one of these in the female germline gives rise to the GSC tumor (Chau et al., 2009; Fu et al., 2015; Li et al., 2013; Ohlstein et al., 2000).
Bam expression in the cystoblast requires effective cell–cell communications elicited by the bone morphogenetic proteins (BMPs) from the somatic cells in the germarium. The BMP ligands Decapentaplegic (Dpp) and Glass-bottom boat (Gbb) are secreted from the somatic cap cells and target the GSCs via receptors Thickveins (Tkv), Saxophone (Sax), and Punt. This ligand-receptor interaction leads to phosphorylation of Mothers against Dpp (Mad), and its translocation into the GSC nucleus, where it associates with the bam promoter to suppress its transcription (L. Luo et al., 2015). In regions where BMP signaling is absent, Bam is expressed to facilitate germline differentiation. Many mutations that result in ovarian germline tumor formation, for example tkv, fused, smurf, and mir-184, are directly or indirectly involved in regulating BMP signaling in the GSC. Fused is a serine/threonine kinase that forms a complex with Smurf to control the fate of GSCs by generating a gradient BMP response (Xia et al., 2010). Mutations of microRNA, miR-184, which inhibits translation of the Sax receptor in wild-type cystoblasts, can also lead to GSC tumor formation (Iovino et al., 2009).
Maintenance of the GSC homeostasis requires the cooperation of different somatic cell types in the stem cell niche. An interesting example on how disruption of interactions among different somatic cell groups in the germarium leads to germline tumors involves the histone lysine-specific demethylase 1 (Lsd1) null mutation (Eliazer et al., 2011; H. Zhang & Cai, 2020). Lsd1 functions specifically within the escort cells to prevent adjacent cap cells from ectopically producing niche-specific BMPs. Loss of Lsd1 results in increased BMP signaling, which represses bam expression in the germline and leads to GSC tumor formation. Chromatin immunoprecipitation coupled with massive parallel sequencing (ChIP-seq) revealed over 100 genomic loci that are potential Lsd1 binding sites, including engrailed, which encodes a homeobox transcription factor (Eliazer et al., 2014). Knockdown of engrailed alleviated the stem-cell-expansion phenotype observed in Lsd1 mutants. Interestingly, Knockdown of other potential direct Lsd1 targets that are unlikely BMP signaling components, also partially suppresses the Lsd1 mutant phenotype. These results suggest that epigenetic regulator Lsd1 restricts the number of GSC-like cells by targeting a diverse group of genes to control the interactions among different cell groups in the germarium (Eliazer et al., 2014).
Similarly, depletion of histone H1 in escort cells also leads to abnormal BMP signaling in germline cells and tumorigenesis. Knockdown of H1 allows escort cells to acquire the ability similar to the cap cells to produce Dpp, resulting in upregulated BMP activity to decrease Bam expression in spectrosome-containing germline cells (SCCs). These overproliferative SCCs are the result of failed GSC differentiation into cystoblasts, probably taking on an intermediate fate. Interestingly, transposon activity increased dramatically in these H1-depleted escort cells (Z. H. Yang et al., 2017). Not surprisingly, loss of Piwi, a transposon silencer in escort cells, also resulted in GSC-like tumors in ovaries throughout adult life. The underlying mechanism appears to be similar to H1 knockdown in escort cells. Piwi-loss leads to Dpp upregulation in escort cells, which upregulates BMP signaling in the germline cells and blocks the differentiation of the GSCs (Z. G. Jin et al., 2013). Together, these studies suggest that abnormal epigenetic regulation of signaling events at the stem cell niche could lead to stem-cell derived tumorigenesis.
The control of Bam expression is also subject to regulation by sex determination genes. Maintenance of the sexual identity of the female germline has been shown to be crucial in preventing germline tumor formation. Differentiation from the GSC to cystoblasts requires physical interaction between Bam and Sex lethal (Sxl), the master regulator in the sex determination cascade. Loss of Sxl leads to a global upregulation of testis genes and a GSC tumor phenotype (Chau et al., 2009; Shapiro-Kulnane et al., 2015). The upregulated testis gene, PHD finger 7 (Phf7), is a male germline sexual identity gene and a key effector of the tumor-forming pathway. Phf7 upregulation is both necessary and sufficient for GSC tumor formation. In the female germline tumors with a loss of Bam expression, Phf7 is also upregulated, suggesting the critical role of sexual identify maintenance in the GSC tumor formation (Shapiro-Kulnane et al., 2015). Another Sxl related GSC tumor model involves an allele of sans fille (snf), which specifically eliminates the Sxl protein in germline cells. The majority of snf tumor GSCs show testis-enriched markers and exhibit an intermediate fate between the stem cells and their differentiated daughter cells (Chau et al., 2009). These tumor cells also contain spectrosomes, similar to the ones produced when histone H1 is depleted in escort cells, suggesting the generation of the GSC- or GSC-like tumor cells converge on failed differentiation and adoption of an intermediate fate.
2.4.2 Somatic follicular cellsTumors can be formed in somatically derived follicular epithelial cells in the ovarian tissue. These cells maintain a classic apical-basal polarity similar to epithelial cells in other tissues. Mutations in genes that show disruption of epithelial polarity (e.g., lgl, srib, and dlg) or endocytic membrane trafficking (e.g., Vps, tsg101) result in formation of multilayers of follicle cells (Bilder et al., 2000b; Tanentzapf et al., 2000; Vaccari & Bilder, 2005). Interestingly, this tumor-like multilayering is most prominent at the two termini of the egg chamber. Not coincidently, JAK–STAT signaling is strong in this terminal region, similar to the “tumor hotspots” in the hinge region of the wing imaginal disc and at the transition zone of the salivary gland imaginal ring (Beccari et al., 2002; McGregor et al., 2002). The precise role of JAK–STAT signaling in tumor formation at the egg chamber termini, however, requires further clarification.
The Notch pathway plays crucial roles in follicle cell differentiation and growth during multiple stages of oogenesis (Klusza & Deng, 2011). The role of Notch in the follicle cells is paradoxically opposite to its role in imaginal tissues. Notch activation triggers a transition from the mitotic cycle to the endocycle in follicle cells (Deng et al., 2001; Lopez-Schier & St Johnston, 2001). Disruption of Notch signaling leads to continued cell division, suggesting a tumor suppressor role of Notch in the follicle cells. The involvement of Notch in tumorigenesis in different Drosophila tissues is thus complex, ranging from an oncogenic role in imaginal tissues and neuroblasts and a tumor suppressor role in the reproductive and intestinal tissues. The Drosophila model may be key to further elucidating the diverse roles of Notch in tumor formation and growth.
2.4.3 Testis stem cellsThe Drosophila testis contains two stem-cell populations: germline stem cells (GSCs) and cyst stem cells (CySCs) (Cheng et al., 2008; Voog et al., 2008). Maintenance of the stem-cell state requires JAK/STAT signaling from the hub. upd overexpression in germline stem cells leads to a striking overproliferation of early-stage cells which identified as a mixture of undifferentiated germline and somatic cells by their morphology and DNA staining characteristics. The overproliferated germ cells lack Bam consistent with a GSC and/or gonialblast identity (Tulina & Matunis, 2001).
Another model involves restoring the Cdc25 homolog String (Stg) in GSCs in aged testis, causing late-onset tumorigenesis. Stg is highly expressed in young GSCs and CySCs and is required for stem cell maintenance. Interestingly, the expression of Stg declines with age. Restoring Stg expression by using germline specific nanos-Gal4 in GSCs in aged testis prevents the age-associated decline in GSC proliferation and CySC number, but also causes tumors that were filled with undifferentiated GSCs and CySC-like cells. The germ cell tumors in these flies appear to be a mixture of GSCs and spermatogonia, which are confirmed by the presence of spectrosomes/fusomes (Inaba et al., 2011). Thus, the male and female germline tumors share some commonalities in the presence of spectrosomes, failed GSC differentiation and disrupted Bam expression.
2.5 Hematopoietic cellsHematopoiesis in Drosophila occurs throughout the life cycle. Embryonic hematopoiesis occurs at the head mesoderm anlage, whereas larval hematopoietic cells arise from the lymph gland (Holz et al., 2003). In adults, four blood cell clusters, called hematopoietic hubs, have been suggested to be located at the dorsal side of the adult fly abdomen (Ghosh et al., 2015a). Hemocytes or Drosophila blood cells can be divided into three subtypes: plasmatocytes, crystal cells, and lamellocytes. Plasmatocytes and crystal cells are hemocytes that account for about 90–95% and 2–5% of the total population, respectively. Lamellocytes, in contrast, are not normally present in healthy larvae but are instead induced by parasitic infection, tissue damage or hematopoietic tumorigenesis (Ghosh et al., 2015b; Lebestky et al., 2000). Drosophila hematopoiesis is used as a model to elucidate the basic mechanisms of hematopoietic differentiation and homeostasis, and for the study of blood cancers such as myeloproliferative neoplasms (MPNs) and acute myeloid leukemia (AML) (Outa et al., 2020).
MPNs are characterized by clonal proliferation of one or more hematopoietic cell lineages (Tefferi & Vardiman, 2008). The Drosophila hematopoietic tumors also involve overproliferation of either the lamellocytes or crystal cells and formation of melanotic masses (Minakhina & Steward, 2006). The JAK/STAT pathway, because of its prominent role in cell proliferation and differentiation in the hematopoietic lineage, is the major pathway that has been manipulated in Drosophila hematopoietic tumor models. Genetic manipulations of other pathways such as p38 MAPK, Ras, Toll, and JNK could also lead to increases of lamellocytes and formation of melanotic tumors (Zettervall et al., 2004). In addition, transgenic flies misexpressing a fused human gene resulted from the NUP98-HOXA9 translocation, a genetic lesion that leads to AML, helped to reveal a conserved molecular mechanism underlying blood cell expansion (Baril et al., 2017).
Gain-of-function mutations of JAK–STAT pathway components in the larval lymph gland can lead to lamellocyte overproliferation and alteration of the proportion of different hemocyte populations. The core pathway factors in JAK/STAT signaling include Hopscotch (Hop), the JAK2 homolog, and Stat92E, the only Drosophila STAT. Misexpression of a hyperactive STAT92E (Stat92EΔNΔC) by a pan-tissue driver actin-Gal4 leads to melanotic tumor formation in the t
留言 (0)