
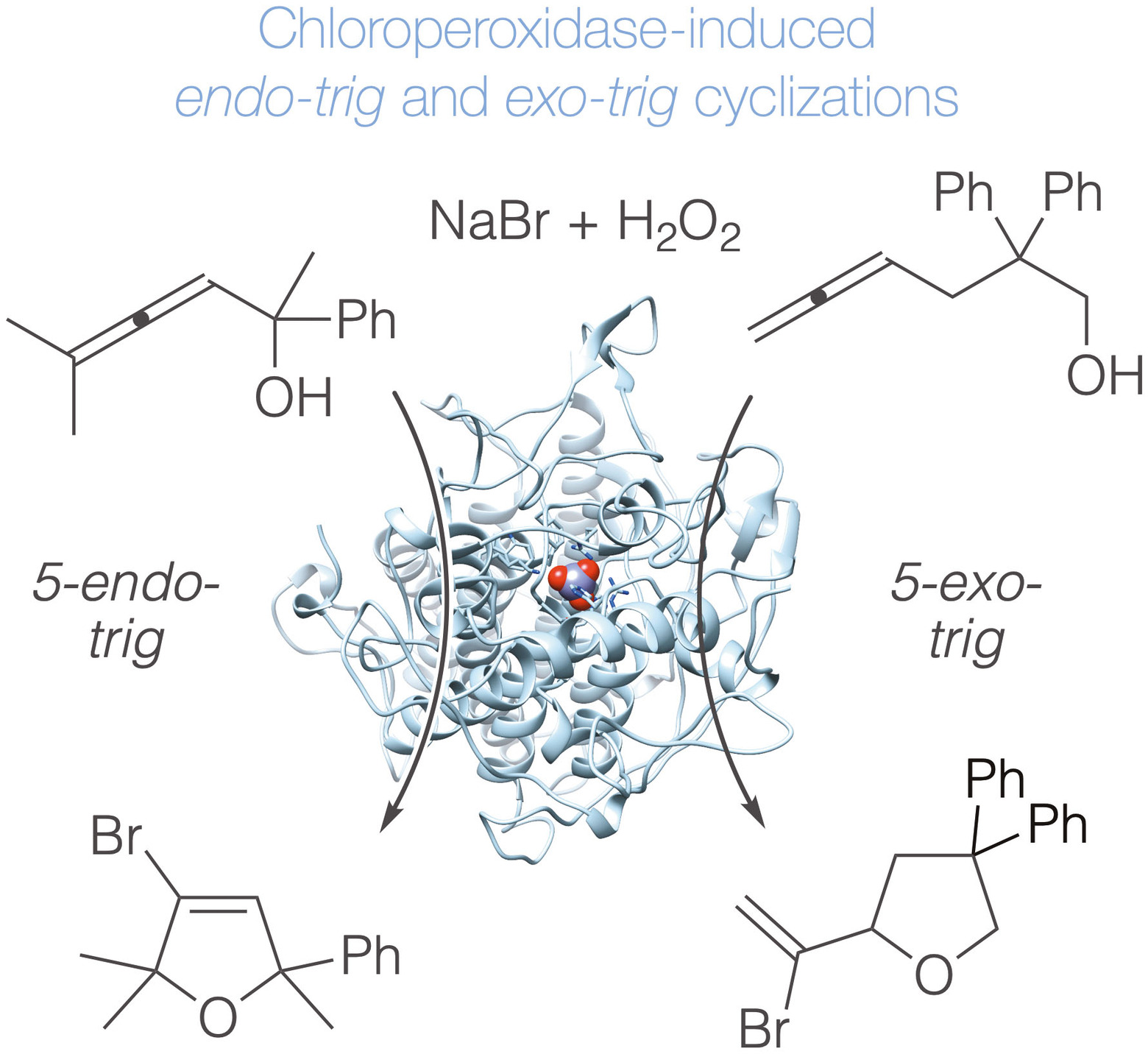
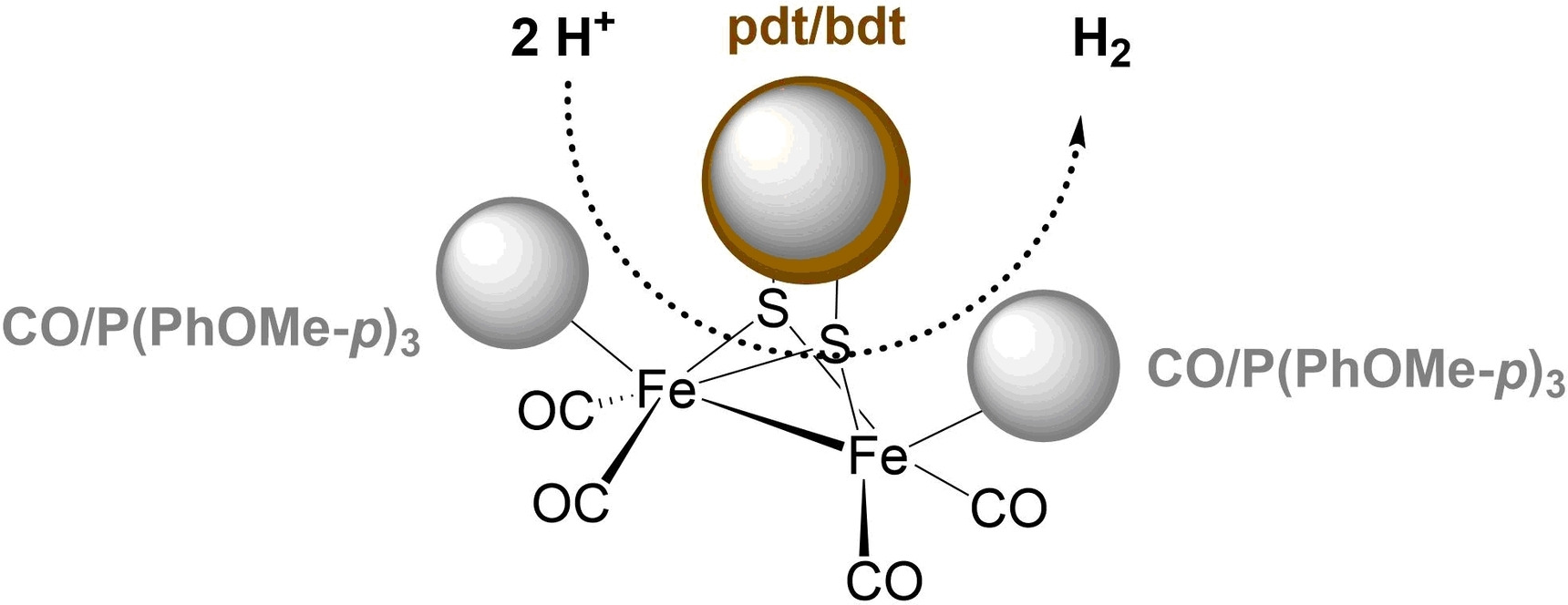
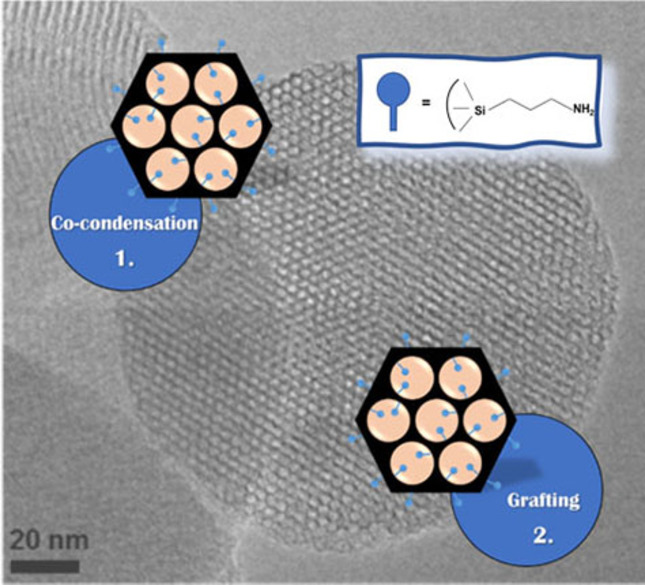
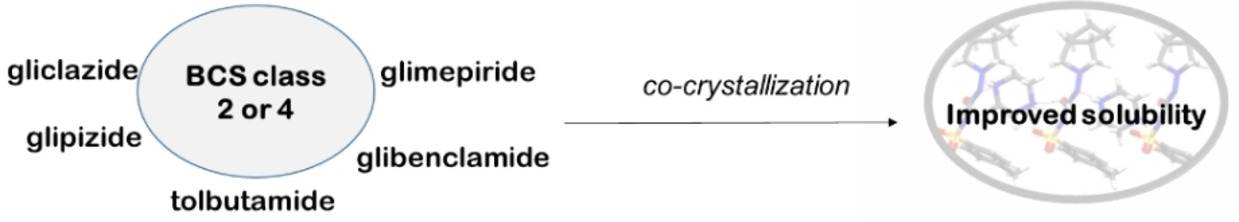
記住我


For each of the two studied substrates, the geometries of a number of enzyme-substrate (ES) complexes were optimized to ensure that the lowest-energy binding modes are identified. The reaction mechanism was then followed for all considered ES complexes. Depending on how the C=N double bond of the imine moiety faces the hydride of the NADPH cofactor, the binding modes can lead to either S- or R-products. In the route to the S-product (S-path), the NADPH is located at the Re face, while in the R-path it is located at the Si face. The binding modes and the following transition states (TSs) will therefore have an -S or an -R in their names in the discussion below.
Reaction of 1-Methyl-3,4-dihydroisoquinolineIn the case of substrate 1, eight ES complexes with different substrate orientations in the active site were optimized. The lowest-energy structures for the complexes leading to S- and R-products (called E:1-S and E:1-R, respectively) are shown in Figure 3, while the other structures with higher energies are depicted in the Supporting Information. In E:1-S, the imino group of 1 forms a hydrogen bond with the backbone of Met122, while in E:1-R, it interacts with one of the water molecules in the active site (w1). In both cases, the overall conformations of the active site residues quite closely resemble the crystal structure, as can be seen from the superposition of the structures provided in the Supporting Information. Interestingly, despite the different orientations of the substrate, the two binding modes E:1-S and E:1-R have the same calculated energy. Other binding modes are up to 10 kcal mol−1 higher in energy, as shown in the Supporting Information.

Optimized structures of A) the lowest-energy enzyme-substrate complexes E:1-S and E:1-R, and B) lowest-energy transition states leading to the two enantiomers, TS-1-S and TS-1-R. Selected distances are given in Å. Relative energies are indicated in kcal mol−1. For clarity, most of the hydrogens are omitted. Atoms kept fixed during the optimizations are labeled with an asterisk.
The transition states of the hydride transfer step were then optimized for all ES complexes. The lowest-energy TS, called TS-1-S, was found to lead to the S-product and stems from the lowest-energy binding mode E:1-S. The calculated barrier for this is 20.1 kcal mol−1 (Figure 4), which is in good agreement with the measured kcat of 0.74 s−1 that corresponds to a barrier of approximately 18 kcal mol−1.10 Importantly, the lowest-energy TS leading to the R-product (TS-1-R) has a calculated barrier of 23.3 kcal mol−1, that is, 3.2 kcal mol−1 higher than the S-path. This result is consistent with the stereopreference observed for the fresh lysate and fresh purified enzyme (85 % and 81 % ee, respectively, in favor of the S-enantiomer).10

Calculated energy profiles for the reaction of substrate 1.
Other optimized TSs are 4–7 kcal mol−1 higher than TS-1-S (see Supporting Information). The resulting enzyme-product complexes, E:2-S and E:2-R, are calculated to be +11.9 kcal mol−1 and +7.9 kcal mol−1 relative to E:1-S, respectively (structures are provided in the Supporting Information).
Scrutinizing the optimized geometries of TS-1-S and TS-1-R in Figure 3, as well as the other TSs in the Supporting Information, helps to shed light on the sources of the observed stereoinduction. We can identify a number of steric repulsions between the substrate and the surrounding groups in TS-1-R that are not present in TS-1-S. The most relevant ones are the repulsions between the aromatic ring of the substrate and the side chains of Ile121 and Val214, and between the methylene group at the C4 position of the substrate and the amide group of the cofactor (Figure 3). In TS-1-S, the substrate points in a different direction and these steric repulsions do not exist.
There are alternative TSs that lead to the R-product that lack these steric clashes, but they suffer from other features that cause their energies to be higher. For example, TS1-1-R2 and TS1-1-R3 (see Supporting Information) lack the H-bond between the imino moiety and the surrounding groups, and in TS1-1-R4 the methyl substituent at the C1 position of the substrate clashes with Pro124.
In addition to the steric clashes, we also note that in TS-1-S the amide group of the cofactor has a favorable non-covalent attractive interaction with the aromatic ring of the substrate, while in TS-1-R, this interaction is lacking. This is visible from a non-covalent interaction (NCI) plot18 provided in the Supporting Information.
Taken together, these contributions result in a higher energy of TS-1-R relative to TS-1-S, inducing thus the observed selectivity. It is interesting to note that the trend in energy between the S- and R-pathways in transition state is not the same as in the ES and EP complexes (Figure 4). In the ES complex, there is practically no difference in energy between the binding modes leading to the different products, while in the EP complex, the R-product binds better than the S-product. These observations mean that it is not possible to predict the stereochemical outcome by simply comparing the energies of the ES or EP complexes as commonly done in docking studies. The full energy profile, including the transition state energies, is required to make a qualified prediction.
To further analyze the stereoselectivity of AoIRED toward substrate 1, we have considered the effects of the Tyr179Ala and Asn241Ala mutations on the calculated energy barriers. Experimentally, an inversion from S to R was observed for both these mutants as compared to the freshly purified wild-type enzyme, with >98 % ee for Tyr179Ala and 71 % ee for Asn241Ala.10
We introduced these mutations to the active site model by replacing the side chains of the tyrosine or the asparagine residues by an ethyl group, representing the alpha and beta carbons of alanine. Such a protocol has been successfully applied in previous work addressing the effects of mutations on the enantioselectivity.13a, 13i For both mutants, we optimized a number of transition states for the hydride transfer, considering different binding modes and orientations of the substrate (in total 11 TSs for each mutation). The calculations could reproduce the experimental results, in that the energy of the TS for the R-pathway was found to be lower than the S-pathway for both the Tyr179Ala and Asn241Ala mutations, by 2.1 and 2.0 kcal mol−1, respectively. By comparing the optimized TS structures of the R- and S-pathways with each other and with their wild-type counterparts, we can analyze the reasons for the observed reversal in selectivity.
We first note that both mutations disrupt the hydrogen bonding network at the active site between Tyr179, Asn241, and the backbone of Ala236. This generates a slightly larger cavity, which allows the substrate to move somewhat more than in the wild-type (Figure 5).

Optimized structures of the lowest-energy transitions states of A) Tyr179Ala (TS-1-STyr179Ala and TS-1-RTyr179Ala) and B) Asn241Ala (TS-1-SAsn241Ala and TS-1-RAsn241Ala) mutations. Selected distances are given in Å. Relative energies for each pair of TSs are indicated in kcal mol−1. For clarity, most of the hydrogens are omitted. Atoms kept fixed during the optimizations are labeled with an asterisk.
In the case of TS-1-RTyr179Ala, a hydrogen bond is formed between the imino group of the substrate and the backbone carbonyl of Ala236, an interaction that is neither present in the TS-1-STyr179Ala nor in the wild-type case. This is mainly caused by a slight rotation of the Asn241 and Met240 side chains, a movement that creates a small cavity that can accommodate the methyl substituent of the substrate.
This rationalization of the effect of Tyr179 on the enantioselectivity is different from the proposal in the literature, where this residue was suggested to form a hydrogen bond to the substrate and thereby orienting it in a specific way.10 Such an interaction is observed in the crystal structure with the R-2 product bound (see Figure 1). However, no hydrogen bond can be observed between the tyrosine and the substrate in the calculations, neither in the wild-type nor in the mutant.
In the case of TS-1-RAsn241Ala, the orientation of the substrate in the active site is the same as in the wild-type case (TS-1-R). Tyr179 is now slightly rotated to engage in a hydrogen bond with Thr244 (Figure 5). Importantly, the mutation of Asn241 and the rotation of Tyr179 create slightly more room for the substrate to move into that direction, alleviating the steric clashes with Val214 and Ile121. This therefore results in a lower energy for TS-1-RAsn241Ala compared to TS-1-SAsn241Ala.
The above analysis shows that it is not straightforward to predict the effects of a specific mutations on the enantioselectivity without calculations, because they can both cause the substrate to bind differently and also affect the interactions and energies of the transition states.
Finally, it is important to point out that we have, in the design of the active site model for the current calculations, used the crystal structure in complex with R-2 (PDB 5FWN). The results for the wild-type active site are consistent with this enzyme being (S)-selective, as observed for the freshly purified enzyme. The fact that the calculations also reproduce the switch in enantioselectivity for the Tyr179Ala and Asn241Ala mutants lends further support to this notion. Nevertheless, it cannot be excluded that the enzyme somehow transforms to the (R)-selective form during the crystallization process.10
Reaction of 2-Propyl-piperideineTo further investigate the enantioselectivity of AoIRED, we have considered the reaction with 2-propyl-piperideine, 3, which shows the opposite selectivity compared to 1, that is, >99 % ee in favor of the R-product.10 This substrate is smaller than 1 and has also an alkyl substituent with rotatable bonds, which will affect its binding to the active site and also the geometry of its transition states.
We have optimized 12 ES complexes with different substrate orientations and conformations. The lowest-energy pro-R and pro-S structures are shown in Figure 6, while the other structures are given in the Supporting Information.

A) Optimized structures of the lowest-energy enzyme-substrate complexes E:3-S and E:3-R, and B) lowest-energy transition states leading to the two enantiomers, TS-3-S and TS-3-R2. Selected distances are given in Å. Relative energies are indicated in kcal mol−1. For clarity, most of the hydrogens are omitted. Atoms kept fixed during the optimizations are labeled with an asterisk.
In contrast to substrate 1, the lowest-energy E:3-R and E:3-S binding modes differ by as much as 4.4 kcal mol−1, in favor of the former (Figures 6 and 7).

Calculated energy profiles for the reaction of substrate 3.
In the E:3-S, the imino group of the substrate forms hydrogen bond with carbonyl group of Met122, while in E:3-R the hydrogen bond is with the amide group of the cofactor. In both cases, the n-propyl substituent is located in the solvent-exposed side of catalytic pocket (Figure 6). The reason for the higher energy of E:3-S is that the n-propyl moiety clashes with Leu175, Tyr179 and Met240. Other pro-S binding modes that lack these clashes are higher in energy because of missing hydrogen bond of the imino group.
For each of the binding modes we have calculated the transition state for the hydride transfer. Interestingly, the lowest-energy TS leading to the R-product does not stem from the lowest-energy binding mode, but from an ES complex that is slightly higher in energy (E:3-R2, +0.9 kcal/mol, see Supporting Information), and will therefore be called TS-3-R2. The calculated barriers for TS-3-R2 and TS-3-S are 20.6 kcal/mol and 21.1 kcal/mol, respectively. The lowest-energy enzyme-product complexes are +5.8 and +7.8 kcal/mol relative to E:3-R, for the S- and R-pathways, respectively (see Supporting Information).
The computations are thus in agreement with the experimental outcome, favoring the formation of R-product,10 but the calculated energy difference of 0.5 kcal/mol is underestimated. As discussed in previous work on modeling enzymatic enantioselectivity, the reason for the underestimation is most likely related to the slight inflexibility of the active site model, which can miss to perfectly adapting to the size of the substrate.13, 14 However, it is important to underline that, since the modeling of enantioselectivity deals with very small energy differences, a perfect numerical agreement with experiments is very difficult to achieve, and the reproduction of experimentally observed trends is generally considered as a very satisfactory result.
Comparisons of the optimized geometries of transitions states provide information about the origins of the observed enantioselectivity. Interestingly, the imino group is not involved in any hydrogen bonds in the lowest-energy transition states TS-3-R2 and TS-3-S. In TS-3-S, the n-propyl group of the substrate clashes with the side chains of Tyr179 and Leu175. In TS-3-R2 the n-propyl points in the opposite direction, making a better fit with the active site cavity.
An interesting aspect about substrate 3 is that there are other TSs for both the R- and the S-pathways that are not much higher in energy compared to TS-3-R2 and TS-3-S (see Supporting Information). For example, there are three TSs in the S-pathway that are within 1 kcal mol−1 from TS-3-S. This result shows that one has to consider all possible conformations and orientations of the substrate in the active site in order to reproduce and rationalize the enantioselectivity. It also shows that it is very difficult to a priori predict how a specific mutation will affect the relative energies, or what the consequences of even a small modification of the substrate will be on the enantioselectivity.
留言 (0)