
記住我


Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disease that primarily affects both cortical and spinal motor neurons [1]. Several genes have been implicated in ALS pathogenesis [2]; however, mutations in these genes account for a minority of cases, and the etiology of the disease remains to be elucidated. Therefore, understanding the exact molecular mechanisms leading to motor neuron loss is crucial for the development of new therapeutic approaches and the discovery of novel and useful biomarkers of disease.
Recent studies have begun to link alteration in tau phosphorylation to ALS pathogenesis with tau pathology reported in both sporadic and familial cases [3, 4]. Tau protein is a member of the microtubule-associated protein (MAP) family and plays a critical role in stabilizing microtubules, the major component of the eukaryotic cytoskeleton involved in cell processes such as cell division, mobility, and intracellular organization, and trafficking of organelles [5]. Tau hyperphosphorylation, accumulation, and mutations have been linked to a group of progressive neurodegenerative diseases collectively known as tauopathies [6-8], in which hyperphosphorylation of key epitopes on tau promotes its disassembly from microtubules, aggregation, and subcellular mislocalization, leading to the formation of inclusions and neurofibrillary tangles (NFTs) in both neurons and glia [6, 8].
Although the exact molecular mechanisms underlying tau toxicity are not yet fully understood, the main consequence of the accumulation of toxic tau is the disruption of neuronal transport [9-11]. This impairment is an early pathogenic event in neurodegeneration, and tau-mediated alterations in neuronal trafficking have been described in several neurodegenerative diseases [9-11]. Furthermore, deficits in neuronal transport were shown to disrupt several cellular functions, including but not limited to alterations in both trafficking and function of mitochondria, synapse loss, excitotoxicity, and cell death [9, 11, 12].
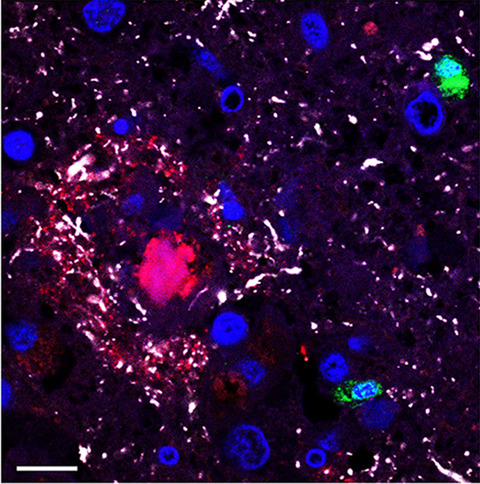
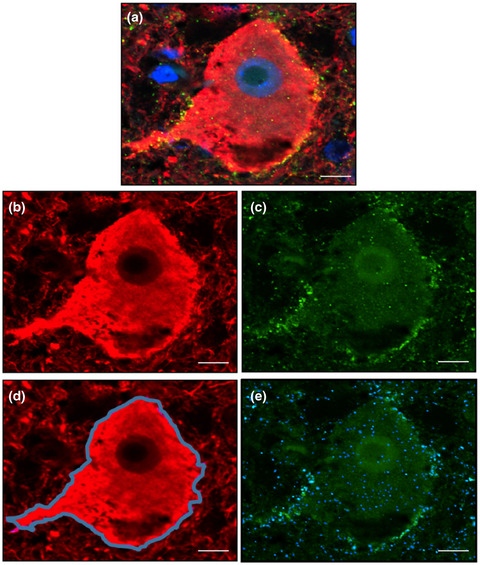
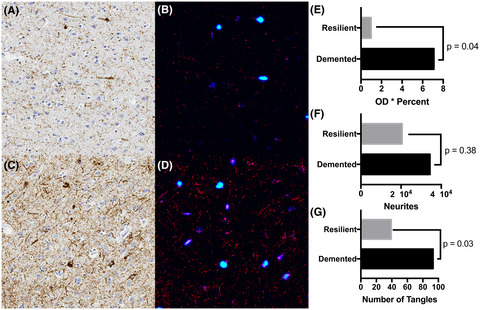
In ALS, there is a significant increase in total tau as well as cytoplasmic inclusions of hyperphosphorylated tau (T175, T217, S208/210, S212, S396, and S404) in post-mortem motor cortex (mCTX) and spinal cord of ALS patients [13-15]. Moreover, alterations in tau and pTau:tau ratio have been reported in ALS cerebrospinal fluid (CSF) [16-19]. Importantly, tau-induced alterations in cellular processes such as excitotoxicity, mitochondrial dysfunction, synapse loss, and impairments of nucleocytoplasmic transport, are also pathogenic features of ALS [1, 20, 21], suggesting that alterations in tau could underlie these molecular events in ALS. Accordingly, recent findings from our group have revealed that hyperphosphorylated tau at S396 (pTau-S396) mislocalizes to synapses in mCTX across ALS subtypes and contributes to mitochondrial fragmentation through interaction with the pro-fission GTPase dynamin-related protein 1 (DRP1) [22], further supporting a pathogenic role for tau in ALS.
Here, we used a large cohort of ALS post-mortem mCTX samples to further investigate whether there were alterations in tau phosphorylation in ALS. In addition, we sought to determine whether there were novel genetic variants in MAPT, the gene encoding tau, in ALS. Lastly, we measured tau and pTau in CSF derived from people living with ALS and healthy controls given the contradictory results from previous biomarker studies in ALS.
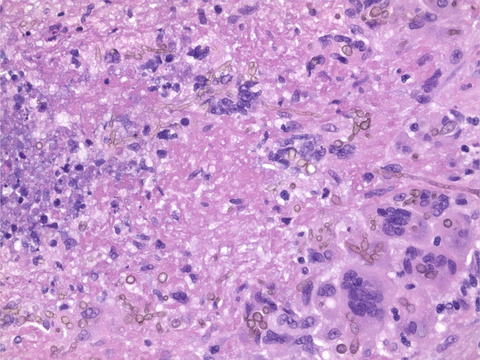
2 MATERIALS AND METHODS 2.1 Human tissue samplesPost-mortem mCTX from ALS and controls were provided by the Massachusetts Alzheimer's Disease Research Center (ADRC) and from the Veterans Affairs Biorepository Brain Bank (VABBB; Merit review BZ002466) with approval from the Massachusetts General Hospital Institutional Review Board (IRB). In total, we assessed 52 ALS and 25 non-neurological control mCTX as well as entorhinal cortex (EC) from two Alzheimer's disease (AD) cases as positive controls. Sixteen ALS and seven control mCTX were used for immunohistochemistry (IHC): the mean age was 68.9 years (SD = 14.6) for control and 62.4 years (SD = 11.5) for ALS. Control samples were 57.1% male, while the ALS group was 68.8% male. Twelve ALS cases were diagnosed with limb onset disease, while two were diagnosed with bulbar onset. Region of ALS onset was unknown for two samples. One case was diagnosed with ALS/frontotemporal dementia (ALS/FTD); however, post-mortem evaluation revealed no positive staining for either β-amyloid or α-synuclein as well as no NFTs or Lewy bodies. Two of the ALS cases were positive for C9ORF72 repeat expansion. Genetic status of all other ALS cases was unknown. All 16 ALS mCTX demonstrated TDP43 proteinopathy. Post-mortem interval (PMI) range was 14–81 h for controls and 20–77 h for ALS. Both control and ALS groups were negative for β-amyloid and α-synuclein upon post-mortem evaluation by MGH ADRC and VABBB, except for a single ALS case that revealed brain alterations likely due to AD (CERAD plaque stage: sparse; Braak neurofibrillary stage: III/IV; moderate amyloid angiopathy) but insufficient for a concomitant diagnosis of AD. Clinicopathological information for the control and all other ALS post-mortem mCTX samples used in this study are summarized in Table 1. The AD cases were both male, aged 88 and 61 years old (PMI: unknown, and 24 h, respectively).
TABLE 1. Post-mortem motor cortex sample information Sex Onset site Genotype fALS Protein aggregates FTD Age at disease onset Age of death PMI Control 1 M N/A N/A N/A N/A N/A N/A 82 50 Control 2 M N/A N/A N/A N/A N/A N/A 66 81 Control 3 M N/A N/A N/A N/A N/A N/A 49 45 Control 4 F N/A N/A N/A N/A N/A N/A 58 18 Control 5 M N/A N/A N/A N/A N/A N/A 60 14 Control 6 F N/A N/A N/A N/A N/A N/A 77 72 Control 7 F N/A N/A N/A N/A N/A N/A 90 24 Control 8 F N/A N/A N/A N/A N/A N/A 79 Unknown Control 9 F N/A N/A N/A N/A N/A N/A 57 13 Control 10 F N/A N/A N/A N/A N/A N/A 60 15 Control 11 M N/A N/A N/A N/A N/A N/A 63 16 Control 12 M N/A N/A N/A N/A N/A N/A 86 10 Control 13 F N/A N/A N/A N/A N/A N/A 73 20 Control 14 M N/A N/A N/A N/A N/A N/A 92 23 Control 15 F N/A N/A N/A N/A N/A N/A 52 10 Control 16 M N/A N/A N/A N/A N/A N/A 55 19 Control 17 F N/A N/A N/A N/A N/A N/A 85 24 Control 18 F N/A N/A N/A N/A N/A N/A >90 45 Control 19 F N/A N/A N/A N/A N/A N/A 79 9 Control 20 M N/A N/A N/A N/A N/A N/A >90 23 Control 21 F N/A N/A N/A N/A N/A N/A >90 24 Control 2 M N/A N/A N/A N/A N/A N/A 89 36 Control 23 M N/A N/A N/A N/A N/A N/A 70 56 Control 24 M N/A N/A N/A N/A N/A N/A >90 86 Control 25 M N/A N/A N/A N/A N/A N/A 66 71 ALS 1 M Limb Unknown Unknown TDP43 No 57 62 46 ALS 2 F Limb Unknown Unknown TDP43; Ubiquitin No 22 34 65 ALS 3 M Bulbar C9ORF72 Yes TDP43; Ubiquitin No 55 61 77 ALS 5 M Limb Unknown No TDP43 No 58 66 43 ALS 6 M Limb Unknown No TDP43 No 62 68 43 ALS 7 M Limb Unknown No TDP43 No 54 78 35 ALS 8 M Limb Unknown Unknown TDP43; tau No 74 85 38 ALS 9 F Limb Unknown Unknown TDP43 No 64 70 30 ALS 10 M Unknown Unknown Unknown TDP43 No Unknown 58 30 ALS 11 M Limb Unknown Unknown TDP43 No Unknown 62 24 ALS 12 M Limb C9ORF72 Yes TDP43 No 57 61 22 ALS 13 M Unknown Unknown Unknown TDP43 No Unknown 64 20 ALS 14 M Limb Unknown Unknown TDP43 No 41 62 24 ALS 15 F Limb Unknown Unknown Ubiquitin No 65 66 24 ALS 16 F Bulbar Unknown Unknown TDP43 No 46 49 25 ALS 17 F Limb Unknown Unknown TDP43 Yes 52 53 30 ALS 18 M Limb Unknown No Unknown Unknown 75 79 Unknown ALS 19 M Unknown Unknown Unknown Unknown Unknown Unknown 69 24 ALS 20 M Bulbar Unknown Unknown Unknown Yes 58 61 24 ALS 21 M Bulbar Unknown Unknown Unknown No 79 81 4 ALS 22 M Limb Unknown Unknown Unknown Unknown 75 76 14 ALS 23 F Bulbar Unknown No Unknown No 76 Unknown 24 ALS 24 M Unknown Unknown Unknown TDP43 Unknown Unknown 58 30 ALS 25 F Unknown C9ORF72 Unknown TDP43 Unknown Unknown 67 19 ALS 26 M Limb Unknown No Unknown No 53 53 24 ALS 27 M Limb Unknown Unknown Unknown No 59 60 19 ALS 28 M Limb Unknown Unknown Unknown No 57 60 22 ALS 29 M Limb Unknown No TDP43 No 49 51 24 ALS 30 M Limb Unknown No TDP43; Ubiquitin No 58 62 24 ALS 31 F Limb Unknown No TDP43 No 90 91 33 ALS 32 F Bulbar Unknown No Unknown No 50 52 28 ALS 33 F Bulbar C9ORF72 Unknown TDP43 No 54 56 Unknown ALS 34 F Bulbar C9ORF72 Unknown TDP43 No 49 58 48 ALS 35 M Bulbar Unknown No Unknown No 53 55 36 ALS 36 M Limb Unknown No Unknown No 50
留言 (0)