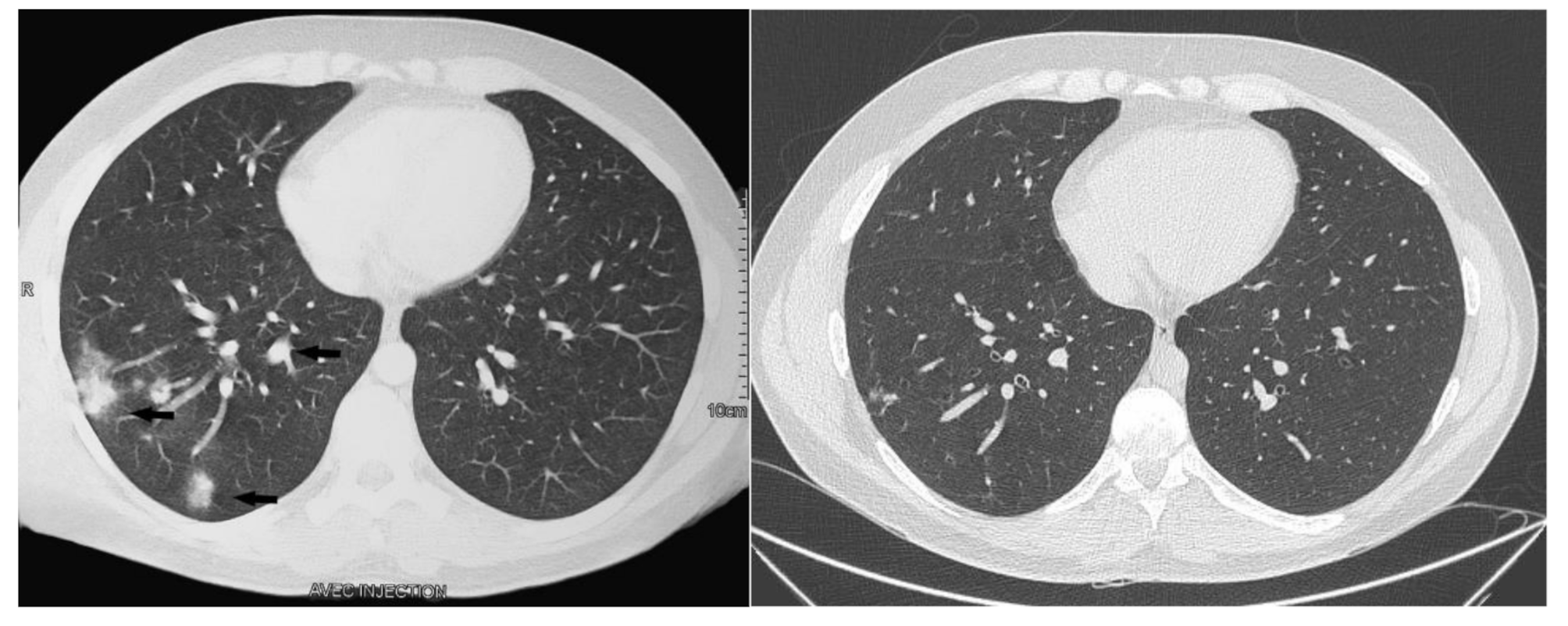
We describe a proven human case of cryptococcosis in a patient with an isolated NKD. The patient’s clinical and radiological presentation was compatible with pulmonary cryptococcosis for an immunocompetent or mildly immunocompromised patient. Radiological manifestations of pulmonary cryptococcosis may include a lung mass (ranging from a simple nodule to a large cavitary mass) to multiple pulmonary nodules, as in our patient, to an interstitial process indicating widespread disseminated infection. The spectrum of clinical presentations is broad and depends on the host’s immune status. Most frequent are cough, chest pain, sputum production and fever, although asymptomatic cases are described [2, 16].
We investigated the patient’s anti-Cryptococcus defenses and demonstrated the persistence of a very low but functional NK compartment associated with a secondary impaired adaptive anticryptococcal CD8 + T cell immunity and the presence of a huge Vβ14 CD8 clonotype. Cryptococcosis has not been described among patients with isolated NKD. However, NKD and cryptococcosis have been described during more complex deficiencies such as AIDS [9], idiopathic CD4 + T-lymphocytopenia [19], and more recently in a young male with C. neoformans and Varicella Zoster meningitis and idiopathic hypereosinophilic syndrome, hypocomplementemia and low NK cells [20].
NK cells are non-B non-T-cell CD3−CD56+ lymphocytes and are the third largest contingent of blood lymphocytes (4–15%) [3]. Most circulating NK cells are cytotoxic and express a low level of CD56 as well as an immunoglobulin G Fc receptor, FcγRIIIA (CD16), and are termed as CD56dim NK cells in contrast with the more immature CD56bright NK cells [3]. CD56dim NK cells constitute the majority of peripheral blood NK cells [21, 22] NK cells are located at the interface of innate and adaptive immunity with three main functions: cytotoxicity, chemokine and cytokine production, and the regulation of immunity through contact-dependent costimulatory and regulatory mechanisms [3, 4, 15]. In infectious diseases, NK cells act against certain elusive T-cell pathogens—particularly Herpesviridae—that negatively regulate MHC class 1–infected cells [3]. NK cells are negatively regulated by MHC class I antigens, while they are triggered by many activation receptors. NK cells are implicated in eliminating pathogens such as viruses but are also involved in the immune response against Cryptococcus spp [7, 8]. NK cells directly kill Cryptococcus using the NKp30, β1-integrin and NKG2D activating receptors [9, 11]. In human pathology, NKD syndrome is divided into classical and functional deficiencies [3]. Our patient had a classic NKD with a stable defect over time and an opportunistic infection that was unlike any previously described NKD cases. NK cells account for nearly 10% of the lung lymphocyte population. NKD may, therefore, favor fungal lung infections such as infection with Cryptococcus, which penetrates the body through the respiratory tract.
The residual NK cells displayed normal NKp30 receptors, IFN-gamma production, degranulation, and perforin expression. However, severe NK cell lymphopenia could lead to a global loss of cytotoxicity, which could have affected the destruction of fungi dependent on perforin, as previously described [4]. NK cells do not only exhibit direct target destruction but also act as regulators of other immune agents via cytokine synthesis and are thus of utmost importance to mounting the robust Th1 response necessary to fight Cryptococcus infection [14]. Our patient had an impaired CD8 T cell response to Cryptococcus. The patient’s T cells did not proliferate after in vitro stimulation by a cryptococcal antigen suspension. Cytotoxic CD8 cells play an important role in the host’s immune response to C. neoformans [5, 7]. This defective CD8 response appeared related, i.e. secondary to the primary NKD, resulting in a weak IFN-γ production, leading to an impaired DC IL-12 production and an impaired TH1 response against C. neoformans [8, 14].
Due to an initial slight increase in the CD8 population, we analyzed TCRVβ, revealing a chronic expansion of a Vβ14 CD8 + clone. The clonotype persisted during the follow-up, though with a slight decrease (the patient stopped his follow-up in 2019). Such clonal CD8 expansion has already been described in healthy subjects and many nonmalignant disease states including immunodeficiencies [23, 24]. These clonotypes do not represent malignant proliferation. In the literature, it is hypothesized that these clonal expansions may reveal reactive CD8 lymphocytes with specificity for the antigens of common pathogens, particularly Herpesviridae—which are responsible for recurrent reactivation [24]. Immunologic memory in these CD8 + T cell clonotypes is necessary to maintain the host pathogenic equilibrium relationship. Still, it can also lead to a reduction in the T-cell repertoire by inhibiting other antigen-specific lymphocytes [24]. NK cells modulate T-cell memory differentiation and regulate and kill recently activated proliferative CD8 + T cells [14]. NKD could, therefore, have led to the persistence of the patient’s CD8 clonotype due to a lack of killing of expanded clonotype cells. The possible negative effect of a preexisting clonotype reducing the NK cell compartment seems less plausible. This phenomenon has not previously been described and should have affected other lymphocyte compartments. Due to the diversity of peptides and HLAs, the antigen responsible for the patient’s Vβ14 clonotype expansion is difficult to assess. We showed that this clonotype, though this measure was performed only once during the follow-up, did not proliferate with a challenge using Cryptococcus antigen.
In conclusion, this case report describes a new type of classical NKD phenotypically characterized by isolated lung cryptococcosis resulting from a complex deficiency affecting both innate (NKD) and adaptive immunity. This case confirms in human pathology the central role of NK cells in Cryptococcus clearance via innate and adaptive immunity.








留言 (0)