
記住我
The ability to discriminate small changes in sensory attributes improves remarkably with practice, a process referred to as perceptual learning (PL). PL has been found to be specific to simple stimulus attributes (e.g., location and orientation) in various visual discrimination tasks (e.g., Ahissar and Hochstein, 1997; Crist et al., 1997; Fahle et al., 1995; Karni and Sagi, 1991; Poggio et al., 1992; for reviews, see Fahle, 2005; Gilbert et al., 2001; Li, 2016; Sagi, 2011). For example, training of discriminating visual stimuli presented at one location of the visual field enhanced behavioral performance significantly, but this learning effect did not transfer to another location, regardless of whether there were distractors presented at this untrained location during the training (e.g., Gutnisky et al., 2009; Karni and Sagi, 1991) or not (e.g., Crist et al., 1997; Fahle et al., 1995). So far, the location specificity of PL has been observed in various visual tasks using different types of stimuli (Ahissar and Hochstein, 1997; Jia et al., 2020; Schoups et al., 1995; Shiu and Pashler, 1992), in both tens of minutes of training (fast learning, e.g., Fahle et al., 1995; Shiu and Pashler, 1992) and extensive training over several days (slow learning, e.g., Harris et al., 2012; Karni and Sagi, 1991; Yotsumoto et al., 2008). The striking specificity of PL to stimulus location leads to a speculation that learning-induced changes take place in relatively early stages of visual cortical areas where neural activities are highly selective for stimulus location (for reviews see Ahissar and Hochstein, 2004; Gilbert et al., 2001; Sagi, 2011; Watanabe and Sasaki, 2015).
Though the location specificity of PL is generally considered as a hallmark of visual PL (e.g., Lu and Dosher, 2022; Lu et al., 2011; Maniglia and Seitz, 2018), the underlying mechanism remains unclear. An important question is whether the location specificity of PL reflects facilitation of processing at the trained location only, or involves suppression of processing at the untrained locations as well. It is notoriously difficult to figure out this question using behavioral measurements, because both learning-induced facilitation and suppression mechanisms would lead to the same behavioral outcomes. For example, in a typical PL study of location specificity (e.g., Ahissar and Hochstein, 1997; Crist et al., 1997; Fahle et al., 1995; Gutnisky et al., 2009; Karni and Sagi, 1991), behavioral performance at the trained location improved significantly, while that at untrained locations did not change or only improved a little after training. Traditionally, such results are attributed to facilitative processing specific to the trained location. However, an alternative hypothesis could not be excluded: although visual discriminability is improved by learning, it could not be reflected at untrained locations due to some sort of learned suppression to those locations. According to literatures, PL may be a complex process (Ding et al., 2003; Fahle, 2005; Ahissar and Hochstein, 2004) involving not only stimulus-specific mechanisms but also general learning mechanisms (such as learning of task rule or task familiarity which are essential for discriminability improvement in a specific visual task; Qu et al., 2014; Zhang et al., 2010). If there is learning-induced suppression to the untrained stimulus attribute (e.g., location or orientation), it would inhibit the expression of the general learning mechanisms, leading to little transfer of learning effects on behavioral performance in the untrained stimulus condition. Behavioral and ERP studies have shown evidence supporting that besides stimulus specificity, generalization is also common in PL (Liu and Weinshall, 2000; Qu et al., 2014). For example, in our previous fast PL studies, while behavioral results showed typical orientation-specific learning effects, ERPs revealed not only specific effects but also generalized effects (as reflected by different ERP components; Ding et al., 2003; Song et al., 2007). Through double training or training-plus-exposure paradigms (e.g., Xiao et al., 2008; Xiong et al., 2016; Zhang et al., 2010; Zhang and Yang, 2014), researchers proposed that learning specificity may result from under-activations of untrained visual neurons responding to untrained locations, and training visible stimuli or exposing invisible stimuli at untrained locations may activate the untrained visual neurons, leading to transfer of learning at untrained locations (Xiong et al., 2016). However, evidence is lacking with regard to whether the untrained locations are merely not activated (while the trained locations are activated) or even suppressed during the training. Till now, it remains unclear, whether location-specificity of PL involves both facilitation of processing at the trained location and suppression of processing at the untrained location.
Here we propose that it’s possible to dissociate these two mechanisms through neural measures. Specifically, ERP measurement with its high temporal resolution and reasonable spatial resolution may provide key insights into the underlying mechanisms driving behavior (e.g., Bao et al., 2010; Ding et al., 2003; Fahle and Skrandies, 1994; Hu et al., 2019; Qu et al., 2010; Qu et al., 2014; Qu et al., 2017; Xi et al., 2020; Zhang et al., 2015). One can record the ERPs evoked by targets and distractors during the training, respectively, and to examine whether they are modulated by learning: if the early ERP activity evoked by targets (at trained location) increases and that evoked by distractors (at untrained location) decreases significantly, it will support that both facilitation of target (or trained location) and suppression of distractor (or untrained location) are induced by PL.
If both facilitation of the trained location and suppression of the untrained location are induced by PL, another important question arises: whether they involve similar cognitive neural processes. Specifically, does the learning effect reflect changes of a voluntary attentional deployment, or an involuntary visual processing? Manipulating attentional settings when measuring the learning effects may help to dissociate these two mechanisms. If a learning effect (facilitation or suppression) reflects changes of voluntary attentional deployment, it will hold when the attentional setting in measurements is similar to the training phase and will disappear when the attentional setting is altered. By contrast, if a learning effect reflects an automatic/involuntary process, it will survive across different attentional settings in measurements. Using this method, we recently revealed that shape specificity of PL (i.e., PL effects specific to the trained non-salient shapes; Hu et al., 2019; Qu et al., 2017) involves both mechanisms; that is, learning induced an initial involuntary attentional capture followed by a later voluntary attentional deployment. To our knowledge, however, such a method has not been systematically applied in investigating the location specificity of PL. So far, only a few studies have reported fast learning effects on early visual ERPs. For example, our previous study (Wang et al., 2010) found that the amplitude of the P1, an early visual ERP component appearing around 100 ms at lateral occipital sites, increased with tens of minutes of training in a difficult visual task. It remains unclear, however, whether such fast PL effects on early ERPs were specific to the trained location and whether these effects reflect changes in voluntary attentional deployment or automatic/involuntary processes.
Combining behavioral and ERP measurements, here we aim to investigate the cognitive neural mechanism of location specificity of fast visual PL. In Experiment 1, we investigated whether fast visual PL induces two kinds of location-specific effects at early stages of visual cortical processing, including both facilitation of trained location (i.e., the target or attended location during training) and suppression of untrained location (i.e., the distractor or unattended location during training). In Experiment 2, we further examined whether fast PL-induced facilitation of trained location and suppression of untrained location (both found in Experiment 1) reflect changes in voluntary attentional deployment or involuntary/automatic processing. To get reliable location-specific PL effects on early ERPs and track these changes across training at both the trained and untrained locations, we employed a new approach involving analysis of contralateral-minus-ipsilateral P1 responses.
2 Experiment 1To examine whether fast PL induces both facilitation of the trained location and suppression of the untrained location, we developed an ERP experimental design that combined a sustained spatial attention paradigm (Hillyard and Anllo-Vento, 1998; Mangun and Hillyard, 1987) and fast PL paradigm of Vernier task (Fahle et al., 1995; Poggio et al., 1992). Throughout the tens of minutes of training, subjects were required to discriminate the direction of Vernier offset in one visual-field location (i.e., the trained location) and ignore Vernier stimuli in the other location (i.e., the untrained location). In addition to behavioral performance at the trained location, ERPs elicited by Vernier stimuli as targets at the trained location and as distractors at the untrained location were recorded, which allowed us to observe learning-induced changes of early visual processing at these two locations. In particular, we want to examine whether early ERPs at the trained location increased with training while those at the untrained location decreased with training. Such a design could also allow us to reveal whether and how spatial attentional effect on early ERPs (i.e., ERP difference between the trained and untrained locations) is modulated by fast learning.
2.1 Methods 2.1.1 Subjects and apparatusWe looked to the existing literature that has examined the early ERP learning effects (e.g., Bao et al., 2010; Wang et al., 2010; Wang et al., 2016) or the sustained spatial attention effects in early ERPs (e.g., Berchicci et al., 2019; Clark and Hillyard, 1996) for guidance, which helps to estimate the necessary sample size for the present experiment. Sixteen right-handed subjects (9 female; ages 18–22 years) with normal or corrected-to-normal vision participated in Experiment 1. All were compensated for their participation, either with payment or credit hours fulfilling a course requirement. The study was conducted according to the Code of Ethics of the World Medical Association (Declaration of Helsinki) and informed written consent was obtained from each subject before the beginning of the experiment.
Experiment 1 was conducted in a dimly lit and sound-attenuated booth. All stimuli were generated and scripted using a MATLAB toolbox Psychtoolbox-3 (Brainard, 1997). Visual stimuli were presented on a 17-inch CRT monitor (Dell), with a resolution of 1,024 × 768 and a refresh rate of 60 Hz. Auditory stimuli were presented by a pair of loudspeakers placed at the left and right sides of the monitor. Subjects’ head position was stabilized with a chin rest at a viewing distance of 100 cm.
2.1.2 Stimuli and taskIn Experiment 1, subjects participated in a four-block training of a simple Vernier task (Figure 1A), in which they were required to discriminate the direction of horizontal offset in the Vernier. As shown in Figure 1B, the Vernier stimulus comprised a pair of identical sinusoidal gratings [diameter = 2.75°, contrast = 0.8, spatial frequency = 3 cpd, orientation = vertical, and a center-to-center distance = 2 λ (1 λ ≈ 0.34°)] on a gray background (mean luminance, 9.86 cd/m2). The position of the lower grating was offset to the left or right of the upper grating with equal probabilities, and the offset size was determined by the psychophysical test before training for each subject. In each “training” block, the Vernier appeared either in the upper left visual field (LVF) or the upper right (RVF) at 5° retinal eccentricity. Specifically, Vernier’s center was located 2.11° above the horizontal line of the screen and 4.53° from the vertical center line. Each Vernier was presented for 50 ms, with a 1,100–1,300 ms interval between successive stimuli (Figure 1B). To avoid repeated appearance of stimuli at the same location for more than 3 consecutive trials, the stimuli were presented at the trained or untrained location in a pseudo-random order. A small black cross in the center of a screen (0.3° × 0.3°; 0.23 cd/m2) was present throughout the block to help with the fixation. Subjects were required to judge whether the lower grating was to the left or right of the upper grating in one visual hemifield (i.e., the trained location) throughout training and ignored the other hemifield (i.e., the untrained location). They were instructed to respond as accurately and quickly as possible. Auditory feedback was given after behavioral responses. The trained location and the untrained location were counterbalanced across subjects; that is, half of them were required to attend the LVF throughout training, while the other half were required to attend the RVF. The two locations were completely matched for stimulus display but only differed in spatial attention settings with such a design. Therefore, any differential electrophysiological responses between these locations could be attributed to the effect of spatial attention and/or fast learning. Subjects received four training blocks, each containing 640 trials (i.e., 320 trials for the attended locations and 320 for the unattended location).
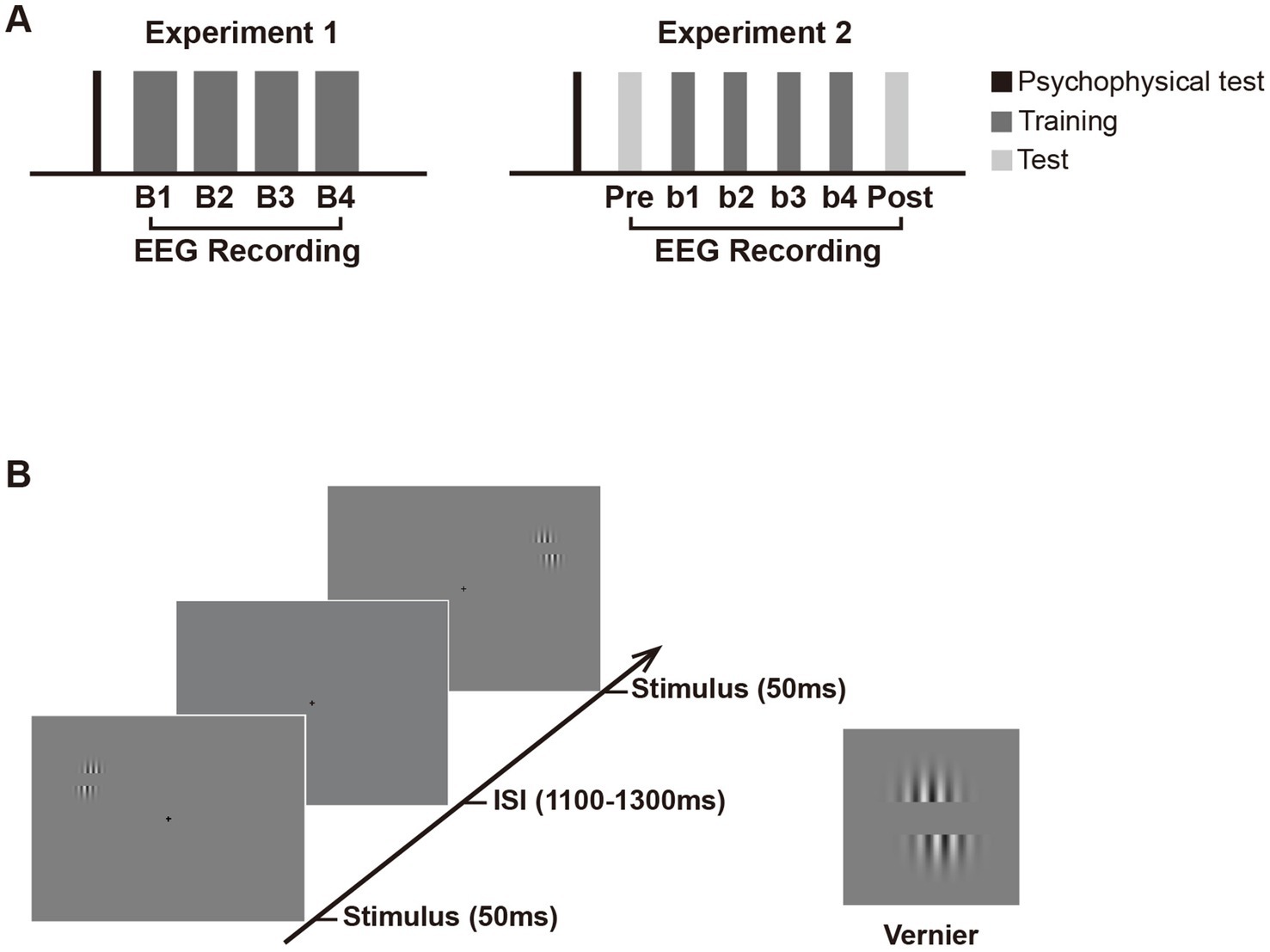
Figure 1. Schematic presentation of the experimental design. (A) Experimental procedure. Experiment 1 consisted of a psychophysical test and four training blocks (B1 to B4) with EEG recording. Experiment 2 consisted of an EEG test before (pretest) and after training (posttest), in addition to similar psychophysical test and training blocks as in Experiment 1. Note that in Experiment 2, the number of trials in each training and test block is half of that in each block in Experiment 1 to avoid fatigue effect. (B) Trial sequence in the training and test phase. In both experiments, Vernier stimulus was briefly flashed (50 ms) at the left or right visual fields with equal probabilities. The inter-stimulus intervals (ISIs) ranged from 1,100 to 1,300 ms. Subjects were required to judge whether the lower grating was to the left or right of the upper grating in one visual hemifield (i.e., the trained location) throughout training and ignore the other hemifield (i.e., the untrained location). They were to respond as accurately and quickly as possible once the Vernier stimulus appeared. If the position of the lower grating was offset to the left of the upper grating, participants were required to press ‘1’ on the numeric keypad; if the lower grating was offset to the right of the upper, they pressed ‘3’. Auditory feedback was given immediately after behavioral response (lower pitched sound for correct response and higher pitched sound for incorrect response). In Experiment 2, the Test and training blocks used identical stimulus displays and only differed by the instructions given to subjects. In the pretest and posttest, subjects were instructed to discriminate the Vernier offset at the untrained location and ignore that at the trained location.
Different from the classic Hillyard sustained spatial attention paradigm (e.g., Clark and Hillyard, 1996; Di Russo et al., 2003; Martinez et al., 1999), the present design required subjects to respond to each stimulus at the attended location and used a longer inter-stimulus interval (ISI). Through such a design, stable behavioral measures of fast learning could be obtained through a large number of trials, and sufficient time were provided to subjects to prepare for the next stimulus. The present design also brought up a problem that greater slow-wave activities from preceding trials (e.g., motor-related ERPs) might overlap with the early ERPs of current trials. Although a jitter of ISI could mitigate the overlapping to certain extent, it could not completely rule out the possibility of contamination from overlapping. To solve this problem, we employed a new approach using analysis of contralateral-minus-ipsilateral ERPs to minimize these overlapping (slow-wave activities) and to get reliable PL effects (For more details, see “2.1.4 Data analysis”).
Each subject was given a psychophysical test before training to determine the individual’s Vernier offset size. This test was separately implemented in the LVF and RVF. For the LVF test, the Vernier was briefly flashed only in LVF (50 ms), followed by a period of 1,600–2,000 ms during which subjects were required to discriminate the Vernier offset. The LVF test contained four blocks of 8 trials, with decreasing Vernier offset from 6 to 3 pixels (1 pixel ≈ 0.018°); that is, the offset sizes were 6 (≈0.108°), 5 (≈0.09°), 4 (≈0.072°), and 3 (≈0.054°) pixels for these four blocks, respectively. The procedure of the psychophysical test for the RVF was identical to that in the LVF, except that Vernier stimuli were only presented in the RVF in each trial. The test order (i.e., test in the LVF first vs. test in the RVF first) was counterbalanced across subjects. For each subject, behavioral performance at each offset size was collapsed across the LVF and RVF for response accuracy calculation. Then the Vernier offset size for which subjects’ discrimination accuracy fell at 75% was regarded as the fixed stimulus parameter used in the subsequent training blocks. The whole experiment (the psychophysical test and training) lasted for about 1.8 h (the training phase lasted for about 1.6 h), including participants’ breaks between blocks.
2.1.3 EEG recordingThe EEG was recorded from 57 scalp sites using the 10–10 system montage. Standard 10–20 sites were FP1, FPz, FP2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8, O1, Oz, and O2. Additional intermediate sites were AF3, AFz, AF4, F5, F1, F2, F6, FC5, FC3, FC1, FCz, FC2, FC4, FC6, C5, C1, C2, C6, TP7, CP5, CP3, CP1, CPz, CP2, CP4, CP6, TP8, P5, P1, P2, P6, PO7, PO3, POz, PO4, and PO8. All scalp channels were recorded using a common average reference online and were then algebraically re-referenced to the average of the left and right mastoids offline. The horizontal and vertical electrooculogram (EOGs) were monitored with bipolar recordings from electrodes at the left and right outer canthi, and from those above and below the left eye. Electrode impedances were kept below 5 kΩ.
The EEG analog signals were digitized at a 512-Hz sampling rate, and a digital antialiasing filter of 0.27 × the sampling rate was applied at the time of recording. After filtering the EEG signals with a digital 40-Hz low-pass filter and then a 0.1-Hz high-pass filter, epochs were extracted that included 100 ms of pre-stimulus baseline and 600 ms of post-stimulus EEG. Trials contaminated by eye blinks, or muscle potentials exceeding ±70 μV at any electrode were excluded before averaging. Besides, trials with horizontal eye movements exceeding ±30 μV in the bipolar HEOG channel (<1% of trials) were discarded by a step-like artifact detection procedure (moving window width = 200 ms, moving step = 10 ms), so that the observed early ERP effects may not be attributed to sudden shifts in eye position (i.e., saccades) triggered by stimulus onset. After artifact rejection, about 300 trials were left for each location in each block. ERPs were then averaged according to each location within each training block to examine changes in amplitudes of the ERP components with training.
2.1.4 Data analysisAccuracies in each training block were calculated for behavioral data. To examine the learning effect on behavioral performance, the accuracies were separately subjected to one-way ANOVA with the factor being training block (B1, B2, B3, and B4).
In the ERP analyses, we are interested in the early ERP changes originating from early visual cortical areas which might underlie location-specific fast PL. In each training block, early ERPs in response to the Vernier at each location (i.e., the trained or untrained location) were measured to reveal the fast learning effects. We first examined the original ERPs and found some overlapping confounds, some of which are common in fast PL studies (as well as in some visual attention studies; see Baumgartner et al., 2018; Ding, 2018; Qu and Ding, 2018; Slotnick, 2018 for reviews and comments in the special issue “Attentional modulation of early visual areas” edited by Slotnick, 2018). One source of confounds came from slow-wave activities from preceding trials, probably including response-related ERPs and/or anticipatory waveforms like the contingent negative variation, both of which originated from high-level cortical processing (see Supplementary Figure 1). In addition, some location-nonspecific training effects from higher brain areas (e.g., midline P1 effect; see Supplementary Figure 2) might also contaminate the possible location-specific effects in early visual cortical processing. These confounds overlapped with early visual evoked potentials (e.g., the C1 and lateral P1), making it hard to get reliable or interpretable location-specific PL effects on the original ERPs. Since the scalp distributions of these confounds were unrelated to the stimulus locations (e.g., the confounding activities were distributed centrally or bilaterally, with maximum amplitudes at or near the midline sites), analyses on the contra-minus-ipsilateral ERPs may eliminate these confounds to a great extent and get reliable location-specific PL effects on early ERPs (especially those on the early P1 at contralateral sites during 75–120 ms) at the trained and untrained locations, respectively. Thus, contra-minus-ipsilateral ERP waves rather than original ERPs were analyzed and reported in the present study. With such a subtraction, the first positive deflection (75–120 ms) in the contralateral-minus-ipsilateral difference wave would reflect the early visual processing of the stimulus at the trained or untrained location, which was called as ‘P1c-i’ (contra-minus-ipsilateral P1) in the present study for convenience. Through visual inspection of grand-average ERPs, we found that learning-induced P1c-i changes appeared at two different time windows (75–95 ms and 100–120 ms) for the trained and untrained locations respectively, both involving occipital sites (Figures 3C,D), which were consistent with previous findings that the contralateral P1 contains two subcomponents (early P1 and late P1; Di Russo et al., 2003). We then performed statistical analyses to test whether this observation is reliable. Specifically, the mean amplitudes of P1c-i were measured at occipital sites (PO7/8, PO3/4, P7/8, and P5/6) in two distinct time intervals (75–95 ms and 100–120 ms). The mean amplitudes of early P1c-i (75–95 ms) and the mean amplitudes of late P1c-i (100–120 ms) were then separately analyzed in a two-way ANOVA with factors being location (Trained vs. Untrained) and training block (B1, B2, B3, and B4). Two-tailed paired t-tests were further used if necessary.
To characterize the training-induced changes of scalp distribution of lateralized P1 (P1c-i), spline interpolated topographical maps of scalp voltage were calculated for the difference waves between training blocks (e.g., the difference in P1c-i amplitude between B1 and B2). Topographies of lateralized activity were typically projected to both sides of the head to map contralateral-minus-ipsilateral differences in previous studies (e.g., Kiss et al., 2008; Qu et al., 2014). Because such voltage maps are mirror-symmetric, only the left side of the head is highlighted in the present study (see also Störmer et al., 2019).
2.2 Results 2.2.1 Behavioral resultsAs shown in Figure 2A, ~ 90 min of training induced a significant change in accuracy [main effect of training, F (3, 45) = 3.006, p = 0.040, ηp2 = 0.167]. Further analysis revealed a trend of improvement in discrimination accuracy after a single training block of trials (B1 vs. B2 improvement, 0.022 ± 0.012, mean ± SE, p = 0.085, 95% CI = [−0.003, 0.046]). In the fourth training block (B4), however, the accuracy decreased significantly (B4 vs. B2, p = 0.043), indicating fatigue effects associated with the training. To examine the detailed time courses of learning and fatigue effects, each training block was bisected into 2 mini-blocks, resulting in 8 mini-blocks (b1 to b8) in total. As illustrated in Figure 2B, accuracy increased gradually across training and reached the maximum in the fourth mini-block (b4 vs. b1, p = 0.009), while fatigue effects did not reach significant until the last mini-block (b8 vs. b4, p = 0.016).
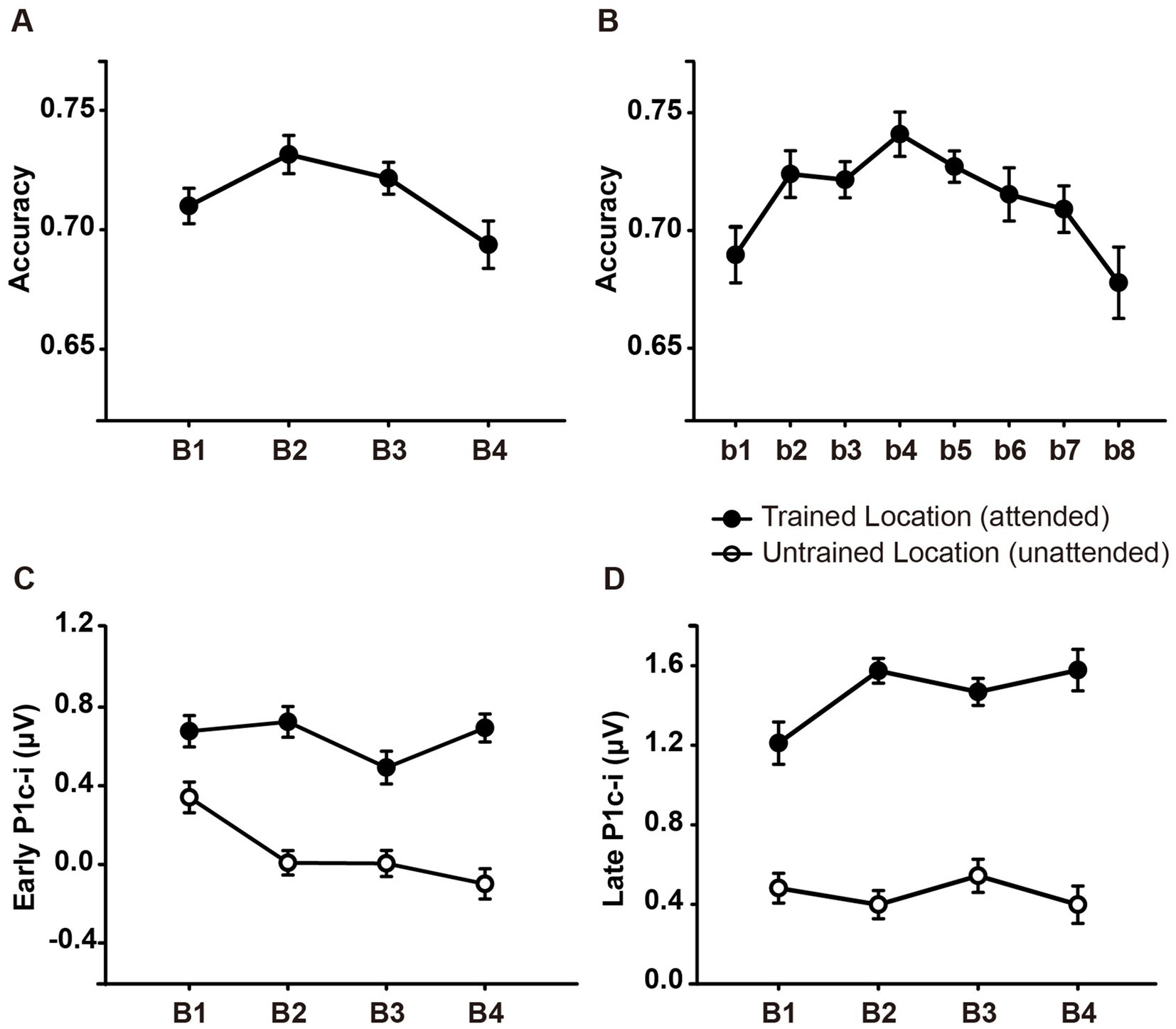
Figure 2. Mean accuracies and ERP amplitudes in each training block of Experiment 1. (A) Vernier’s discrimination accuracy at the trained location improved after a single training block of trials. (B) Each training block was bisected into 2 mini-blocks, resulting in 8 mini-blocks in total. There was a significant fatigue effect in the last mini-block (b8). (C,D) P1c-i amplitudes were measured at posterior electrodes (collapsed across PO7/8, PO3/4, P7/8, and P5/6) in two different time intervals (i.e., 75–95 ms and 100–120 ms). Training decreased early P1c-i amplitude (75–95 ms) evoked by distractors at the untrained (unfilled circles) but not that evoked by targets at the trained location (i.e., filled circles). In contrast, training increased late P1c-i amplitude (100–120 ms) at the trained but not at the untrained location. Error bars indicate within-subject standard errors (Cousineau, 2005).
2.2.2 ERP resultsThe original ERPs (i.e., contralateral and ipsilateral ERPs) of targets at the trained location and of distractors at the untrained location are illustrated in Figures 3A,B. Since the original ERPs contained some overlapping confounds which are common in fast PL and spatial attention studies (see Methods for details), here we measured the contra-minus-ipsilateral wave during the P1 time window, and called the difference wave as ‘P1c-i’ in the present study. Although the subtraction method might also eliminate the earliest component C1 to a large extent due to C1’s approximately midline distribution under the present stimulus condition (Di Russo et al., 2003), it could definitely exhibit differences of early P1 between contra- and ipsi-lateral sites. The P1c-i waves of targets at the trained location and those of distractors at the untrained location (Figures 3C,D) both showed a positive deflection during 75–120 ms, which is consistent with previous findings that the P1 was larger over the contralateral scalp than the ipsilateral scalp during this time window (e.g., Clark and Hillyard, 1996; Di Russo et al., 2003; Figure 1 in McDonald et al., 2022). In each training block, the amplitude of the P1c-i at each location was analyzed to track changes of brain activities during training. As shown in Figures 3C,D, the amplitude of the P1c-i at the trained location increased with training during 100–120 ms, whereas that at the untrained location decreased within 100 ms after stimulus onset.
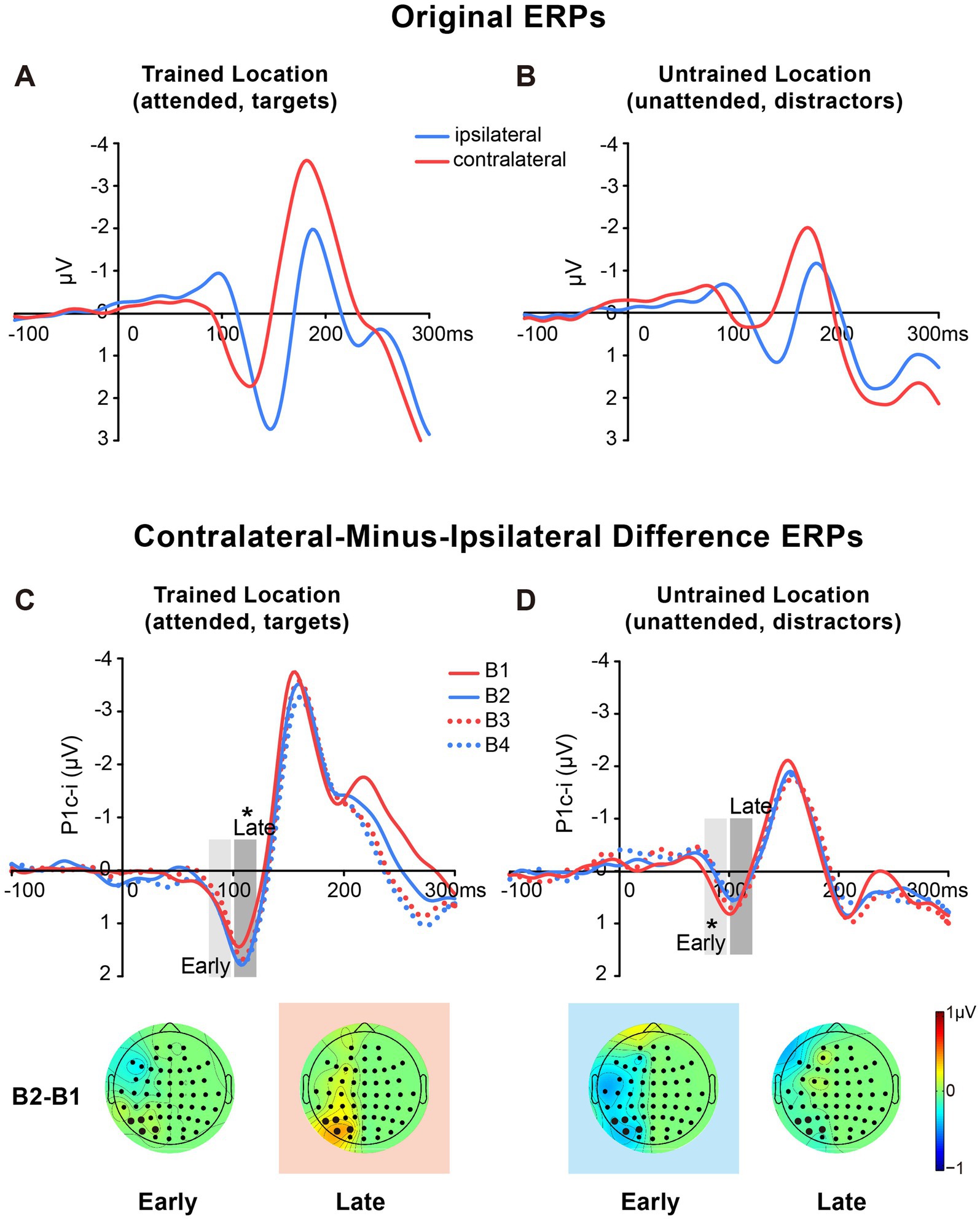
Figure 3. Grand average ERPs and voltage topographies in Experiment 1. (A,B) Original ERP waveforms (averaged across four training blocks) elicited by Vernier stimuli as targets at the trained location (A) and as distractors at the untrained location (B) at electrodes (collapsed across PO7/8, PO3/4, P7/8, and P5/6) contralateral and ipsilateral to the stimulus locations. (C,D) Contralateral-minus-ipsilateral difference waveform, called P1c-i, elicited by targets at the trained location (C) or distractors at the untrained location (D) in each training block (B1, B2, B3 and B4). P1c-i waves were measured in two different time intervals (i.e., 75–95 ms and 100–120 ms), shown as shaded rectangles. Topographical voltage maps of the B2 minus B1 difference amplitude averaged over the 75–95 ms and 100–120 ms time window, respectively. These voltage distributions of difference waves were projected onto the left hemisphere (for details see Data Analysis). In the maps, the electrodes highlighted in bold represent a channel group used for the analysis of learning effects, including PO7/8, PO3/4, P7/8, and P5/6. Red in the topographic plot indicates training-induced larger P1c-i, and blue indicates training-induced smaller P1c-i. * p < 0.05.
2.2.2.1 Early P1c-iStatistical analyses of the P1c-i further confirmed the distinct learning effects at the trained and untrained locations in two time intervals. For the early P1c-i (75–95 ms), a significant interaction of location × training block was found [F (3, 45) = 2.986, p = 0.041, ηp2 = 0.166]. Further analysis showed a training-induced significant decrease in the early P1c-i amplitudes at the untrained location [One-way ANOVA with the factor as training block, F (3, 45) = 5.412, p = 0.003, ηp2 = 0.265; linear trend, F (1, 15) = 16.048, p = 0.001, ηp2 = 0.517]. The decrement of early P1c-i mainly occurred between the first two blocks (B1 vs. B2 decrement, −0.333 ± 0.131 μV, mean ± SE, p = 0.023, 95% CI = [−0.61, −0.05]; Figures 2C,3D), and preserved in the following training blocks (B1 vs. B4, −0.440 ± 0.122 μV, p = 0.003, 95% CI = [−0.70, −0.18]; B2 vs. B4, −0.107 ± 0.106 μV, p = 0.327, 95% CI = [−0.33, 0.12]; Figure 2C). In contrast, there were no significant differences in early P1c-i amplitudes at the trained location across training blocks [F (3, 45) = 1.356, p = 0.268, ηp2 = 0.083; Figures 2C, 3C].
2.2.2.2 Late P1c-iFor the late P1c-i (100–120 ms), a significant interaction of location (Trained vs. Untrained) × training block (B1, B2, B3, and B4) was also found [F (3, 45) = 2.992, p = 0.041, ηp2 = 0.166]. Further analysis showed that training increased the late P1c-i amplitude at the trained location significantly [One-way ANOVA with the factor as training block, F (3, 45) = 2.911, p = 0.045, ηp2 = 0.163; linear trend, F (1, 15) = 3.091, p = 0.099, ηp2 = 0.171]. The increase of late P1c-i mainly occurred after a single training block (B1 vs. B2 increment, 0.364 ± 0.141 μV, mean ± SE, p = 0.021, 95% CI = [0.06, 0.66]; Figures 2D, 3C), and almost preserved in the following training blocks (B1 vs. B4, 0.367 ± 0.187 μV, p = 0.068, 95% CI = [−0.03, 0.77]; B2 vs. B4, 0.004 ± 0.137 μV, p = 0.980, 95% CI = [−0.29, 0.30]; Figure 2D). In contrast, no significant differences in late P1c-i amplitudes were found across training blocks at the untrained location [F (3, 45) = 0.564, p = 0.642, ηp2 = 0.036; Figures 2D, 3D].
As reported above, both the early P1c-i effect at the untrained location and the late P1c-i effect at the trained location occurred mainly within the first two blocks and were preserved in the last block (B4). The preservation of these early ERP learning effects in B4 indicates that they are not sensitive to fatigue, which is different from the behavioral effect. Since behavioral result showed significant learning effect within the first training block (i.e., between b1 and b2), to further examine whether such quick effect also appears on the P1c-i, we divided the first two blocks into 4 mini-blocks (Figure 4). Results showed that, the early P1c-i at the untrained location decreased significantly between b1 and b4 (p = 0.004), but not between b1 and b2 (p = 0.460); similarly, the late P1c-i at the trained location increased significantly between b1 and b4 (p = 0.020), but not between b1 and b2 (p = 0.536). Taken together, the present results showed different time course of early ERP learning effects from that of behavioral effect, indicating that they involve different mechanisms. While the early and late P1c-i effects mainly reflect location-specific PL, the behavioral performance might involve not only specific PL effects but also some general effects associated with training (e.g., task-rule learning and/or fatigue).
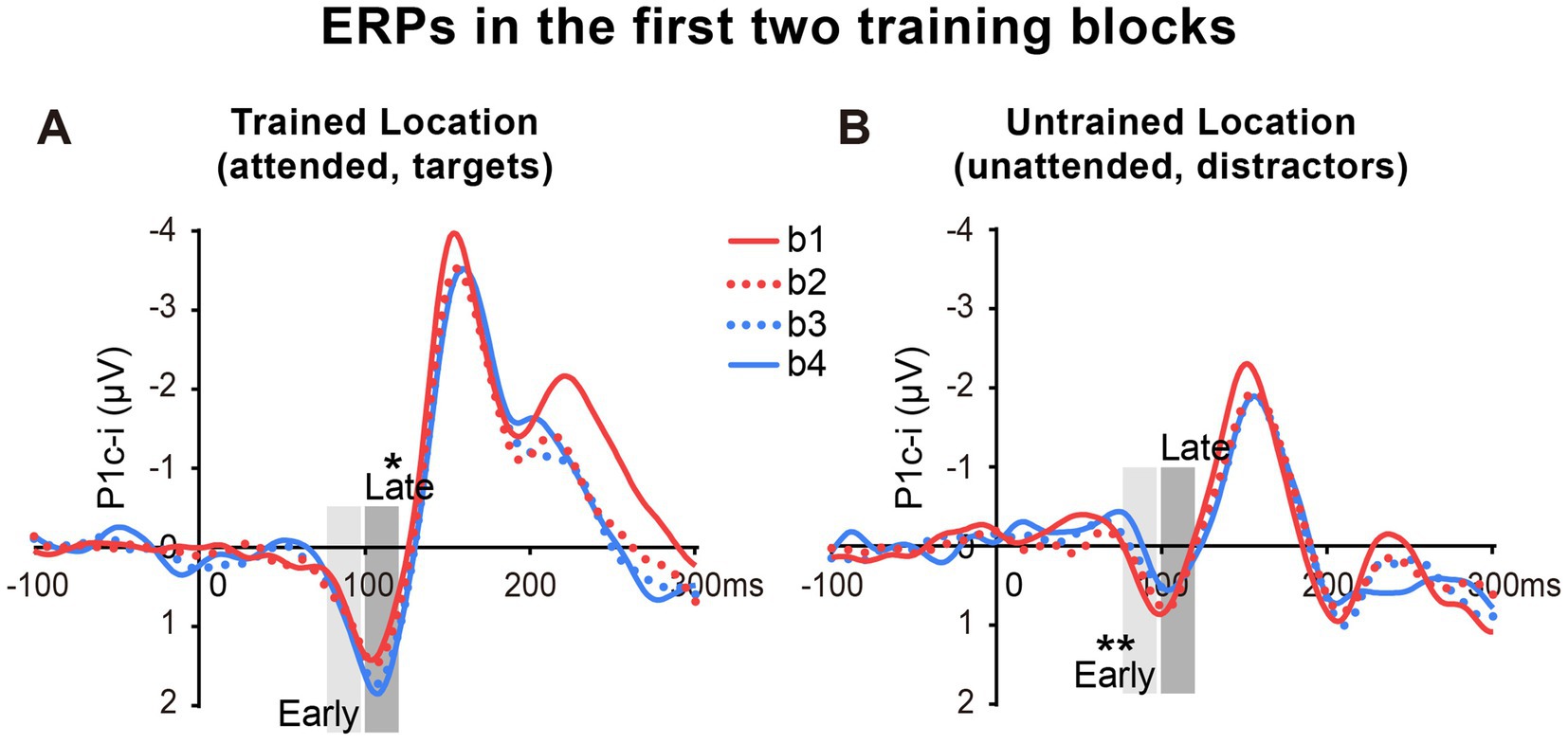
Figure 4. Grand average ERPs in the first two training blocks in Experiment 1. (A,B) The first two blocks were bisected into 4 mini-blocks (b1, b2, b3 and b4) to further examine the detailed time courses of the P1c-i learning effects, especially whether the effects appeared quickly within the first training block (i.e., after a single mini-block b1). The P1c-i elicited by targets at the trained location (A) or distractors at the untrained location (B) in each mini-block were shown. P1c-i waves were measured in two different time intervals (i.e., 75–95 ms and 100–120 ms), shown as shaded rectangles.
2.2.2.3 Inter-trial target/distractor effectTo examine whether the present PL effects on the P1c-i are a consequence of transient inter-trial priming (Britton and Anderson, 2020; Jiang et al., 2013; Wang and Theeuwes, 2018), we further compared the P1c-i effects between conditions when a given stimulus (target or distractor) was preceded by the same stimulus (i.e., repetition condition) or not (i.e., no-repetition condition). Results showed that training decreased the early P1c-i evoked by distractors in both distractor repetition and no-repetition conditions (ps < 0.02), and there was no significant difference for learning effects between these two conditions (p = 0.217). Similarly, for the late P1c-i at the trained location, there was no significant difference for training-induced late P1c-i increment between conditions when targets at the trained location was repeated or not (p = 0.888). These results suggest that inter-trial priming does not play a role in both facilitation of the trained location and suppression of the untrained location.
3 Experiment 2In Experiment 1, we found that tens of minutes of training induced two location-specific effects in early ERPs: an increase of late P1c-i (~100–120 ms) evoked by targets at the trained location and a decrease of early P1c-i (~75–95 ms) evoked by distractors at the untrained location. To confirm these findings and to further examine whether these location-specific fast PL effects reflect changes in voluntary attentional deployment or involuntary/automatic visual processing, we designed Experiment 2. Subjects were given a test block both before (pretest) and after training (posttest), in addition to similar training blocks as in Experiment 1. Test and training blocks used identical stimulus displays and only differed by the instructions given to subjects: while training blocks (i.e., attend-repeat-location blocks) required subjects to discriminate the offset direction of Vernier at the trained location and ignore the untrained location, test blocks (i.e., attend-opposite-location blocks) required subjects to judge the Vernier offset at the untrained location and ignore the trained location. That is, the attentional setting of the test was different from that of the training. The trained location which was the target/attended location during the training became the distractor/unattended location in the test, whereas the untrained location which was the distractor/unattended location during the training became the target/attended location in the test.
In Experiment 2, we first expected to replicate the two PL-induced ERP effects in Experiment 1: an increase of the late P1c-i elicited by targets at the trained location and a decrease of the early P1c-i elicited by distractors at the untrained location. To a great extent, such a replication could avoid the Type I and Type II errors (Luck and Gaspelin, 2017; noted that similar to previous PL studies, early ERP effects observed in Exp. 1 show modest statistical significance, indicating that PL effects on early visual ERPs are not strong; replication would be a better approach than multiple-comparison corrected test to detect such weak signals). Then we examined whether these two effects still exist in the tests. If the target facilitation effect (indexed by the late P1c-i) could not be observed at the untrained location in the test, it will further confirm that this effect is specific to the trained location. Similarly, if the distractor suppression effect (indexed by the early P1c-i) could not be observed at the trained location in the test, it will further confirm that this effect is specific to the untrained location. In addition, since the attentional setting in the test was different from that during the training, if the late P1c-i effect at the trained location or the early P1c-i effect at the untrained location appears during the training but disappears in the test (i.e., the observation of the learning effect is dependent on the attentional setting in measurements), we may infer this learning effect reflects changes in voluntary attentional deployment. In contrast, if these early ERP learning effects appear in both the training and the test (i.e., the observation of the learning effect is independent of the attentional setting in measurements), we may infer they reflect modifications in automatic, involuntary visual processing.
3.1 Method 3.1.1 Subjects and apparatusWe chose the same sample size as that in Experiment 1. Sixteen right-handed subjects (9 female; ages 19–29 years) with normal or corrected-to-normal vision participated in Experiment 2. The apparatus was similar to that used in Experiment 1.
3.1.2 ProcedureStimulus and procedure were similar to those described in Experiment 1 except for the following differences. All subjects were given a test block before (pretest) and after training (posttest) in Experiment 2 (Figure 1A). Test and training blocks used identical stimulus displays and only differed in the instructions given to subjects. In test blocks, subjects were required to discriminate the direction of the horizontal offset of the Vernier stimulus at the untrained location and ignore that at the trained location. The number of trials in each training block (and each test block) were reduced to half of Experiment 1 to minimize fatigue effects since two test blocks were added in Experiment 2 and significant fatigue effects were found in the last block of Experiment 1. Subjects received six blocks (i.e., four training blocks and two test blocks), each block containing 320 trials (i.e., 160 trials for the attended location and 160 trials for the unattended location). The whole experiment (the psychophysical test and training) lasted for about 1.3 h (the training phase lasted for about 1.1 h), including participants’ breaks between blocks.
3.1.3 EEG recordingThe EEG recordings in Experiment 2 were identical to Experiment 1 with the following exceptions. Electroencephalogram (EEG) was recorded in both training and test blocks from 58 scalp electrodes of the 10/10 system. Standard 10–20 sites in the scalp were FP1, FPz, FP2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8, O1, Oz, and O2. Additional intermediate sites were AF3, AFz, AF4, FC5, FC3, FC1, FCz, FC2, FC4, FC6, C5, C1, C2, C6, TP7, CP5, CP3, CP1, CPz, CP2, CP4, CP6, TP8, P5, P1, P2, P6, PO7, PO3, POz, PO4, PO8, I5, I3, Iz, I4 and I6. After artifact rejection, about 145 trials were left in each block for each location (i.e., the attended and unattended locations).
3.1.4 Data analysisIn order to examine learning effects on behavioral performance, accuracies were separately subjected to one-way repeated-measure ANOVAs with the factor being training block (b1, b2, b3 and b4). In addition, a pair-wise t-test was used to evaluate behavioral performance at the untrained location before and after training (i.e., pre- and post-tests) for examining whether the improved performance is specific to the trained location.
ERP mean amplitudes of the P1c-i in response to Vernier stimulus at each location were measured in each block. Consistent with Experiment 1, the mean amplitudes of P1c-i were measured in two distinct time intervals (i.e., 75–95 ms and 100–120 ms) at occipital sites (PO7/8, PO3/4, P7/8, and P5/6). To replicate the two PL-induced ERP effects in Experiment 1, early and late P1c-i amplitudes at each location were then separately subjected to a one-way ANOVA with factors being training block (b1, b2, b3, and b4), respectively. Pair-wise t-tests between pre- and post-tests were conducted to further investigate whether these two ERP learning effects were location-specific and whether they were modified in the tests.
3.2 Results 3.2.1 Behavioral resultsFor all subjects, accuracies in the training and test blocks were calculated to reveal the fast PL effect and its location specificity. As shown in Figure 5A, ~60 min of training significantly improved behavioral performance [main effect of block, F (3, 45) = 11.258, p < 0.001, ηp2 = 0.429], as reflected by the higher accuracy in b4 (b1 vs. b4 improvement, 0.077 ± 0.017, mean ± SE, p = 0.0004; 95% CI = [0.04, 0.11]). Consistent with previous behavioral studies showing location specificity of PL, the performance improvement disappeared at the untrained location when subjects were required to discriminate the Vernier offset at this location in the tests [pre vs. post, 0.027 ± 0.028, t(15) = 0.989, p = 0.338, d = 0.247; Figure 5A]. Accuracy was significantly lower in the posttest than in b4 (p = 0.0003), further confirming the location specificity of PL.
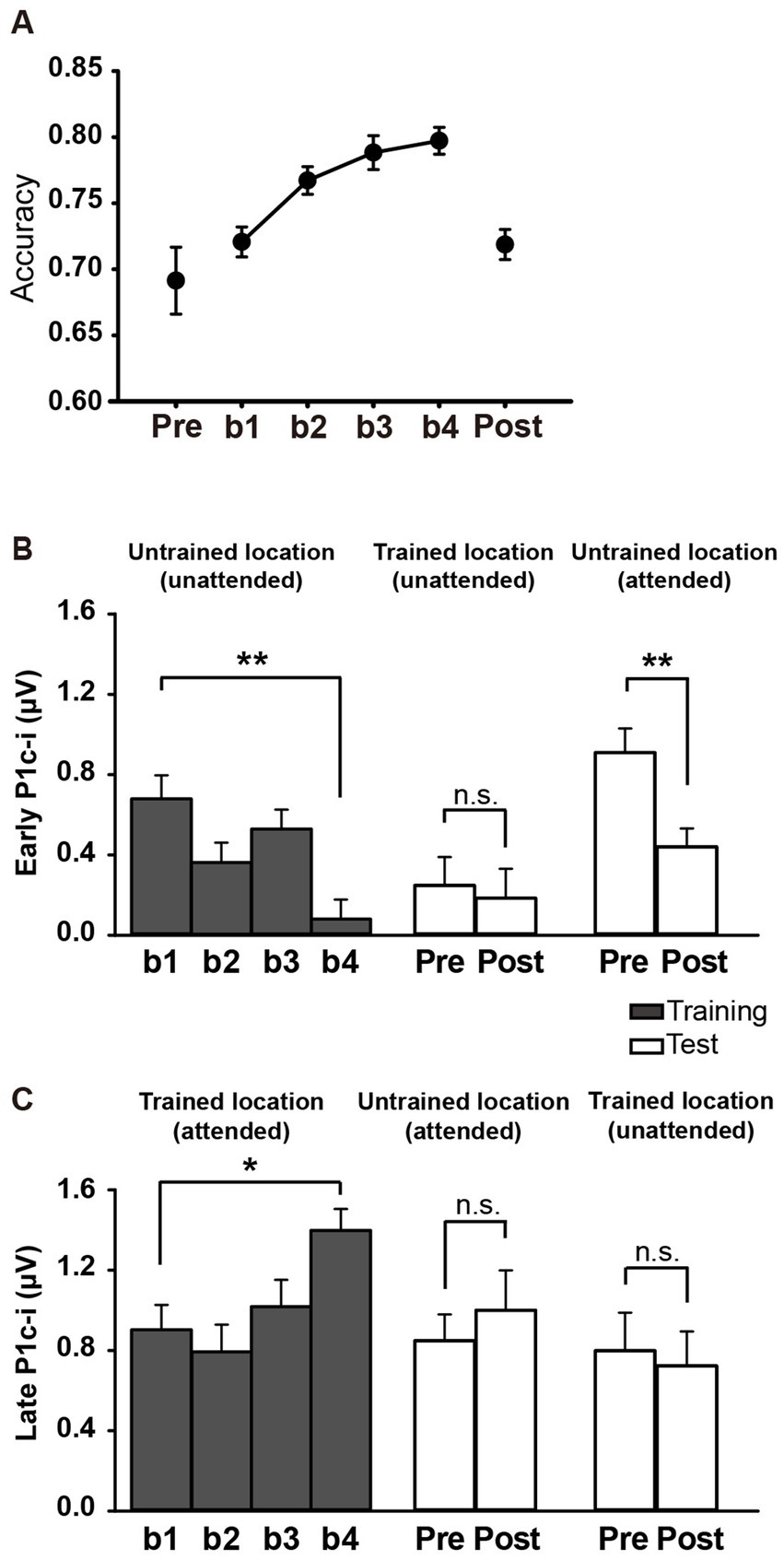
Figure 5. Behavioral and ERP results in Experiment 2. (A) Mean accuracies during training and test blocks. Behavioral performance improved significantly with training, as reflected by higher Vernier discrimination accuracy in b4, and this learning effect was specific to the trained location. (B,C) Mean amplitudes of the early and late P1c-i in training and test blocks. P1c-i waves were measured at electrodes (collapsed across PO7/8, PO3/4, P7/8, and P5/6) in two distinct time intervals (i.e., 75–95 ms and 100–120 ms). Training decreased the early P1c-i (75–95 ms) at the untrained location, as indexed by a smaller early P1c-i in b4 than in b1, and this effect was preserved when the untrained location became attended in the tests. The early P1c-i effect was not observed at the trained location when it was the distractor/unattended location in the tests. In contrast, the late P1c-i (100–120 ms) at the trained location increased between b1 and b4. This effect disappeared when the trained location became unattended in the tests. The late P1c-i effect was not observed at the untrained location when it was the target/attended location in the tests. * p < 0.05, ** p < 0.01, and n.s. indicates p > 0.5. Error bars indicate within-subject standard errors.
3.2.2 ERP resultsThe P1c-i waves of targets at the trained location and those of distractors at the untrained location during the training were shown in Figures 6A,B, respectively. The P1c-i waves of distractors at the trained location and those of targets at the untrained location in the tests were illustrated in Figures 6C,D, respectively.
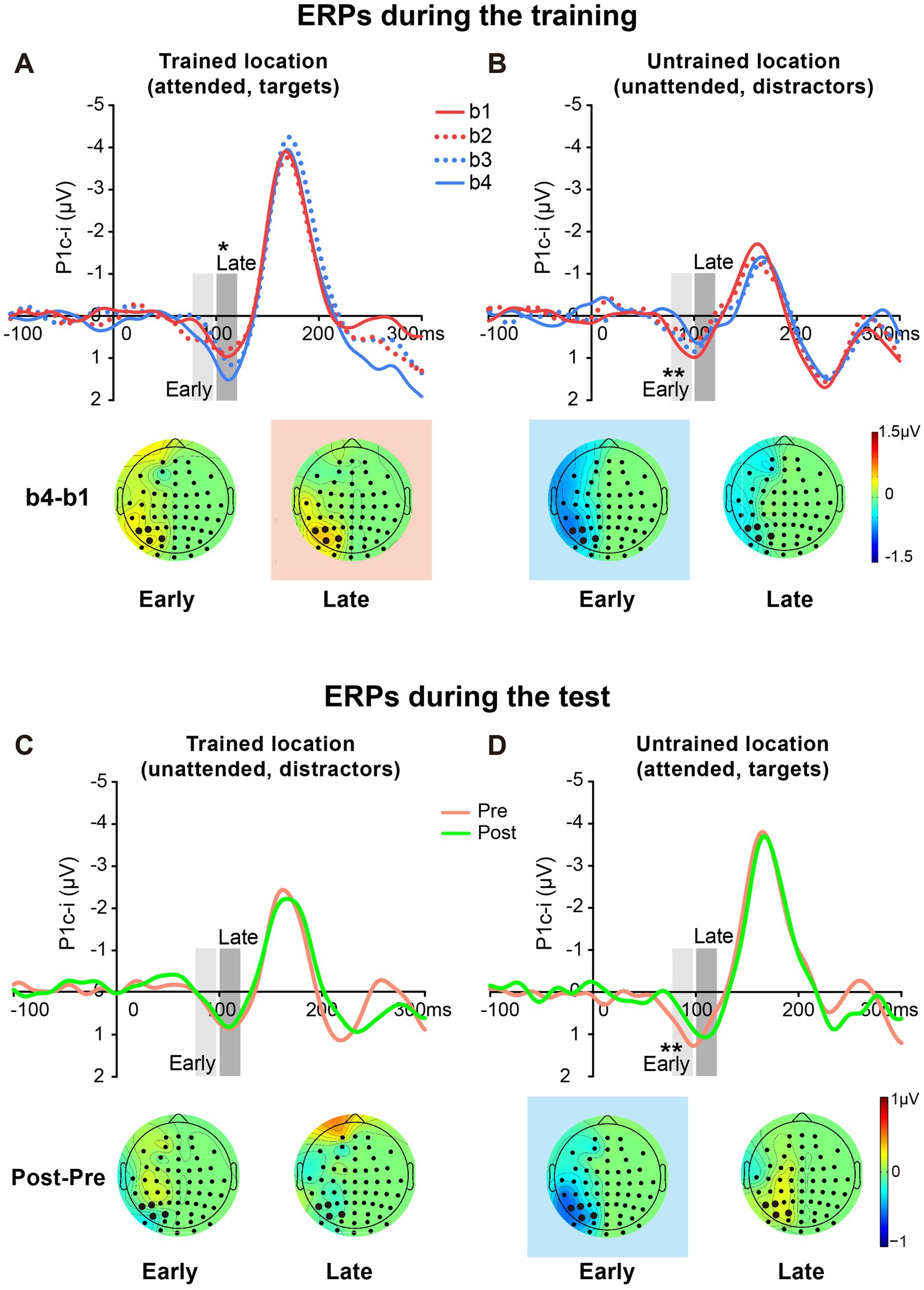
Figure 6. Grand average ERPs and voltage topographies in training and test blocks of Experiment 2. (A,B) Contralateral-minus-ipsilateral difference ERP waveforms elicited by targets at the trained location (A) and by distractors at the untrained location (B) during the training. P1c-i waves were measured in two different time intervals (i.e., 75–95 ms and 100–120 ms), shown as shaded rectangles. Topographies of these intervals were plotted based on ERP difference waves constructed by subtracting b1 from b4. These voltage distributions of difference waves were projected onto the left hemisphere. Similar to Experiment 1, training increased late P1c-i (100–120 ms) at the trained but not the untrained location, and decreased early P1c-i (75–95 ms) at the untrained but not the trained location. (C,D) Contralateral-minus-ipsilateral difference ERP waveform elicited by distractors at the trained location (C) and by targets at the untrained location (D) in the tests. Topographies of two intervals were plotted based on ERP difference waves between pretest and posttest. The electrodes highlighted in bold represent an electrode group (PO7/8, PO3/4, P7/8, and P5/6) used for the analysis of learning effects. Red in the topographic plot indicates training-induced larger P1c-i, and blue indicates training-induced smaller P1c-i. * p < 0.05, ** p < 0.01.
3.2.2.1 Early P1c-iSimilar to Experiment 1, a significant interaction of location (Trained vs. Untrained) × training block (b1, b2, b3, and b4) was found for the early P1c-i [75–95 ms; F (3, 45) = 7.932, p = 0.002, ηp2 = 0.346]. Again, training significantly decreased the early P1c-i amplitude at the untrained location (F (3, 45) = 5.673, p = 0.002, ηp2 = 0.274; the decrement occurred across the four training blocks rather than after a single block: b1 vs. b4, −0.599 ± 0.171 μV, mean ± SE, p = 0.003, 95% CI = [−0.96, −0.24]; b1 vs. b2, −0.316 ± 0.171 μV, p = 0.084, 95% CI = [−0.68, 0.05]; see Figures 5B, 6B), but not at the trained location (b1 vs. b4, 0.316 ± 0.227 μV, p = 0.185, 95% CI = [−0.17, 0.80]; Figure 6A).
The early P1c-i decrement was not observed at the trained location in the tests [pre vs. post, −0.063 ± 0.222 μV, t(15) = −0.286, p = 0.779, d = −0.071; Figures 5B, 6C], reinforcing that this learning-induced suppression effect on the early P1c-i is specific to the untrained location. In addition, the early P1c-i effect observed at the untrained location during the training (Figure 6B) was preserved when the untrained location became attended in the test [pre vs. post, −0.469 ± 0.134 μV, t(15) = −3.499, p = 0.003, d = −0.875; Figures 5B, 6D]; that is, the early P1c-i effect appeared regardless of whether the untrained location was attended or not, indicating that this suppression effect reflects modifications in automatic/involuntary processing at the untrained location.
3.2.2.2 Late P1c-iConsistent with Experiment 1, training significantly increased the late P1c-i amplitude at the trained location (F (3, 45) = 4.243, p = 0.010, ηp2 = 0.220; the increment occurred across the four training blocks rather than after a single block: b1 vs. b4, 0.495 ± 0.169 μV, mean ± SE, p = 0.010, 95% CI = [0.14, 0.86]; b1 vs. b2, −0.110 ± 0.204 μV, p = 0.597, 95% CI = [−0.55, 0.33]); Figures 5C, 6A), but not at the untrained location [F (3, 45) = 1.062, p = 0.357, ηp2 = 0.066; Figure 6B].
The late P1c-i increment was not observed at the untrained location in the tests [pre vs. post, 0.151 ± 0.253 μV, t(15) = 0.599, p = 0.558, d = 0.150; Figures 5C, 6D], reinforcing that the learning-induced facilitation effect on late P1c-i is specific to the trained location. Different from the early P1c-i effect, the late P1c-i learning effect observed at the trained location during the training (Figure 6A) disappeared when the trained location became unattended in the test [pre vs. post, −0.075 ± 0.274 μV, t(15) = −0.275, p = 0.787, d = −0.069; Figures 5C, 6C], indicating this facilitation effect reflects changes of voluntary allocation of attention to the trained location.
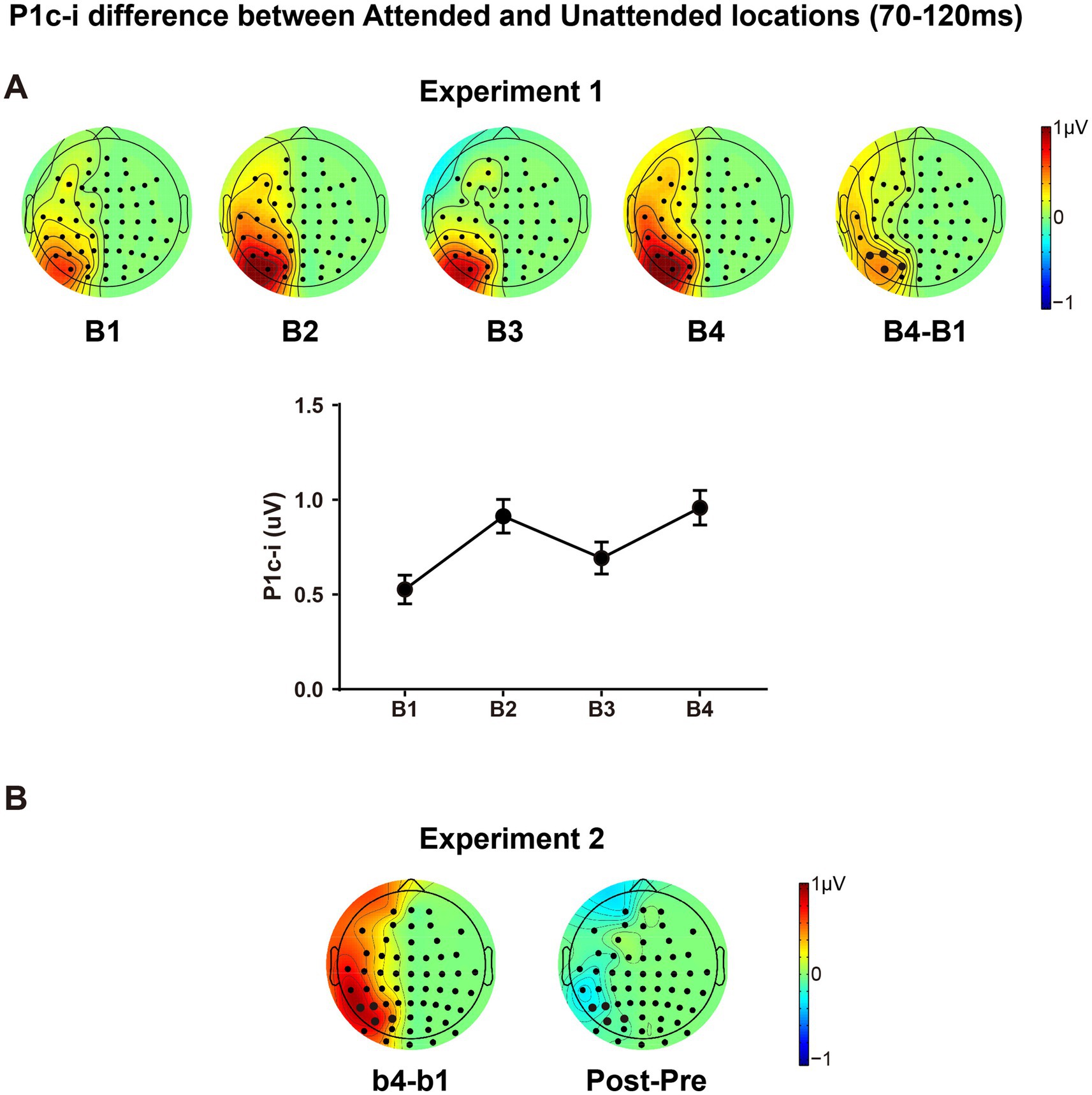
3.2.2.3 PL-induced changes of spatial attention effectThe present design also allowed us to investigate whether and how spatial attentional effect on early ERPs is modulated by fast learning. As shown in Figure 7, there is a clear attentional effect in P1c-i (70–120 ms) for each training block, as reflected by topographical voltage maps of the P1c-i difference between the attended and unattended locations. This spatial attention effect on P1c-i was enhanced significantly through tens of minutes of training in both Experiment 1 [B1 vs. B4 increment, 0.432 ± 0.131 μV, t(15) = 3.290, p = 0.005, d = 0.823] and Experiment 2 [b1 vs. b4 increment, 0.769 ± 0.250 μV, t(15) = 3.081, p = 0.008, d = 0.770]. However, this fast learning effect on visuospatial attention disappeared when the attended and unattended locations switched in the tests [Pre vs. Post, −0.136 ± 0.185 μV, t(15) = −0.734, p = 0.474, d = −0.184], which is consistent with the location specificity of fast PL.
留言 (0)