
記住我

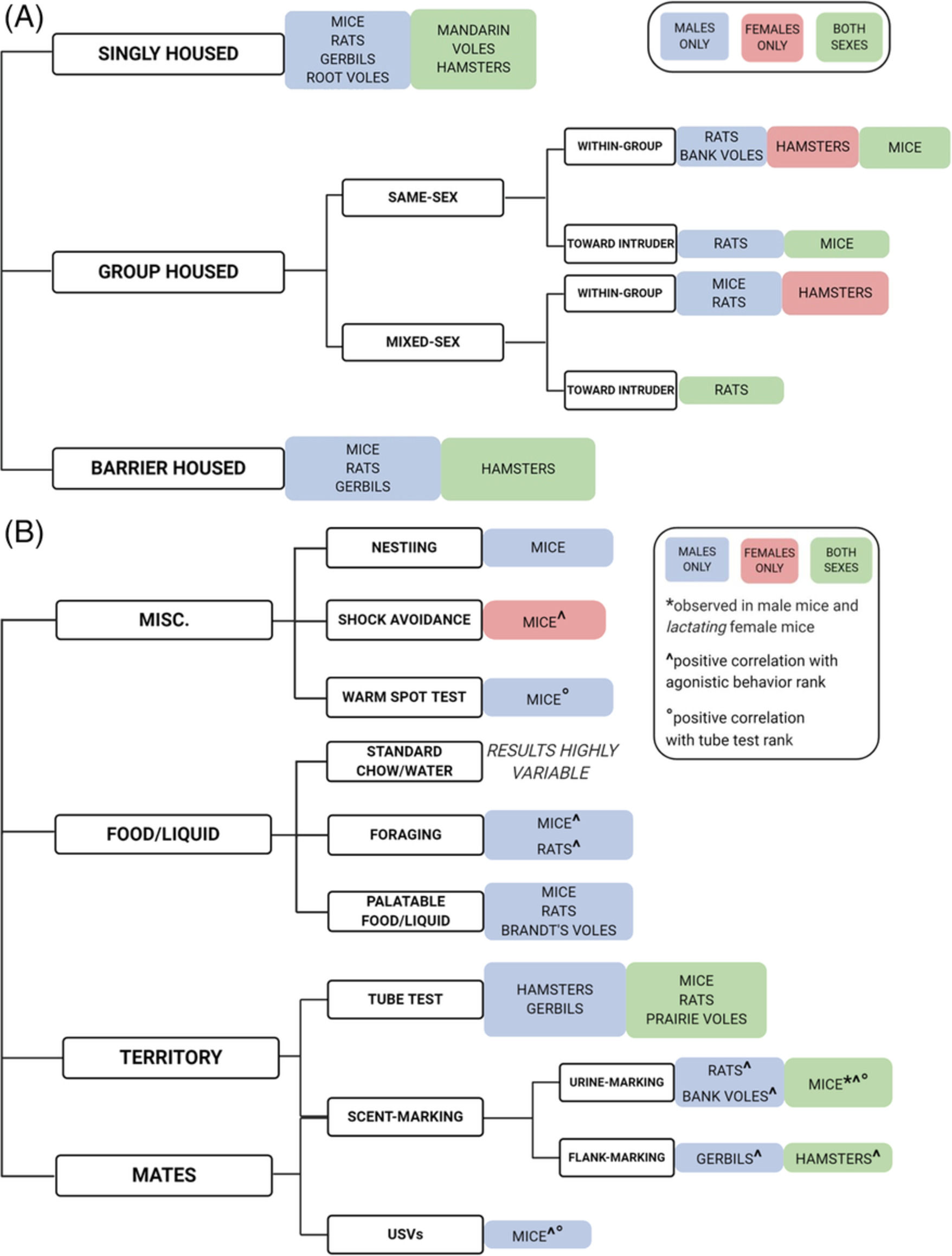



Not all individuals who initiate drug or alcohol use will eventually lose control over their use and/or meet diagnostic criteria for substance use disorder (SUD). This progression is moderated by a myriad of genetic and environmental factors that interact with one another in a developmentally- and sex-dependent manner.1-4 Evidence from twin studies supports the idea that the majority of risk for developing an SUD relates to a single substance-nonspecific genetic factor, with lesser influences of common and unique environmental influences,5 but more recent genome-wide association studies indicate a multitude of common and distinct genetic factors.6, 7 A major focus of current addiction studies is therefore to discover potential genetic and neurobiological factors that moderate individual SUD risk. Multiple heritable phenotypes have been shown to be predictive for heightened likelihood to seek out and use drugs in both humans and laboratory animals, including novelty preference and seeking,8-10 locomotor response to novelty,4, 11 anxiety-related behaviors,12-14 altered stress responses,15-17 circadian phenotypes,18, 19 and impulsivity. These behaviors may have overlapping neurogenetic components, and by studying them in tandem, common and unique genetic factors may be identified and investigated for their relationship to SUD traits.
Impulsivity is the trait-like proclivity to engage in excessive, uncontrolled, or rash reward pursuit and consumption.20-22 These behaviors are considered pathological when they are intrusive, disrupt normal life routines, cause clinical distress, or lead to harmful outcomes.23 Impulsivity is furthermore identified as having multiple dimensions, each of which can be separately measured and may have a unique relationship to addiction vulnerability. This is exemplified in the widely used Barratt Impulsiveness Scale (BIS-11), which measures self-reported cognitive, motor, and non-planning impulsiveness.24 Behavioral tasks have also been developed to quantify impulsive phenotypes in humans, such as the Go/No-Go task, delay discounting, reversal learning, Five-Choice Serial Reaction Time Task (5-CSRTT), and stop signal reaction time task, which have analogs for use in animal models. Research on animal models exposed a unique relationship between types of impulsivity and various pharmacological interventions, in that drug treatment did not uniformly increase, decrease, or maintain different impulsive phenotypes.21, 25, 26 No significant bivariate correlation between impulsive action (5-CSRTT) and impulsive choice (delay discounting) has been found in either lab rats or humans; and furthermore, three factors reflecting statistically orthogonal measures of impulsivity were identified in human subjects: self-report, impulsive action, and impulsive choice.27, 28 These findings may very likely be attributed to different underlying neurogenetic mechanisms regulating each type of impulsivity.
Delay discounting, a common test of impulsive choice, was initially created to assess rats and pigeons,29 although it is now used in both human and animal subjects with variations to the methodology. Delay discounting is a paradigm established to assess an individual's tendency to reduce (discount) the subjective value of a reward if it must wait to receive it. The delay discounting procedure aims to establish how the subject therefore discounts the delayed reward, either by altering the volume of reward or the length of delay. A fundamental aspect of delay discounting is that the subject makes an action to choose one of two options and then must wait for the consequence, thus differing from other similar procedures such as the differential reinforcement task,21 and measuring impulsive choice (frequent selection of the small immediate lever) versus self-control (frequent selection of the large, delayed option).30
Reversal learning, a measure hypothetically linked with impulsive action, revolves around changing reinforcement contingencies: one of an array of actions (e.g. pressing the left most lever) is paired with an outcome (e.g. receiving a food reward), and the subject learns to discriminate between those contingencies.31 After reaching an accuracy criterion, the contingencies are reversed (e.g., only pressing the right lever, not the left, now leads to the food reward). Subjects must show cognitive and behavioral flexibility by constraining their previous responses and discarding the initially learned rule in order to maximize reward. Greater difficulty with stopping or updating behavior during reversal learning has been suggested to reflect greater impulsive action. Studies have shown that this behavioral inflexibility is genetically linked to impulsivity.31, 32 In addition to responding after reversal of response contingency, premature (inter-trial interval) responding within reversal learning can be measured. This measure is analogous to premature responding in the 5-CSRTT. Both measures of anticipatory responses are considered to be waiting impulsivity, or the inability to withhold response in anticipation of a reward-related cue.20
Inbred mouse panels permit estimates of heritability of these traits, which is operationally defined as the proportion of phenotypic variation that is explained by genetic variation. In inbred lines, heritability is estimated as the proportion of phenotypic variance accounted for by strain. Past genetic reference population studies have considered this to be an effective estimate of heritability, considering each mouse from each strain is, to the extent maximally possible, genetically identical to one another.33, 34 Environmental and technical sources of variance are reduced within these panels, further increasing the ability to detect heritability of a trait over external influence and providing an advantage over human twin studies.35
Past efforts have utilized recombinant inbred (RI) panels to identify the heritability and genetic architecture of impulsive phenotypes. Laughin et al36 conducted reversal learning in 51 BXD mouse strains and identified heritable strain variation in impulsivity, as well as a genome-wide significant quantitative trait locus (QTL) on chromosome 10. Positional candidate genes, including Syn3—encoding the synaptic phosphoprotein Synapsin III—expressed from this QTL were selected based upon expression phenotypes that were genetically correlated with the behavioral phenotype. A subsequent study using the BXD strains that exhibited extreme differences in reversal learning found that the poor reversal learning strains more rapidly acquired cocaine self-administration and administer cocaine at greater rates than do good reversal learning strains.37
Utilizing mouse populations with greater genetic diversity may expand observed phenotypic ranges and lead to new insights in neurogenetics and neurobiology of impulsivity and its relationship to SUDs. The collaborative cross is a genetically diverse, multi-parental RI panel. It alleviates genetic bottleneck limitations through the intercross of five classical inbred strains (A/J, C57BL/6J, 129S1/SvImJ, NOD/LtJ, NZO/HlLtJ), and three wild-derived inbred strains (CAST/EiJ, PWK/PhJ, WSB/EiJ), to capture >90% of the known genetic variation present in the laboratory mouse.38 In addition to having a large amount of genetic diversity, the collaborative cross also has generally balanced allele frequencies and evenly distributed recombination sites,39, 40 resulting in greater statistical power to detect genetic correlations among phenotypes gathered in different laboratories.
The present study aims to examine heritability and genetic correlations between locomotor response to novelty, palatable food consumption (a measure of reward sensitivity), reinforcement learning, two tests of impulsivity (delay discounting and reversal learning), and other cataloged addiction-related behaviors measured by others in the 8 CC founder strains and 10 CC strains. The tests of impulsivity are designed to measure three types of impulsivity: impulsive action (total trials to criteria in reversal learning), waiting impulsivity (anticipatory responses measured during reversal learning) and impulsive choice (indices of preference for an immediate reward). We hypothesized that the three measures of impulsivity are heritable traits, although not necessarily coinherited (genetically correlated). Locomotor activity and palatable food consumption are similarly anticipated to be heritable traits, each of which has shown conflicting evidence in the literature of being positively correlated with impulsivity.41-43 These studies were conducted as part of a multi-lab collaboration in the Center for Systems Neurogenetics of Addiction. A set of strains were tested across different laboratories for impulsivity traits as well as SUD-related traits. Results from these efforts will provide insights into the inter-lab replicability of impulsivity traits as well as the genetic relationships among a large number of traits that may be predictive of SUDs.
2 MATERIAL AND METHODS 2.1 SubjectsAll inbred and recombinant inbred mice involved in this study were born at The Jackson Laboratory (JAX; Bar Harbor ME). A subset was subsequently shipped to Binghamton University, while the remainder were phenotyped at JAX, in the facilities of the Behavioral Phenotyping Core of the Center for Systems Neurogenetics of Addiction. All procedures were performed according to the “Guide for the Care and Use of Laboratory Animals” (National Research Council, 2011) in the AAALAC accredited programs at Binghamton University or JAX, after approval by the relevant Institutional Animal Care and Use Committees.
2.1.1 Binghamton University siteSubjects were shipped from JAX to Binghamton University between 35–49 days of age. A total of 6 cohorts were shipped between 8/2016 and 8/2019. Mice from the eight founder strains were delivered in the first three cohorts, each of which was balanced to include representatives from all 8 strains and both sexes. Ten strains of CC mice were also studied; they were first included in cohort three. Details on mouse strains/cohorts are provided in Table 1. A total of 286 mice were tested at the Binghamton site.
TABLE 1. Strains tested at the Binghamton University site Mouse ID JAX Stock No. MGI ID N Cohorts NOD/ShiLtJ 001976 2162056 18 1–3 129S1/SvImJ 002448 2160041 18 1–3 A/J 000646 2159747 18 1–3 C57BL/6J 000664 3028467 18 1–3 NZO/HlLtJ 002105 2668669 18 1–3 CAST/EiJ 000928 2159793 18 1–3 WSB/EiJ 001145 2160667 18 1–3 PWK/PhJ 003715 2160654 18 1–3 CC002/UncJ 021236 5649080 12 4–6 CC004/TauUncJ 020944 5649082 23 3–6 CC006/TauUncJ 022869 5649237 12 4–6 CC011/UncJ 018854 5649240 12 4–6 CC012/GeniUncJ 028409 5694080 12 4–6 CC025/GeniUncJ 018857 5649246 11 4–6 CC028/GeniUncJ 025126 5659485 12 4–6 CC032/GeniUncJ 020946 5649248 12 4–6 CC041/TauUncJ 021893 5649251 24 3–6 CC061/GeniUncJ 023826 5649258 12 4–6Upon arrival, mice were housed in the same groups in which they were shipped, with three mice of the same strain and sex being grouped together in a cage. The colony room was maintained on a 12 h light/dark cycle (lights on at 0615 h) and at an average temperature of 69–70°F. During this initial acclimation period, food (Lab Diet 5001, ScottPharma Solutions) and water was available ad libitum. A nestlet and a translucent red acrylic tube (9.75 cm long, 5 cm diameter, approximately 65 g) were placed in each cage. For removal from the cage, the mice were briefly handled by their tails using a gloved hand. Wild-derived inbred strains and some CC strains (CC004/TauUncJ and CC011/UncJ) were handled by the tail with forceps.
All three wild-derived strains (CAST/EiJ, WSB/EiJ, and PWK/PhJ) exhibit high levels of aggression and are at high risk of injury when group-housed in adulthood. To avoid injury, all mice of both sexes and of all strains, including the wild and non-wild derived lines, were singly housed in identical caging conditions starting at PND 60. Thus, all animals tested in this study experienced a uniform environment, and single-housing was only instituted in adulthood. Mice were acclimated to these conditions for 10 days until PND 70.
Prior to the initiation of the operant conditioning protocols described below, mice were introduced to a schedule of limited access to chow. Mice were weighed daily during food restriction and percent of free-feeding body weight was calculated by dividing the current weight by the free-feeding weight. During the limited access to food period, mice were fed once a day between 3 pm and 5 pm; chow quantity provided per day was titrated until mice reach 80%–85% or 83%–88% (CAST/EiJ, PWK/PhJ, WSB/EiJ) of their free feeding weights. Wild-derived inbred strains were maintained at a higher free-feeding percentage because of observed lethargy and dehydration when their weights approached 80%. Once mice reached their target weights, operant testing began (see Table 2 for reversal learning mouse weights; Table 3 for delay discounting mouse weights). If, at any point during the testing period, a mouse dropped below 80% of its free feeding weight, the quantity of chow provided was increased. If increased food availability did not lead to a recovery of body weight to ≥80% within a day, it was temporarily returned to ad libitum food access until its weight had recovered.
TABLE 2. Average of strain's body weight during each stage of reversal learning testing, represented as a percentage of their initial free-feeding weight Mouse ID Start Training Acquisition Reversal NOD/ShiLtJ 27.2 ± 0.99 g 85.8 ± 0.76% 84.2 ± 0.71% 85.8 ± 0.74% 129S1/SvImJ 23.7 ± 0.64 g 88.0 ± 1.04% 86.6 ± 1.09% 85.6 ± 1.02% A/J 21.8 ± 0.61 g 87.8 ± 0.62% 84.7 ± 0.65% 84.8 ± 0.52% C57BL/6J 23.6 ± 0.51 g 85.6 ± 0.47% 84.7 ± 0.61% 85.1 ± 0.54% NZO/HlLtJ 41.0 ± 1.24 g 88.4 ± 1.30% 83.7 ± 0.92% 83.6 ± 0.64% CAST/EiJ 15.5 ± 0.38 g 86.0 ± 0.92% 85.3 ± 0.86% 84.4 ± 0.65% WSB/EiJ 15.7 ± 0.40 g 86.7 ± 0.92% 86.3 ± 0.77% 87.6 ± 0.74% PWK/PhJ 17.2 ± 0.40 g 85.5 ± 0.43% 85.7 ± 0.67% 85.1 ± 0.61% CC001/UncJ 24.3 ± 0.90 g 84.1 ± 0.94% 83.1 ± 0.88% 82.8 ± 0.92% CC002/UncJ 24.2 ± 2.07 g 86.7 ± 0.66% 85.5 ± 0.56% 84.8 ± 0.86% CC003//UncJ 27.7 ± 0.59 g 84.0 ± 1.04% 84.8 ± 0.98% 84.9 ± 1.15% CC004/TauUncJ 28.6 ± 1.44 g 86.8 ± 1.76% 84.7 ± 0.94% 85.6 ± 1.45% CC005/TauUncJ 19.9 ± 0.92 g 87.1 ± 0.89% 85.8 ± 0.86% 85.9 ± 0.57% CC006/TauUncJ 23.8 ± 0.98 g 86.0 ± 1.04% 87.0 ± 0.71% 86.9 ± 0.57% CC008/GeniUncJ 32.4 ± 0.73 g 84.7 ± 1.01% 86.5 ± 0.86% 85.7 ± 0.85% CC011/UncJ 25.7 ± 1.01 g 87.6 ± 0.88% 85.5 ± 1.09% 85.4 ± 0.73% CC012/GeniUncJ 20.8 ± 1.38 g 87.3 ± 1.21% 86.6 ± 1.17% 87.9 ± 0.49% CC013/GeniUncJ 27.2 ± 1.01 g 83.8 ± 0.88% 86.0 ± 0.98% 86.8 ± 0.77% CC015/UncJ 21.0 ± 1.36 g 86.1 ± 0.78% 83.5 ± 0.77% 84.8 ± 0.90% CC016/GeniUncJ 24.6 ± 1.87 g 85.0 ± 0.33% 83.2 ± 0.49% 82.7 ± 0.36% CC017/UncJ 21.0 ± .59 g 85.3 ± 0.67% 83.9 ± 0.93% 84.4 ± 0.90% CC019/TauUncJ 19.1 ± 2.91 g 86.3 ± 0.27% 84.7 ± 2.25% 83.8 ± 2.17% CC023/GeniUncJ 22.5 ± 1.23 g 84.4 ± 85.1% 85.1 ± 2.48% 82.2 ± 1.59% CC025/GeniUncJ 23.6 ± 1.43 g 86.1 ± 0.32% 83.1 ± 1.83% 83.1 ± 0.50% CC026/GeniUncJ 25.3 ± 1.79 g 84.8 ± 0.50% 84.5 ± 0.83% 82.9 ± 0.65% CC027/GeniUncJ 24.4 ± 1.40 g 86.9 ± 0.50% 85.1 ± 1.11% 83.7 ± 1.13% CC028/GeniUncJ 29.4 ± 2.03 g 86.3 ± 0.57% 84.5 ± 0.27% 83.8 ± 1.18% CC030/GeniUncJ 23.9 ± 1.23 g 86.4 ± 0.50% 85.2 ± 0.20% 85.7 ± 0.44% CC032/GeniUncJ 25.7 ± 1.60 g 90.7 ± 1.55% 86.0 ± 0.75% 86.0 ± 0.00% CC033/GeniUncJ 21.7 ± 1.77 g 82.8 ± 0.59% 85.4 ± 1.27% 84.7 ± 0.86% CC036/UncJ 25.7 ± 1.75 g 85.4 ± 0.66% 85.2 ± 1.87% 84.5 ± 1.57% CC037/TauUncJ 22.8 ± 0.80 g 84.7 ± 2.15% 84.8 ± 0.15% 85.3 ± 0.90% CC040/TauUncJ 32.9 ± 3.60 g 86.1 ± 0.10% 81.7 ± 1.35% 79.4 ± 0.25% CC041/TauUncJ 24.9 ± 1.42 g 88.1 ± 0.84% 85.3 ± 0.92% 85.8 ± 0.13% CC043/GeniUncJ 20.1 ± 0.00 g 84.5 ± 1.05% 84.4 ± 2.85% 86.1 ± 2.45% CC051/TauUncJ 32.9 ± 3.27 g 85.2 ± 0.49% 83.5 ± 0.88% 82.9 ± 0.32% CC057/UncJ 28.3 ± 2.25 g 83.0 ± 0.70% 82.4 ± 0.05% 84.0 ± 0.30% CC059/TauUncJ 35.1 ± 2.60 g 83.9 ± 1.00% 82.0 ± 0.15% 82.6 ± 0.50% CC061/GeniUncJ 21.8 ± 0.00 g 87.4 ± 0.00% 85.1 ± 0.00% 87.2 ± 0.00% CC075/UncJ 25.9 ± 1.70 g 85.8 ± 0.15% 81.4 ± 0.20% 81.7 ± 0.20% CC078/TauUncJ 25.7 ± 2.05 g 87.7 ± 1.25% 87.7 ± 3.00% 86.1 ± 1.25% CC079/TauUncJ 21.2 ± 3.70 g 85.3 ± 0.60% 83.8 ± 0.75% 84.2 ± 0.40% CC080/TauUncJ 20.1 ± 5.05 g 84.0 ± 1.05% 86.5 ± 0.05% 84.0 ± 0.10% CC084/TauJ 28.7 ± 2.55 g 83.7 ± 0.15% 83.0 ± 0.65% 83.7 ± 0.60% Note: SEM is represented as ± the mean. TABLE 3. Average of strains' body weight during each stage of delay discounting testing, represented as a percentage of their initial starting weight (grams) Mouse ID Start 0 s 3 s 6 s 9 s NOD/ShiLtJ 27.4 ± 1.98 g 89.0 ± 1.13% 89.3 ± 1.10% 88.7 ± 0.75% 88.9 ± 1.07% 129S1/SvImJ 22.5 ± 0.78 g 89.8 ± 1.42% 87.7 ± 1.17% 90.1 ± 1.39% 89.8 ± 1.61% A/J 21.7 ± 0.54 g 88.9 ± 1.32% 90.8 ± 1.19% 91.3 ± 0.61% 91.0 ± 0.96% C57BL/6J 24.2 ± 1.20 g 90.2 ± 1.23% 85.5 ± 0.86% 89.5 ± 1.32% 88.7 ± 0.95% NZO/H1LtJ 39.7 ± 1.15 g 88.6 ± 1.07% 89.2 ± 1.16% 89.8 ± 0.89% 87.8 ± 0.95% CAST/EiJ 15.5 ± 0.56 g 90.0 ± 1.32% 89.8 ± 0.77% 91.5 ± 1.06% 90.5 ± 0.88% WSB/EiJ 17.2 ± 0.58 g 89.1 ± 1.15% 89.3 ± 1.13% 88.2 ± 1.04% 90.3 ± 1.35% PWK/PhJ 16.0 ± 0.57 g 91.4 ± 0.94% 86.8 ± 0.75% 89.7 ± 0.79% 88.2 ± 1.31% CC002/UncJ 26.0 ± 0.79 g 87.8 ± 0.85% 90.5 ± 1.25% 88.2 ± 1.26% 88.5 ± 0.93% CC004/TauUncJ 26.9 ± 1.14 g 87.8 ± 0.96% 87.6 ± 1.03% 88.3 ± 1.26% 87.8 ± 1.23% CC006/TauUncJ 24.0 ± 0.77 g 88.7 ± 1.65% 90.7 ± 1.51% 90.0 ± 0.76% 87.0 ± 1.41% CC011/UncJ
留言 (0)