
記住我
Vaccination is the main pillar of the worldwide effort to contain and ultimately end the ongoing severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) pandemic. Although multiple efficacious vaccines have been developed at unprecedented speed, the number of failed or underwhelming vaccine programs illustrates the challenges in mounting a protective immune response against SARS-CoV-2. The ideal vaccine antigens should direct the immune response to the most vulnerable structure on virion surfaces: the viral spike protein in the prefusion conformation.
The SARS-CoV-2 spike protein catalyzes the fusion of virions with host cells in a highly coordinated manner. The protein is synthesized as a single polypeptide chain that is proteolytically cleaved by a furin protease into subunit 1 (S1) and subunit 2 (S2) during virus assembly. The receptor-binding domain (RBD) in S1 mediates attachment to angiotensin-converting enzyme 2 (ACE2) that is expressed on the surface of the host cell, whereas S2 contains critical elements required for fusion. Virions display functional spike trimers in a metastable prefusion conformation (i.e., a long-lived state that has not reached the favorable energy minimum). ACE2-receptor binding stabilizes RBDs in the receptor-accessible “up” configuration. Although the molecular mechanism and the order of events during SARS-CoV-2 fusion have not been elucidated in detail, it is clear that fusion requires further proteolytic maturation of S2 by cell-surface or endosomal proteases, as well as dissociation of the S1 subunit from S2. After these steps, the hydrophobic fusion peptide of S2 is thought to be inserted into the membrane of the host cell. The structural changes that follow allow trimeric S2 to adopt a lower energy conformation that brings the viral membrane into close contact with the host cell membrane. This ultimately results in fusion of the two membranes.
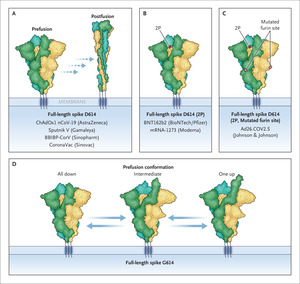
Figure 1. Figure 1. Strategies to Stabilize Prefusion Conformation of the SARS-CoV-2 Spike Protein.
Figure 1. Strategies to Stabilize Prefusion Conformation of the SARS-CoV-2 Spike Protein. Approved vaccines against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) rely on the original D614 spike variant and encode or contain full-length, unmodified spike proteins, which are prone to adopt the postfusion conformation even in the absence of angiotensin-converting enzyme 2 (Panel A), or spike protein mutants that stabilize the prefusion conformation in an all–receptor-binding domains (RBDs)–down configuration (Panels B and C). These spike mutants include the full-length spike with the mutations K986P and V987P (labeled “2P”) (Panel B) and the full-length spike 2P with mutated furin cleavage site (R682S and R685G, labeled “mutated furin site”) (Panel C). The full-length G614 spike exhibits a stable prefusion conformation that can reversibly adopt an all-down (all RBDs in the down state), intermediate (one RBD in an intermediate state), or one-up (one RBD in an up state) configuration and thus constitutes a promising vaccine antigen (Panel D).
During the lifetime of SARS-CoV-2 virions and in laboratory preparations of trimeric spike protein, a substantial fraction of spike proteins prematurely undergoes these changes spontaneously, in the absence of ACE2 and without catalyzing fusion. Because these steps are irreversible, spike proteins in the postfusion conformation are no longer able to catalyze fusion, which reduces infectivity. Importantly, the spike protein is the major target of neutralizing antibodies in infected or vaccinated people. The prefusion conformation of the spike protein represents the most relevant target, because antibodies can successfully interfere with infection only if they prevent binding to target cells or prevent fusion itself. Most vaccine designs therefore include modifications to amino acids in the spike protein. For example, substitutions with proline stabilize the prefusion conformations and alter the furin-cleavage site (Figure 1).1,2
In late February 2020, a point mutation in the SARS-CoV-2 spike protein, the D614G variant, was identified. The presence of this mutation predicts the substitution of the amino acid aspartate at position 614 (D614) with glycine (G614). A few months later, this variant virus had become the dominant form, which suggests that D614G endows the virus with an evolutionary advantage. It is notable that G614 is found in all current variants of concern. Earlier work has suggested that G614 allows the RBD of the spike protein to more readily adopt configurations in which one of the RBDs is in the up state — a configuration that is compatible with ACE2 binding.3
In a recent study, Zhang et al.4 reported the exact structure of spike trimers containing D614 or G614. In most previous studies, the researchers solved the structures of a stabilized, secreted spike version: the transmembrane domain was replaced with an artificial trimerization domain, and stabilizing mutations were introduced to prevent spike cleavage and conformational changes (the 2P mutations2). Zhang et al. solved the structures of the unmodified spike protein, including the transmembrane domain. They solubilized the molecule with the help of detergents — arguably, the closest one can get to the native structure without preserving the lipid bilayer. The D614 protein was less stable, and a substantial fraction of the molecules assumed the postfusion conformation, with S1 detached from the trimeric spike. In contrast, G614 was predominantly found in the prefusion conformation with hardly any signs of premature activation. Accordingly, a larger fraction of G614 spike proteins on the surface of virions is in the prefusion conformation, which renders virus particles more infectious.
To understand the molecular origin of this beneficial stabilization, the authors compared the structural details of the two variants. The G614 spike trimers were found in one of three configurations: all RBDs in the down state, one RBD in an intermediate state, or one RBD in an up state. The latter two configurations were not frequently adopted by the D614 spike, perhaps because RBD-up configurations in D614 rendered the spike more likely to undergo the full transition to the postfusion conformation. Importantly, the relatively small amino acid glycine at position 614 allowed a loop containing amino acid 630 to adopt a folded structure that fits into a small cleft near the interface of S1 and S2. This structural element was not observed in the D614 structure and contributes to the stabilization of the prefusion conformation. By providing additional interactions, the folded “630 loop” prevented S1 shedding and more generally kept spike trimers in the fusion-competent prefusion conformation. This loop must unfold in at least two of the three spike protomers to expose one RBD in the up configuration, a requirement for ACE2 binding. Adoption of the postfusion conformation probably requires unfolding of all three 630 loops. Hence, kinetic barriers separate the different configurations, prolonging the sampling time of the different prefusion conformations and minimizing the risk of accidentally triggering the irreversible changes that lead to the postfusion conformation.
SARS-CoV-2 variants containing the G614 mutations are equally vulnerable to neutralization by therapeutic monoclonal SARS-CoV-2 spike antibodies or to neutralization by patient sera after infection or vaccination.5 The pronounced stabilization of the prefusion conformation of the G614 spike with no RBD or one RBD in the up position provides an evolutionary advantage because of the increased infectivity of virions; Zhang et al. have provided a molecular explanation for this phenomenon. The G614 spike would be a formidable antigen for future vaccines, because premature conversion to the postfusion conformation is limited and because it is prone to adopting the relevant one-RBD–up configuration.
留言 (0)