
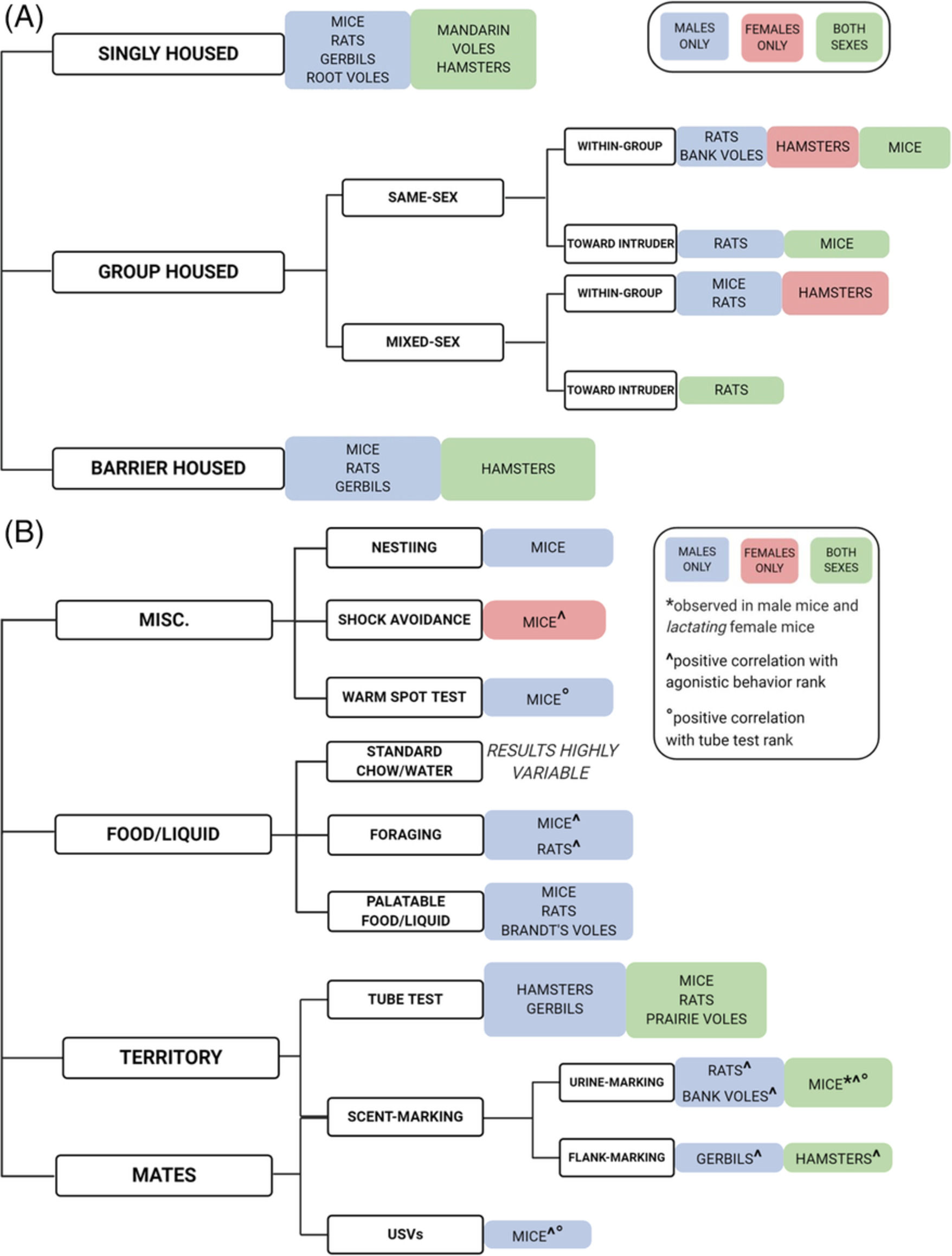
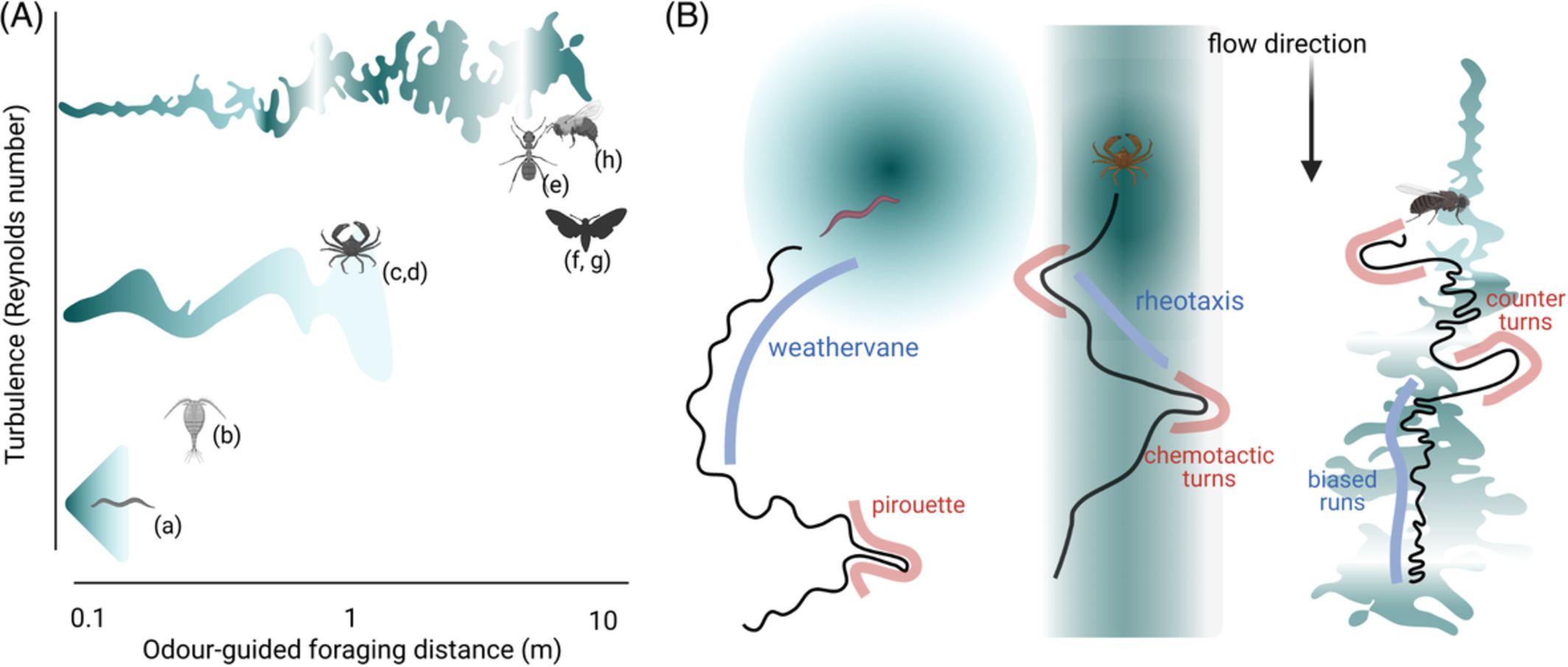
記住我




Multiple sclerosis (MS) is a central nervous system disease characterized by inflammation and myelin sheath injury.1 Learning and memory disorders are the most common symptoms of MS.2, 3 Accumulating evidence has shown that abnormalities in hippocampal microstructures, volume reduction, and extensive myelin loss are associated with learning and memory impairment in MS patients.4, 5 However, the exact mechanism of demyelination-associated cognitive impairment remains unclear.
The myelin sheath is a multi-layered membrane, unique to the nervous system. Approximately half the volume of the human brain is composed of myelinated axons that facilitate rapid and efficient signal transmission central to human cognitive capacities.6, 7 In the central nervous system, myelin is a multi-layered dense membrane structure enclosed by oligodendrocyte (OL) cell membrane outside the axons of neurons. Myelin plays an important role in insulating, improving the conduction speed of nerve impulses, and providing nutritional support for nerve axons.8 Demyelination of axons leads to considerable pathophysiological changes that result in axonal dysfunction, degeneration, and loss of sensory and motor functions. Impaired myelination in the hippocampus has been documented in various neurological diseases including MS, Alzheimer's disease, and depression.5, 9 It has been shown that impaired myelination and neurodegeneration in the hippocampus lead to cognitive deficits.10, 11
Myelin basic protein (MBP) and myelin-associated glycoprotein (MAG) are proteins believed to be important in the process of myelination of nerves in the nervous system. It was found that the expression of MBP in the hippocampus is age-dependent, tissue-specific, and it decreases with time, while the neurons in the hippocampus are not affected, suggesting that MBP leads to age-related cognitive decline.12 Like Alzheimer's disease, rheumatoid arthritis shows cognitive impairment which is associated with higher levels of autoantibodies against MBP leading to the damage of myelin integrity.13 The myelin sheath of central neurons is composed of OL membrane. OL regeneration has emerged as a promising new approach for the treatment of demyelinating disorders. A disintegrin and metalloproteinase protein17 (ADAM17), also known as TNF-α-converting enzyme (TACE), is essential for OL development by modulating OL progenitor cells (OPCs) expansion, cell cycle exit, and survival.14 ADAM17 is responsible for shedding epidermal growth factor receptor (EGFR) ligands and activating EGFR on OL surfaces known to play a role in neuronal remyelination.14, 15 These results show that ADAM17 may be involved in the regeneration of OL and the remyelination of central neurons. iRhoms, highly conserved inactive proteases, belong to the rhomboid family that includes the iRhom1 and iRhom2 subtypes.16 iRhoms regulate membrane protein transportation and proteasome activity in the endoplasmic reticulum.17, 18 Li et al. demonstrated that iRhom1 mRNA was easily detected but iRhom2 mRNA were very low in the brain.19 In addition, iRhom1 is mainly expressed in neurons, astrocytes, and OLs, while iRhom2 is mainly expressed in microglia.19 The study verifies that mature ADAM17 in iRhom1−/− mice is barely detectable in different parts of the brain like hippocampus, cortex and cerebellum in sharp contrast to wild-type mice. They conclude that iRhom1 is not essential for mouse development but regulates ADAM17 maturation in the brain, while ADAM17 maturation in microglia is controlled by iRhom2.19 Another study indicates that iRhom1 is essential for ADAM17 maturation in mouse embryonic fibroblasts.18 Moreover, iRhom1 is one of crucial upstream regulators of ADAM17 and the EGFR signaling pathway.19 However, a role of iRhom1 during cuprizone-induced myelin loss and memory dysfunction has never been addressed. We currently tested expression of iRhom1/ADAM17/EGFR axis in cuprizone-treated mice. More importantly, we evaluated the effects of iRhom1 overexpression in the hippocampus on preventing cognitive injury in cuprizone-induced MS.
2 MATERIALS AND METHODS 2.1 AnimalsMale C57BL/6J (20–22 g) mice were obtained from the Experimental Animal Center of Anhui Medical University. Animals were housed in a controlled environment with a temperature of 22 ± 2°C and humidity of 60% under a 12 h light/dark cycle in group of 3–4. Food and water were available ad libitum. All animal procedures were approved by the Committee for Experimental Animal Use and Care of Anhui Medical University.
2.2 ChemicalsPrimary antibodies of MBP (lot: AG09017659), ADAM17 (lot: AG09254178, EGFR (lot: AG08079087), and CC1 (lot: AH01101936) were purchased from Bioss, Beijing. Primary antibodies of β-actin (lot: 18AW0412) were purchased from ZSGB-Bio. Primary antibodies of MAG (lot: 55 K8768) were purchased from Affinity. Primary antibodies of iRhom1 (lot: 13S5892) were purchased from Biorbyt. Primary antibodies of p-EGFR (lot: I03280683) were purchased from Wanleibio. Horseradish peroxidase-conjugated secondary antibodies were obtained from ZSGB-Bio. Other general agents were commercially available.
2.3 Establishing the demyelination mouse modelExperimental demyelination was induced by feeding 8-week-old male wild-type C57BL/6 mice a diet containing 0.2% cuprizone mixed with a ground standard rodent chow. The cuprizone diet was maintained for 7 weeks. The mice were assigned into a control group or a cuprizone group (n = 12 for each group). The body weights of mice at 1, 2, 3, 4, 5, 6, and 7 weeks of cuprizone treatment were measured.
2.4 Establishing the model of mouse with iRhom1 overexpression in the hippocampusFor assessing the function of iRhom1 on myelin sheath around the hippocampus, pHBAAV2/9-CMV-m-iRhom1-3flag 1 μl (1.0 × 1012 vg/ml) was injected into the hippocampus of 4-week-old male C57BL/6J mice. After the injection, the animals were weighed and then anesthetized by intraperitoneal injection of 4% chloral hydrate at a dose of 10 ml/kg. The heads of the anesthetized animals were then shaved and prepared. The mice were fixed on a brain stereotaxic instrument and the head was sterilized with a 75% alcohol cotton ball. The scalpel was placed along the midline of the skull to make a 1.5 cm incision. The tip of the micro injector was positioned to the halogen door and this position was used as the starting position, 2.0 mm backward, and then 1.5 mm moving the micro sampler is opened side by side. One microliter of iRhom1 overexpressing AVV was injected into the hippocampus of the left and right sides of the brain at a depth of 1.8 mm. After the injection, the iodophor was applied to the wound and sutured. The mice were removed from the brain stereotaxic apparatus and placed in a cage. For GFP expression analysis, mice were injected with 1 μl pHBAAV2/9-ZsGreen (1.0 × 1012 vg/ml; n = 7). For HBAAV2/9 vector assessment, mice were injected with 1 μl pHBAAV2/9 (1.0 × 1012 vg/ml; n = 7). Four weeks later, mice with iRhom1 overexpression in the hippocampus and HBAAV2/9 mice were fed 0.2% cuprizone for 7 weeks. The body weights of mice at 1, 2, 3, 4, 5, 6 and 7 weeks of cuprizone treatment were measured. Cuprizone feeding was discontinued after 7 weeks. The mice were either perfused for immunostaining analysis, Western blotting or Real Time PCR.
For the recovery study, after 6 weeks, cuprizone feeding was discontinued. The mice were returned to a regular diet and allowed to recover for 3 weeks prior to tissue analysis. The body weights of mice at 1, 2, 3, 4, 5, 6, 7, 8 and 9 weeks of cuprizone treatment were measured. Each group consisted of at least four mice for immunostaining analysis and three mice for Western blotting.
2.5 Morris water maze testThe Morris water maze (MWM) task was conducted to measure mouse long-term learning and memory function as described previously.20 Morris water maze is a black barrel-like experimental device with a diameter of 120 cm at the bottom and a height of 50 cm at the top. The bottom is divided into four quadrants I, II, III and IV. The platform is placed in the middle of the third quadrant. The spatial acquisition phase lasted for 5 days, and mice were trained on water maze with four trials per day. A trail lasted for 90s or until the mouse reached the hidden platform and remained on the platform for 15 s. If the mouse did not reach the platform in 90s, it was gently guided and allowed to rest for 15 s. On the 5th day, mouse locus, distance, and time were collected by a behavioral analysis system. On the 6th day, the platform was removed, and mice were tested with a probe trial. The mice were then put into the water from the quadrant I and the mice loci and the platform crossing times recorded within 90 seconds. The distance and speed of the mice in the quadrant III were analyzed. The swimming speed, time in the target quadrant, escape time, and path length were recorded.
2.6 Open field testThe open-field experiment tested the spontaneous activity and exploratory ability of mice. An open-field movable box used in this experiment is a square opaque white plastic box. A camera, which can take pictures of mice loci, is installed above the open-field box. The mice were put into the open-field box for 3 min to adapt to the environment. The experiment was then performed for another 3 min after which the box was cleaned with 75% alcohol after each run. The locus and the number of erection times were recorded during the experiment.
2.7 Brain section preparationAfter the last behavioral test, mice were intraperitoneally anesthetized and then transcardially perfused with 0.9% saline followed by 4% paraformaldehyde using a perfusion pump. The brains were removed, dissected in the mid-sagittal plane, post-fixed overnight at 4°C, dehydrated in a series of graded ethanol solutions and finally embedded in paraffin. Coronal sections at 5 μm were serially cut using a paraffin slicing machine, or the frozen slicer was pre-modulated at −20°C and the blade mounted and its angle adjusted to the desired position. The slice thickness was set to 20 μm and the OTC-embedded sample tissue was adhered to the tissue tray with an OTC embedding agent and then mounted on the advancement. The positions of the sample and the blade were adjusted and the rocker was shaken with a moderate amount of force to cause slicing. The patch action should be fast and light and the tissue patch should be free of breakage, bubble and it should be flat and wrinkle-free.
2.8 Luxol fast blue stainingLuxol fast blue (LFB) staining was carried out for the analysis of myelin injury in the hippocampus. The sections were incubated with the 0.01% LFB solution at 60°C overnight. Then, sections were alternately immersed in 70% ethanol and lithium carbonate solution for differentiation. The slices were rinsed repeatedly with double steaming water. Finally, the slides were rinsed with distilled water, dehydrated by gradient dehydration of anhydrous ethanol, and sealed with neutral gum. LFB staining was used to statistically analyze the areas of pigment loss in the hippocampus of mice using Image-Pro Plus software.
2.9 Immunofluorescence stainingThe tissue sections were rinsed three times with 0.01 mol/L PBS for 5 min each time, and the sections were incubated with 5% BSA for 1 h at 37°C. After 5% BSA was decanted, the prepared primary antibody was added dropwise to the sections and placed in a wet box at 4°C overnight. After overnight incubation, the primary antibody solution was removed and the sections rinsed three times with PBS for 5 min each time. FITC or CY3-labeled secondary antibody was added and further incubation for 2 h at room temperature was done. The sections were then rinsed in PBS three times for 5 min at each time. DAPI was then added and incubation was done for 10 min under darkness at room temperature. This was followed by another cycle of rinses with PBS, three times for 5 min each. A fluorescent anti-quenching agent, glycerine, was subsequently added. Finally, the sections were placed under an upright fluorescence microscope and photographed, and the fluorescence intensity in the images analyzed using Image J Pro Plus software.
2.10 Western blottingAbout 50 mg of brain tissue was homogenized in a homogenizer containing 400 ul of lysate, 10% phosphatase inhibitor and 10 ul of PMSF. For ADAM17 protein, the lysis buffer contained 1 μM Batimastat. After being lysed for 30 min on ice, the samples were centrifuged at 12,000 r/min at 4°C for 15 min, the supernatant was separated. Protein concentration was measured using the BCA kit. A 1:2 mixture of the lysates with Laemmli sample buffer (Bio-Rad, Hercules, CA) was heated for 10 min at 100°C. The proteins were resolved by sodium dodecyl-sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Bedford, MA), and incubated with a blocking buffer (Tris-buffered saline/Tween 20 [TBST]/5% nonfat dry milk) overnight at 4°C. Immunoblot was done with the mouse antibody (1:1000) raised against iRhom 1, MBP, MAG, ADAM17, EGFR and p-EGFR followed by the appropriate horseradish peroxidase (HRP)-conjugated goat anti-mouse antibody (1:10,000). Immunodetection was then performed using the enhanced chemiluminescence system (ECL, Pierce, Rockford, IL) using hydrogen peroxide and luminol as substrates.
2.11 Real-time PCRAnimal brain tissues were collected, total RNA was extracted by TRIzol method, and cDNA was synthesized by reverse transcription according to the instructions of the reverse transcription kit, followed by PCR amplification. β-actin was used as an internal reference. Predesigned Quantitect primers were used for all genes except for β-actin. Primers sequences for β-actin are as follows: Forward:5′-CATTGCTGACAGGATGCAGAA-3′; Rreverse:5′-ATGGTGCTAGGAGCCAGAGC-3′. Set the cycle temperature to 95°C for 30 s, 95°C for 5 s and 60°C for 34 s, 40 cycles in total. Measure the number of cycles (cyclethreshold, Ct) that the fluorescence signal in each reaction tube undergoes within the set range, value and analyze the results according to the 2–ΔΔCt method.
2.12 Statistical analysisAnalysis of variance (ANOVA) for data analysis using the statistical program SPSS 17.0 (Chicago, IL) software was utilized. Data related to escape latency in the MWM test were analyzed using two-way ANOVA with repeated measures followed by Bonferroni or Dunnett's T3 post hoc test. Other data were analyzed by one-way ANOVA. Data are expressed as means ± SEM. p < 0.05 was considered statistically significant.
3 RESULTS 3.1 Up-regulated expression of iRhoms/ADAM17/EGFR axis in cuprizone-treated miceTo investigate the effect of iRhom1 on demyelination-linked cognitive impairment, induction of acute demyelination with cuprizone is performed by feeding adult mice a diet of 0.2% cuprizone.21 In our study, we found that mice treated with cuprizone showed significant body weight loss, cognitive impairments as well as decreased locomotor activity by MWM task and OFT test (Figure S1). Besides, brain sections were stained for myelin fiber and MBP (indicator for OLs) to assess the impact on demyelination after cuprizone exposure. LFB staining analysis was employed to detect myelinated hippocampal fibers. As shown in Figure S2A-B, cuprizone significantly caused injury to the myelin sheath, as thinner myelinated axons were observed in this group when compared with mice in the normal group (t(2) = 6.009, p < 0.05). Immunofluorescence staining for MBP protein showed that significant myelin loss occurred as evidenced by the reduction in MBP immunoreactivity relative to control (t(2) = 5.293, p < 0.05) (Figure S2C). This was further confirmed by Western blot assays for MBP (t(2) = 13.53, p < 0.01) and MAG proteins (t(2) = 7.643, p < 0.05) (Figure S2E-F). In addition, Olig2 (OPC marker) mRNA level was augmented (t(5) = 14.62, p < 0.01) by cuprizone. These results confirmed MS model was established successfully.
Next, we observed the expression of iRhom1 in the brain of cuprizone-treated mice. We found that treatment of 7-week cuprizone up-regulated iRhom1 expression when compared with controls (normal diet-fed mice) (Figure 1A, (t(2) = 20.71, p < 0.01). iRhom 1 is a key regulator of mature-ADAM17, while ADAM17 is ubiquitously expressed in the central nervous system (CNS) and has key functions in myelination of neurons.15, 22 Therefore, nerve lysates were prepared and subjected to Western blot analysis to access the relationship between cuprizone-induced myelin loss and ADAM17. Higher amounts of ADAM17 were observed when compared with the normal diet-fed mice group (Figure 1B, t(2) = 9.187, p < 0.05). Besides, ADAM17 expression is responsible for shedding EGFR ligands and activating EGFR on OL involved in remyelination of neurons.14, 15 In the present study, higher amounts of p-EGFR (activated EGFR) protein expression were observed when compared with controls (normal diet-fed mice) by Western blot assay (Figure 1C, t(2) = 6.077, p < 0.05).

Up-regulation of the iRhom1/ADAM17/EGFR axis in the brain of cuprizone-treated mice. (A). iRhom1 protein expression (t(2)=20.71, p < 0.01) was compared between the two groups. (B). expressions of ADAM 17 (t(2)=9.187, p < 0.05) proteins were detected in the two groups. (C). p-EGFR protein expression (t(2)=6.077, p < 0.05) was compared between the two groups. (D). iRhom2 protein expression (t(2)=4.402, p < 0.05) was compared between the two groups. “*” Denotes statistical significance when compared with controls. Data are expressed as mean ± SEM
It is accepted that iRhom2 has a role in neuroinflammation and neuroinflammatory diseases.23 In addition, MS is a chronic inflammatory disease.24 Therefore, iRhom2 protein expression was detected by Western blot. As a result, iRhom2 protein expression was upregulated under the cuprizone treatment (Figure 1D, t(2) = 8.63, p < 0.05). Taken together, enhancement of the iRhoms/ADAM17/EGFR axis observed following cuprizone-induced demyelination suggested that iRhoms may be involved in myelination.
3.2 The model of mice with iRhom1 overexpression in the hippocampusiRhom1−/− mice have almost no mature and functional ADAM17 in the brain, which is essential for OL development,14, 19 and iRhom1 is perhaps more likely to play a role in physiological processes in the brain.23 Therefore, to investigate iRhom1 function on myelination in cuprizone-fed mice, the aav plasmid was employed to establish mice with iRhom1 overexpression in the hippocampus in vivo. First, to verify a successful iRhom1 overexpression in the hippocampus mouse model, we injected 1 μl each of pHBAAV2/9-ZsGreen (1.0 × 1012 vg/ml) and pHBAAV2/9-CMV-m-iRhom1-3flag (1.0 × 1012 vg/ml) plasmids into the hippocampus. Four weeks later, images were obtained from the mice transfected with the pHBAAV2/9-ZsGreen plasmid to verify precise transfection into the hippocampus (Figure 2B). By Western blotting, iRhom1 protein expression was found to be highly expressed in the hippocampus of mice when compared with adeno associated virus (aavnull group), and iRhom1 mRNA level was consistent by real time PCR (Figure 2C,F, F2,18 = 25.51, p < 0.001). To verify whether iRhom1 overexpression in the hippocampus activates ADAM17/EGFR signaling pathway, protein expression and mRNA level of ADAM17 were detected by western blot and real time PCR. The results demonstrated that compared with aavnull group, iRhom1 overexpression in the hippocampus in the setting of cuprizone enhanced protein expression and mRNA level of ADAM17 (Figure 2D,G, F2,18 = 26.47, p < 0.001), as well as p-EGFR expression (Figure 2E, F2,18 = 15.43, p < 0.01). While aavnull group showed no effect on protein expressions of iRhom1, p-EGFR and ADAM17 compared with control group (Figure 2). To further verify whether iRhom1 overexpression in the hippocampus could improve OLs (CC1 as OLs marker) expressing ADAM17 and p-EGFR, immunofluorescence double staining was used. The image showed that iRhom1 overexpression in the hippocampus could augment amount of OLs as well as promote OLs expresses ADAM17 and p-EGFR proteins compared with aavnull group (Figure 2H,I). These data confirmed that the iRhom1 overexpression in the hippocampus mouse model was successfully established and specific iRhom1 overexpression in the hippocampus facilitated ADAM17/EGFR signaling pathway.

The model of mouse with iRhom1 overexpression in the hippocampus. (A). Timeline depicts the experimental design and time points of analysis. (B). After 4 weeks of injection, frozen sections showed viral infection in the hippocampus. Protein expressions of iRhom1 (C). (F2,18 = 25.51, p < 0.001), ADAM17 (D). (F2,18 = 26.47, p < 0.001), and p-EGFR (E). (F2,18 = 15.43, p < 0.01) were compared among the three groups. mRNA levels of iRhom1 (F). and ADAM17 (G). were detected. Immunofluorescence double staining for CC1/ADAM17/DAPI (H). and CC1/p-EGFR/DAPI (I). was detected. “*” Denotes statistical significance when compared with controls (n = 7, **p < 0.01, ***p < 0.001). iRhom1over is abbreviated as iRhom1 overexpression in the hippocampus. Data are expressed as mean ± SEM. One-way ANOVA, Bonferroni test
3.3 Specific iRhom1 overexpression in the hippocampus rescued cuprizone-induced cognitive dysfunctionDuring the 7 weeks of iRhom1 overexpression in the hippocampus mice treatment with cuprizone, body weight was measured. As expected, iRhom1 overexpression in the hippocampus significantly altered body weight loss induced by cuprizone. We observed a significant increase in body weight in cuprizone-fed mice with iRhom1 overexpression in the hippocampus when compared with cuprizone-fed WT mice, and no role of adeno associated virus (aavnull group) on body weight was observed (Figure 3A). Interestingly, mice with iRhom1 overexpression in the hippocampus could rescue cuprizone-induced cognitive dysfunction. Mice with iRhom1 overexpression in the hippocampus could rescue cuprizone-induced prolongation of escape latency (F2,18 = 11.62, **p < 0.01, △p < 0.05) and quickly find the platform after training (Figure 3B,C, F2,18 = 30.66, ***p < 0.001, △△△p < 0.001) and could spend longer durations in the target quadrant when compared with the cuprizone-fed mice by MWM task assay (Figure 3D,E). Besides, iRhom1 overexpression in the hippocampus improved the exploratory ability of cuprizone-fed mice by OFT test (Figure 3I-L): Moving Distance (F2,18 = 17.19, **p < 0.01, △△p < 0.05); Number of Stand up (F2,18 = 27.1, **p < 0.01, △△p < 0.01); Center Moving Distance (F2,18 = 12.47, **p < 0.01, △△p < 0.01); Center Moving Distance ratio (F2,18 = 13.36, **p < 0.01, △p < 0.05). However, MWM task assay showed no difference in velocity among these groups by MWM task assay (Figure 3F). These data suggested that specific iRhom1 overexpression in the hippocampus could reverse poor cognitive capability and exploratory ability induced by cuprizone.

iRhom1 overexpression rescues cuprizone-induced cognitive dysfunction. (A). The body weights of mice at 1, 2, 3, 4, 5, 6, and 7 weeks of cuprizone treatment were measured. (B–H). Behavioral changes were tested by the Morris water maze test: (B). Escape Latency was measured from the 1st to the 5th day at the 7th week. (C). Swimming trajectory was shown on the 1st and the 5th day at the 7th week. (D). Times of crossing the platform (F2,18 = 11.9, **p < 0.01, △p < 0.05); (E). Time in target quadrant (F2,18 = 30.66, ***p < 0.001, △△△p < 0.001); (F). Swimming velocity. (G). Distance in target quadrant (F2,18 = 10.16, **p < 0.01, △p < 0.05); (H). Escape Latency (F2,18 = 11.62, **p < 0.01, △p < 0.05). (I-L). Behavioral changes were tested on the 7th week using the open field test: (I). Moving Distance (F2,18 = 17.19, **p < 0.01, △△p < 0.05); (J). Number of Stand up (F2,18 = 27.1, **p < 0.01, △△p < 0.01); (K). Center moving distance (F2,18 = 12.47, **p < 0.01, △△p < 0.01); (L). Center Moving Distance ratio (F2,18 = 13.36, **p < 0.01, △p < 0.05). CPZ is abbreviated as cuprizone. “*” Denotes statistical significance when compared with aavnull group (n = 7, **p < 0.01, ***p < 0.001). △denotes statistical significance when CPZ + iRhom1over group compared with cuprizone-fed mice transfected with the blank virus (n = 7, △p < 0.05, △△p < 0.01, △△△p < 0.001). iRhom1over is abbreviated as iRhom1 overexpression in the hippocampus. Data are expressed as mean ± SEM. One-way ANOVA, Bonferroni test
3.4 iRhom1 overexpression in the hippocampus rescued cuprizone-induced myelin lossTo address whether the improvement in cognitive capability and exploratory ability observed after iRhom1 overexpression in the hippocampus were due to alterations in hippocampus myelinated fibers, we investigated the structure of myelinated fibers and myelin-associated protein expressions. Compared with cuprizone-fed mice, cuprizone-fed mice with iRhom1 overexpression in the hippocampus had a similar amount of myelinated fiber structure when compared with normal diet-fed mice by LFB assay (Figure 4A, F2,18 = 16.23, *p < 0.05, △p < 0.05). Similar findings were observed with regards to myelin-associated protein expression. MBP (F2,18 = 24.35, **p < 0.01, △△△p < 0.001) and MAG (F2,18 = 9.399, *p < 0.05, △△p < 0.01) protein expressions were significantly higher in cuprizone-fed mice with iRhom1 overexpression in the hippocampus when compared with cuprizone-fed mice by Western blot assay (Figure 4B). To validate whether the protective function of specific iRhom1 overexpression in the hippocampus on myelin injury is associated with ADAM17/EGFR signaling pathway, expressions of ADAM17 and p-EGFR were observed by Western blot. As a result, both protein expressions of p-EGFR (F2,18 = 20.11, **p < 0.01) and ADAM17 (F2,18 = 18.99, **p < 0.01) (Figure 4B) were highly up-regulated in cuprizone+iRhom1over group, as well as mRNA level of MBP (F2,18 = 4.09, **p < 0.01), MAG (F2,18 = 3.722, *p < 0.05), ADAM17(F2,18 = 12.35, **p < 0.01), EGFR (F2,18 = 10.07, **p < 0.01), and iRhom1 (F2,18 = 5.72, **p < 0.01) were consistent by Real Time PCR (Figure 4C). No effect of adeno associated virus (aavnull group) on iRhom1 protein expression was observed (Figure 4). Furthermore, the apoptosis had been observed by Western blot on p-p53 expression. As a result, iRhom1 overexpression in the hippocampus sustained OLs survival and attenuated apoptosis protein p-p53 expression compared with cuprizone + aavnull group (Figure 4D, F2,18 = 4.23, **p < 0.01). Next, to investigate the effect of iRhom1 overexpression in the hippocampus on cuprizone-induced inflammation, iRhom2 mRNA level was detected by Real Time PCR. The result indicated that iRhom1 overexpression in the hippocampus declined iRhom2 mRNA level (Figure 4E, F2,18 = 3.526, *p < 0.05). These findings suggested that specific iRhom1 overexpression in the hippocampus not only reduced demyelination but also alleviated inflammation in MS.

iRhom1 overexpression in the hippocampus reverses cuprizone-induced myelin loss. (A). Myelin sheath displayed positive LFB staining in the hippocampal CA1 area and statistical analysis (F3,20 = 16.23, *p < 0.05, △p < 0.05). (B). Myelin associated proteins in the brain including MBP (F2,18 = 24.35, **p < 0.01, △△△p < 0.001), MAG (F2,18 = 9.399, *p < 0.05, △△p < 0.01), p-EGFR (F2,18 = 20.11, **p < 0.01), ADAM17 (F2,18 = 18.99, **p < 0.01) and iRhom1 were detected by Western blotting. (C). MBP (F2,18 = 4.09, **p < 0.01), MAG (F2,18 = 3.722, *p < 0.05), ADAM17(F2,18 = 12.35, **p < 0.01), EGFR (F2,18 = 10.07, **p < 0.01), and iRhom1 (F2,18 = 5.72, **p < 0.01) mRNA level were validated by real time PCR. (D). Expression of p-p53 protein was detected by Western blotting. (E). iRhom2 (F2,18 = 3.526, *p < 0.05) mRNA level were validated by Real Time PCR. CPZ is abbreviated as coprizone. *denotes statistical significance when compared with aavnull group (n = 7, *p < 0.05, **p < 0.01). △denotes statistical significance when compared with cuprizone-fed mice transfected with the blank virus (n = 7, △p < 0.05, △△p < 0.01, △△△p < 0.001). iRhom1over is abbreviated as iRhom1 overexpression in the hippocampus
3.5 Cognitive capability improved following removal of cuprizoneTo address whether iRhom1 overexpression in the hippocampus contributes to further improve cognitive capability after cuprizone removal, mice with pHBAAV2/9-CMV-m-iRhom1-3flag injection were fed with a regular diet for 3 weeks after 6 weeks of cuprizone treatment (Figure 5A). As expected, the body weights of mice with iRhom1 overexpression in the hippocampus were similarly with those of normal diet-fed mice during the cuprizone feeding period. However, mice on the cuprizone diet experienced weight loss that developed slowly following the removal of cuprizone (Figure 5B). Next, to address whether there was difference in cognitive capability and exploratory ability among all groups following the removal of cuprizone, the Morris Water Maze and open field tests were used. Interestingly, both cognitive capability and exploratory ability were investigated in all groups. However, no significant differences were detected in behavioral tests (Figure 5C-M).

Cognitive capability improved following the removal of cuprizone. (A). Timeline depicts the experimental design and time points of analysis. (B). The body weights of mice at 1, 2, 3, 4, 5, and 6 weeks of cuprizone treatment and 7, 8, and 9 weeks of cuprizone removal instead of normal diet were measured. (C-I) Behavioral changes were tested by the Morris water maze test: (C). Escape Latency was measured from the 1st to the 5th day at the 9th week. (D). Swimming trajectory was shown on the 1st and the 5th day at the 9th week. (E). Escape Latency; (F). Distance in target quadrant; (G). Times of crossing the platform; (H). Time in target quadrant; (I). Swimming velocity. (J-M). Behavioral changes were tested by the open field test: (J). Moving Distance; (K). Number of Stand up; (L). Center Moving Distance; (M). Center Moving Distance ratio. There were no statistical differences between the three groups of mice. CPZ is abbreviated as coprizone. NS denotes no statistical significance when iRhom1over + CPZ recover group compared with CPZ recover group. “*” Denotes statistical significance when compared with aavnull group (n = 7, *p < 0.05, **p < 0.01). △denotes statistical significance when compared with cuprizone-fed mice transfected with the blank virus (n = 7, △p < 0.05, △△p < 0.01). iRhom1over is abbreviated as iRhom1 overexpression in the hippocampus
3.6 Remyelination developed following removal of cuprizoneAfter behavioral testing, the level of hippocampal remyelination was investigated. Consistent with behavioral results obtained, no significant difference was observed in all groups as seen after LFB staining (Figure 6A). Besides, similar results occurred in MBP and MAG protein levels by Western blot assays (Figure 6B,C). The level of hippocampal remyelination among all groups reached that of the control animals. These data indicated that hippocampal remyelination developed following removal of cuprizone. iRhom1 overexpression showed no synergistic effect on remyelination at 3 weeks after cuprizone removal. This result may be related to no measurable long-term loss in myelination in control mice treated with cuprizone or the short observation time after cuprizone removal. Therefore, we will continue to extend the cuprizone treatment time or observation time points of CNS remyelination after cuprizone removal in the following work.

Remyelination occurred following the removal of cuprizone. (A). Myelin sheath displayed positive LFB staining in the hippocampal CA1 area. Myelin associated proteins in the hippocampus including MBP (B), MAG (C) were detected among the three groups using Western blotting. There were no statistical differences among the three groups of mice studied (n = 7). CPZ is abbreviated as coprizone. iRhom1over is abbreviated as iRhom1 overexpression in the hippocampus. NS denotes no statistical significance when iRhom1over + CPZ recover group compared with CPZ recover group
4 DISCUSSIONMS is a chronic inflammatory autoimmune disease of the central nervous system.25 Learning and memory dysfunction has been shown to be a common symptom in MS patients and is associated with hippocampal neuron demyelination, but the exact mechanism remains opaque.2, 13 Our results showed that hippocampal iRhom1 overexpression significantly inhibited cuprizone-induced decrease in hippocampal MBP and MAG contents in mice. In addition, iRhom1 overexpression also reduced hippocampal demyelination and alleviated cognitive and locomotor activity impairments in cuprizone-induced MS model, suggesting that iRhom1 is implicated in the demyelination-related cognitive impairment.
iRhom1, a member of the rhomboid family, occurs in the endoplasmic reticulum.26 This protein functions as an oncogene in multiple cancers, but little is known about its function in the brain.27 Previous studies demonstrated that iRhom1−/− mice have almost no functional ADAM17 in the brain, which is essential for OL development,14, 19 and iRhom1 is perhaps more likely to play a role in physiological processes in the brain.23 Therefore, we investigated the effect of iRhom1 overexpression on demyelination-linked cognitive impairment in cuprizone-induced MS mice model. We found that hippocampal myelin content was significantly decreased, and the structure of hippocampal myelin was damaged in MS mice brain. In addition, mice treated with cuprizone showed significant learning and memory impairments as well as decreased locomotor activity. These are similar to findings in the previous study.28 These results confirmed MS model was established successfully. Next, we over-expressed iRhom1 in the hippocampus to evaluate the role of iRhom1 on cuprizone-induced cognitive impairment and locomotor activity. We found that hippocampal iRhom1 overexpression significantly reversed cuprizone-induced body weight loss, cognitive dysfunction, and decline of locomotor activity in mice, suggesting that iRhom1 played an important role in improving cognitive capability in cuprizone-induced MS model.
Next, we further investigated the mechanism of iRhom1 protecting cognition from cuprizone damage. Cuprizone leads to selective death of OLs which results in the demyelination of white matter in the brain.29 In the present study, we found that cuprizone treatment reduced MBP (as mature OL indicators) and MAG contents in the brain of mice, which could be reversed by hippocampal iRhom1 overexpression. In addition, iRhom1 overexpression in the hippocampus could restore the defects in OL cell survival and diminish the apoptosis protein p-p53 expression, which attributed to prevent cuprizone-induced OLs apoptosis and substantially alleviated cuprizone-induced hippocampal demyelination in the hippocampus. These data suggested iRhom1 could prevent demyelination induced by cuprizone. ADAM17 activation can promote OL regeneration and CNS remyelination, suggesting a primary role of ADAM17 in MS progression.14, 30 Li et al. demonstrates that iRhom1 plays an important role in regulating ADAM17 maturation in the brain,19 but little is known about the function of ADAM17 and iRhom1 in the brain. Also, deficiency of ADAM17 resulted in the inhibition of EGFR activation, and overexpression of EGFR seems to be enough to make up for the slow remyelination induced by lack of ADAM17.14, 15 Our results demonstrated that hippocampal iRhom1 overexpression significantly up-regulated the expressions of ADAM17 and p-EGFR. Taken together, iRhom1 overexpression might improve cognitive impairment induced by cuprizone via regulating ADAM17/EGFR signal pathway to ameliorate OLs loss.
MS is a chronic, inflammatory, demyelinating neurodegenerative disease.31 As one membership of inactive Rhomboid-like proteins iRhoms, iRhom2 is expressed in immune cells, mostly macrophages and the macrophage-like microglia in the brain, but little or no iRhom2 is expressed in the rest of brain (i.e., neurons and glial cells).19 iRhom2 has a role in neuroinflammation and neuroinflammatory diseases.23 Therefore, iRhom2 expression was investigated under the cuprizone treatment. The result indicated that iRhom2 expression was elevated in MS mice, while hippocampal iRhom1 overexpression significantly downregulated cuprizone-induced iRhom2. Altogether, these results suggest hippocampal iRhom1 overexpression not only reduced demyelination but also alleviated inflammation in MS.
Interestingly, iRhom1 expression was up-regulated by cuprizone, but the damage trend was continuing in the present study. The reasons may be as follows. Specific targets of cuprizone are mature OLs, which fail to fulfill the extensive metabolic demand and consequently undergo apoptosis to lead to demyelination,23, 32 which was consistent with our finding. Therefore, cuprizone-induced high expression of iRhom1 was impossible presented in the mature OLs which were loss in the context of cuprizone. Increasing evidence demonstrates that cuprizone can increase the number of OPCs.30 In our study, we also found that Olig2 (OPC marker) mRNA level was augmented in the setting of cuprizone, suggesting that OPCs were accumulated. Therefore, we speculated up-regulation of iRhom1 expression induced by cuprizone may be associated with increased OPCs. Indeed, new oligodendrocytes are generated from OPC to form new myelin sheaths.31 Despite a large amount of OPCs, the morphology of immature OLs with cuprizone treatment was less developed.33 As a result, up-regulated iRhom1 induced by cuprizone showed no effect on preventing myelin loss, while may serve as a reserve for myelin repair which was preliminarily investigated in this study.
Once demyelination is completed, new oligodendrocytes, generated from the pool of OPC, begin to form new myelin sheaths soon after cuprizone removal.31 Remyelination mainly mediated by OPCs restores over 60% of hippocampal myelin as well as learning and memory.30 In the context of remyelination, high levels of ADAM17 or EGFR promote the process of oligodendrogenesis.14, 34 We found that up-regulation of iRhom1 promoted the maturation of ADAM17 and p-EGFR levels in the context of cuprizone, which might contribute to adequately modulating OL regeneration and remyelination after cuprizone removal so that there was no significant difference of remyelination between cuprizone group and cuprizone+ iRhom1 overexpression group at 3 weeks after cuprizone removal. Generally, the remyelination of the central nervous system was investigated at 1 or 3 weeks after CPZ removal.14, 33 Thus, in our study, we chose to observe the CNS remyelination at 3 weeks after CPZ removal. Demyelination is usually induced by cuprizone treatment for 3 ~ 6 weeks.22, 35 Therefore, in our study, we treated the mice with cuprizone for 6 weeks. Our finding suggested that hippocampal iRhom1 overexpression highly improved cuprizone-induced myelin injury but could not enhance remyelination after cuprizone removal. Indeed, this result may be related to no measurable long-term loss in myelination in control mice treated with cuprizone or the short observation time after cuprizone removal. Next, we will continue to extend the cuprizone treatment time or observation time points of CNS remyelination after cuprizone removal in the following work.
Our results unambiguously demonstrate that iRhom1 overexpression exerted be
留言 (0)