
記住我
Brucellosis, a severe zoonotic infectious disease caused by bacteria of the genus Brucella, poses a significant global public health threat due to its widespread prevalence (Pappas et al., 2006). This disease not only hampers the development of the livestock industry but also spreads to humans through direct contact with infected animals, ingestion of inadequately processed dairy products, or inhalation of aerosols containing the bacteria (Fuchs et al., 2016; Abdel-Hamid et al., 2021). Clinical symptoms in humans include fever, fatigue, and joint pain, and in severe cases, it can lead to complications such as endocarditis, arthritis, and even death (Pappas et al., 2005; Franco et al., 2007). Therefore, the development of efficient and accurate diagnostic methods for brucellosis is crucial for its prevention, control, and treatment.
Currently, the diagnosis of brucellosis mainly relies on various methods, including pathogen detection, serological testing, and molecular biology techniques. Among these, serological testing is widely used due to its simplicity, low cost, and ability to reflect the immune status of the patient to some extent (Araj, 2010; Yagupsky et al., 2019). However, most existing serological diagnostic antigens are based on the cell wall components or outer membrane proteins of Brucella. Although these antigens possess certain immunogenicity, their diagnostic sensitivity and specificity are often compromised in practical applications due to factors such as the patient’s immune status, the stage of infection, and cross-reactivity (Chart et al., 1992; Bonfini et al., 2018).
VirB proteins, as key components of the Type IV secretion system (T4SS) in Brucella, play an important role in the interaction between the bacteria and host cells (Boschiroli et al., 2002; Zheng et al., 2024). The VirB system is not only involved in the survival and replication of Brucella within host cells but is also closely related to its pathogenic mechanisms. Despite the crucial role of VirB proteins in the biological functions of Brucella, there has been little research systematically analyzing their value in the serological diagnosis of human brucellosis. This gap in research limits our understanding of the pathogenic mechanisms of brucellosis and hinders the development and application of novel diagnostic antigens.
In this study, we aim to utilize Tandem Mass Tag (TMT) proteomics technology to prepare and evaluate the value of Brucella VirB proteins in the serological diagnosis of human brucellosis. TMT technology, known for its high sensitivity and high throughput in protein quantification, allowed for precise identification and quantification of proteins in complex biological samples, providing strong technical support for the screening and validation of novel diagnostic antigens (Moulder et al., 2018; Deng et al., 2022). Through TMT proteomics, we identified highly expressed VirB proteins in wild-type Brucella strains, prepared recombinant Type IV secretion system proteins, and established a serological detection method based on these proteins. The goal was to enhance the sensitivity and specificity of brucellosis diagnosis, thereby offering new antigen choices for the early diagnosis and prevention of the disease.
Materials and methodsSerum samples and bacterial strainsIn this study, a total of 100 positive and 96 negative serum samples were obtained from the Xuzhou Center for Disease Control and Prevention, all confirmed as positive or negative through tube agglutination tests. Additionally, serum from 40 febrile patients infected with other pathogens (stored in the laboratory, with detailed information available in Supplementary File 1: Cross-Reactivity Assessment) was used to evaluate the cross-reactivity of the developed method. To identify highly expressed proteins in wild strain of Brucella abortus, as well as to discover antigenic proteins that can be utilized in the diagnosis of human brucellosis, the vaccine strain Brucella abortus A19 and the wild-type Brucella abortus DT21, both isolated and preserved by the China Animal Health and Epidemiology Center, were also utilized in this study.
Proteomics analysisBacterial cultureThe preserved bacterial strain was inoculated into 500 mL of Tryptic Soy Broth (TSB, STBMTSB12, Millipore, USA) medium and incubated at 37°C with shaking for 24-48 hours. After incubation, 5 mL of 1% formaldehyde was added to inactivate the bacteria, which was then stored at 4°C for later use.
Proteomics analysisProteomics analysis was performed according to standard protocols referenced from the literature, including steps such as protein extraction and quantification, protein digestion and TMT labeling, LC-MS/MS analyses, and qualitative and quantitative analysis of proteins (Deng et al., 2022). The peak intensities of TMT-tagged reagent ions were quantitatively compared across samples (Supplementary Table S1). The statistical analysis of the differential proteins identified was conducted using analysis of variance (ANOVA), with a significance threshold set at p < 0.05. Proteins exhibiting a fold change greater than 1.2 (ratio≥1.2) or less than 0.83(ratio ≤ 0.83) were classified as highly expressed proteins.
Preparation of recombinant T4SS proteinsBased on TMT proteomics analysis, we selected highly expressed T4SS proteins from the wild-type strain. The amino acid sequences of these proteins were retrieved from the NCBI protein database. Using the UniProt website (https://www.uniprot.org/uniprotkb), we predicted and removed transmembrane regions, signal peptides, and hydrophobic regions, constructing the recombinant sequences of the Type IV secretion system proteins. The sequence was then submitted to Beijing Protein Innovation Co., LTD. for codon optimization to suit prokaryotic expression. Gene synthesis was carried out, and a 6xHis tag was added to facilitate subsequent protein purification.
The synthesized recombinant protein gene was cloned into the pET30a expression vector. The vector was then transformed into competent BL21 cells for IPTG-induced expression. The procedure was conducted as follows: Competent BL21 cells, previously stored at -80°C, were thawed on ice and subsequently mixed with pET30a(+). The mixture was incubated on ice for 30 minutes, followed by a heat shock at 42°C for 90 seconds, after which the cells were immediately cooled on ice for 2 minutes. Subsequently, 800 μL of LB medium (L113084, Aladdin, USA) was added, and the cells were incubated at 37°C for 45 minutes. The culture was then centrifuged at 3214×g (Eppendorf Centrifuge 5810R, Germany) for 3 minutes, with most of the supernatant discarded, leaving approximately 100-150 μL, in which the cells were resuspended. The resuspended cells were plated onto LB agar plates containing the appropriate antibiotic and incubated overnight at 37°C. The following day, the cultured bacterial solution was transferred into 250 mL of LB liquid medium supplemented with the corresponding antibiotic and incubated at 37°C with shaking at 200 rpm using DHZ-DA Large-capacity full-temperature oscillator (Changzhou Guoyu Instrument Manufacturing Co., China) until the optical density at 600 nm (OD600) reached 0.6-0.8. Induction of protein expression was achieved by adding 0.5 mM IPTG (16758, Sigma, Germany) and continuing the incubation at 37°C for 4 hours. The culture was then centrifuged at 8228×g for 6 minutes, the supernatant was discarded, and the cell pellet was collected. The pellet was resuspended in 20-30 mL of 10 mM Tris-HCl (pH 8.0) solution and subjected to ultrasonic disruption (500 W, 180 cycles, 5 seconds per cycle with 5-second intervals). A 100 μL aliquot of the disrupted bacterial suspension was centrifuged at 18514×g for 10 minutes. Of the resulting supernatant, 50 μL was transferred to a separate Eppendorf tube, while the pellet was resuspended in 50 μL of 10 mM Tris-HCl (pH 8.0) solution. To ascertain the presence of the target protein in either the supernatant or the pellet, 12% SDS-PAGE (P0012AC, Beyotime Biotechnology, Shanghai, China) electrophoresis was performed for subsequent purification.
The nickel column (Ni Sepharose 6 Fast Flow, GE Healthcare) was washed with deionized water until the pH reached 7.0, then equilibrated with approximately 100 mL of 10 mM Tris-HCl (pH 8.0, T3253, Sigma, Germany) solution. The column was further equilibrated with approximately 50 mL of 10 mM Tris-HCl (pH 8.0) solution containing 0.5 M NaCl (A501218-0001, Sangon Biotch, Shanghai, China). The sample containing the target protein was diluted and loaded onto the column. After loading, the column was washed with 10 mM Tris-HCl (pH 8.0) solution containing 0.5 M NaCl. The protein was eluted using 10 mM Tris-HCl (pH 8.0) solutions containing 15 mM, 60 mM, and 300 mM imidazole (with 0.5 M NaCl). The protein peaks were collected, and purification efficiency was analyzed by 12% SDS-PAGE electrophoresis. The protein was quantified using the BCA Protein Quantification Kit (P0010, Beyotime).
Establishment of indirect ELISA method and serum detectionThe indirect enzyme linked immunosorbent assay (iELISA) method was established as follows: The purified protein was first diluted in carbonate buffer solution (CBS, pH=9.6) to a concentration of 10 µg/mL, and 100 µL per well was added to a 96-well microplate (Corning, USA). The plate was incubated overnight at 4°C. After washing three times with PBST, 300 µL of blocking solution (5% skim milk in PBS) was added to each well and incubated at 37°C for 2 hours. The plate was washed again with PBST, then human serum diluted in PBS (1:200) was added and incubated at 37°C for 1 hour. After three more washes with PBST, 100 µL of HRP-conjugated rabbit anti-human IgG (diluted 1:10,000, A18903, Thermo Fisher, USA) was added to each well and incubated at 37°C for 1 hour. The plate was washed three times with PBST, tetramethylbenzidine (TMB, T2573, TCI, Japan) substrate solution was added, and the plate was incubated in the dark for 10 minutes for color development. The reaction was stopped with 2M H2SO4, and the OD450 was measured using a microplate reader (Versa Max microplate reader, MD, USA). Laboratory-stored lipopolysaccharide (LPS, provided by the China Animal Health and Epidemiology Center, 3 mg/mL) and Rose Bengal Ag (diluted 1:400, IDEXX Pourquier, Montpellier, France) were used as control antigens, and serum samples were tested in triplicate using the same procedure. Sensitivity, specificity, area under the curve (AUC), and the cut-off value were determined by receiver operating characteristic curve (ROC) analysis.
Evaluation of cross-reactivity in indirect ELISA methodFollowing the procedure described above, sera from febrile patients without brucellosis were tested using the constructed Brucella T4SS recombinant proteins to evaluate the cross-reactivity by comparing with LPS and Rose Bengal Ag. Cross-reactivity was assessed based on the cut-off value determined by the ROC curve.
Statistical methodsDot plot and ROC curve analyses were conducted using GraphPad Prism version 6.05. Statistical analyses were performed using unpaired Student’s t-test and ANOVA, with a significance level set at P < 0.05.
ResultsSelection of recombinant type IV secretion system proteinsThrough TMT quantitative analysis, a total of 152 highly expressed proteins were identified in the wild-type strain, and 102 highly expressed proteins were identified in the vaccine strain (Supplementary File 2). Among the highly expressed proteins of the wild strain, we identified seven T4SS proteins, including six VirB proteins (VirB3, VirB4, VirB8, VirB9, VirB10, VirB11) and one T4SS putative outer membrane lipoprotein (BMEII0036). After predicting and removing transmembrane regions, signal peptides, and hydrophobic regions through the UniProt website (https://www.uniprot.org/uniprotkb), we constructed recombinant sequences of the seven proteins for prokaryotic expression (Table 1).
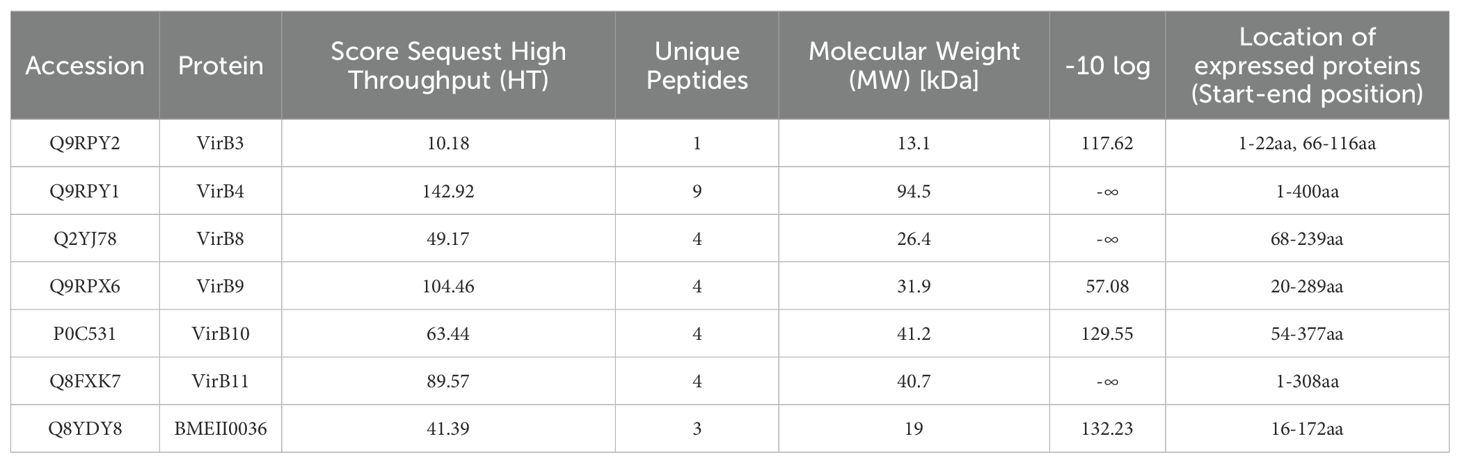
Table 1. Proteomics-based analysis and screening of Type IV secretion system proteins.
Preparation of recombinant T4SS proteinsAll seven recombinant proteins were successfully expressed and purified through prokaryotic expression, as shown in Figure 1 and Supplementary File 3. After quantification by BCA assay, the concentration was adjusted to 0.5 mg/mL in PBS and stored at -20°C for future use.
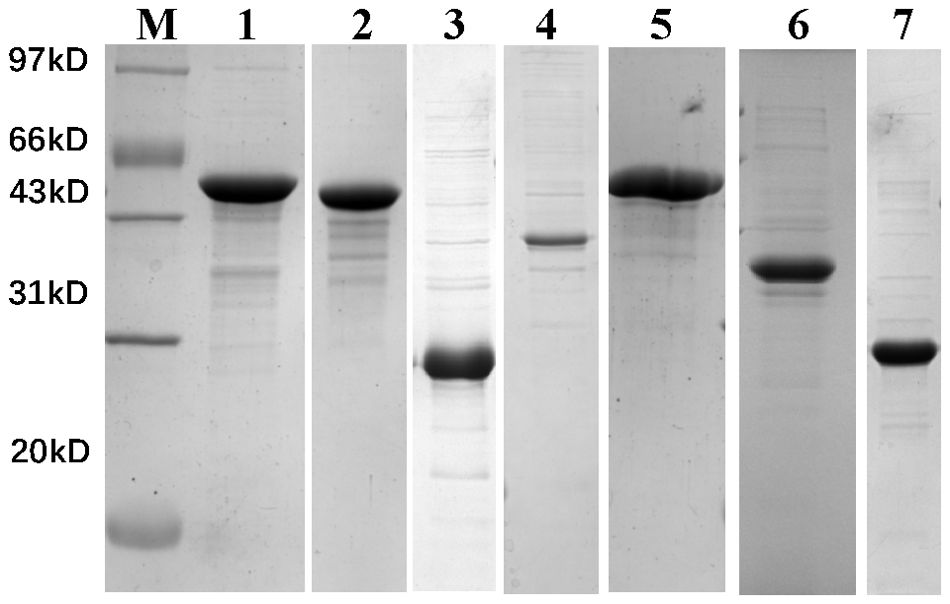
Figure 1. 12% SDS-PAGE analysis of recombinant protein prokaryotic expression results. M, marker; Lane 1, VirB4; Lane 2, VirB11; Lane 3, VirB8; Lane 4, VirB3; Lane 5, VirB10; Lane 6, VirB9; Lane 7, BMEII0036.
Results of iELISAAccording to the ROC curve analysis, the diagnostic accuracy of the recombinant proteins, ranked from highest to lowest, is as follows: rVirB3, rVirB4, rVirB9, rBMEII0036, rVirB8, rVirB11, and rVirB10. The area under the ROC curve (AUC) for each protein is 0.9979, 0.9914, 0.9825, 0.9817, 0.9782, 0.9764, and 0.9476, respectively, which is slightly lower compared to LPS and Rose Bengal Ag. According to the Youden index calculation, the sensitivity of these proteins is all above 0.9100, and the specificity is all above 0.9167. The highest sensitivity is 0.9900 (95% CI, 0.9455 - 0.9997) for rVirB4 and rVirB9, and the highest specificity is 0.9896 (95% CI, 0.9433 - 0.9997) for rVirB3. The sensitivity and specificity are slightly lower than those of LPS and Rose Bengal Ag. The results are presented in Table 2, Figure 2, and Supplementary File 1.
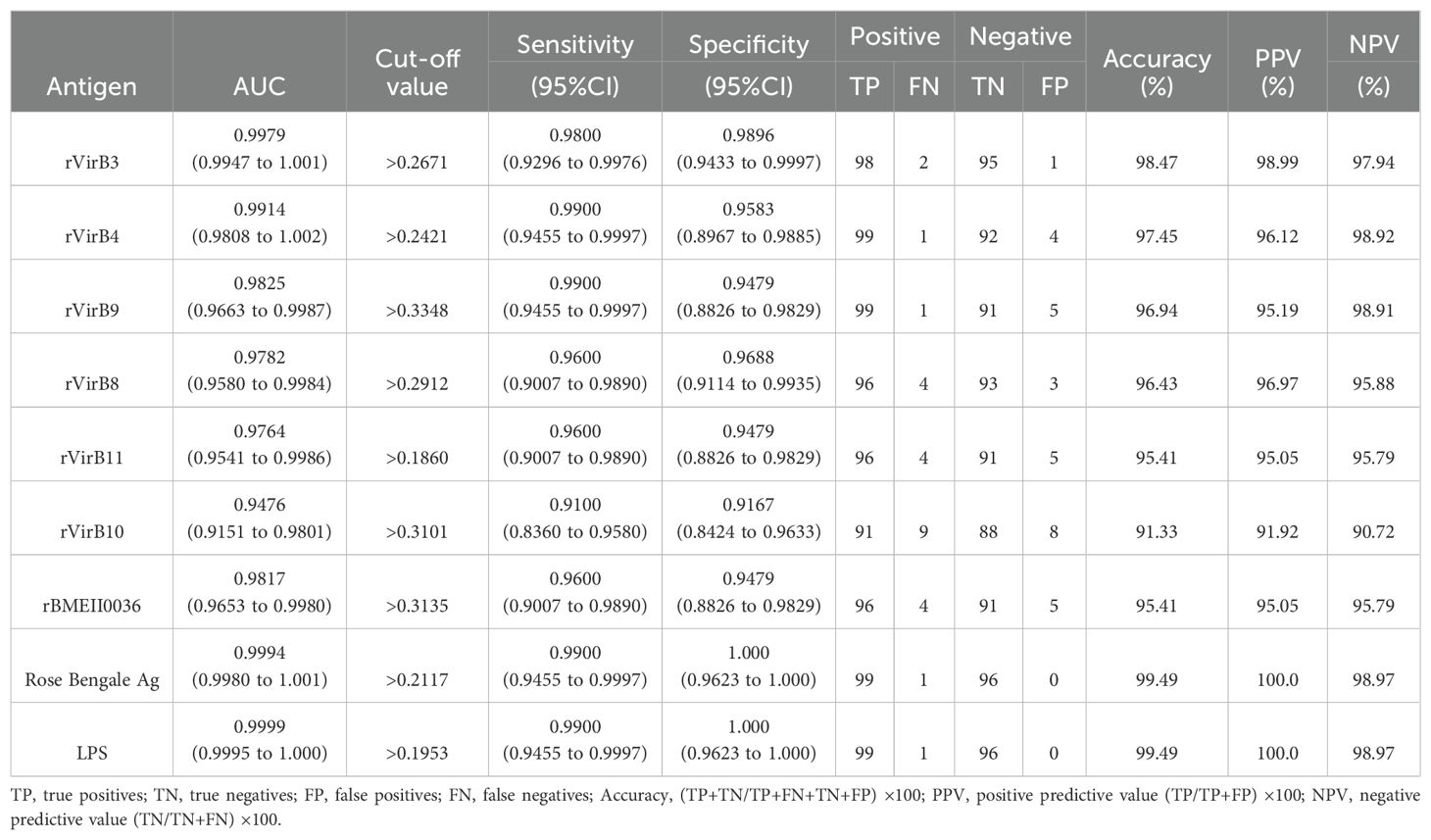
Table 2. Evaluation of ELISA results of the recombinant proteins.
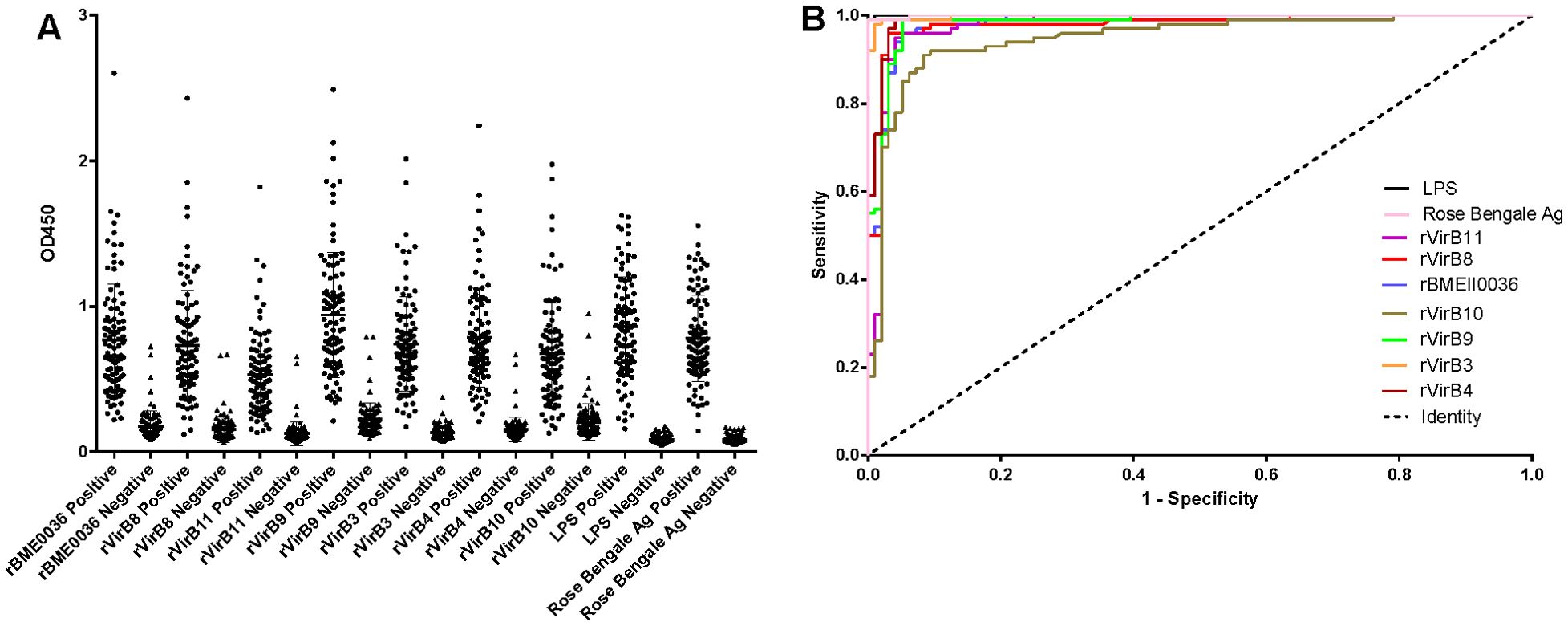
Figure 2. I-ELISA analysis of human serum samples. (A) Dotplot of the human sera. (B) ROC analysis of human sera.
Cross-reactivity assessmentUsing iELISA and based on the determined cut-off values, cross-reactivity was observed in 2, 5, 8, 2, 1, 5, and 0 out of 40 serum samples from clinical febrile patients without brucellosis when tested with rVirB3, rVirB4, rVirB9, rBMEII0036, rVirB8, rVirB11, and rVirB10, respectively. In contrast, cross-reactivity with LPS and Rose Bengale Ag was observed in 16 and 18 cases, respectively. The cross-reactive pathogens with LPS and Rose Bengale Ag were primarily concentrated in Escherichia coli. Specifically, cross-reactivity with LPS included 8 cases of Escherichia coli infection, 3 cases of Staphylococcus aureus, and 1 case each of Enterococcus faecium, Klebsiella pneumoniae, Moraxella osloensis, Pseudomonas putida, and Streptococcus dysgalactiae. Cross-reactivity with Rose Bengale Ag was observed in 18 cases, including 7 cases of Escherichia coli infection, 2 cases each of Enterococcus faecium, Klebsiella pneumoniae, and Staphylococcus aureus, and 1 case each of Aeromonas sobria, Moraxella osloensis, Pseudomonas aeruginosa, Pseudomonas putida, and Rothia mucilaginosa. The results are shown in Supplementary File 1.
DiscussionThe T4SS is a crucial virulence factor of Brucella, composed of 12 protein complexes named VirB1 to VirB12, encoded by the VirB regions (Ke et al., 2015). Numerous studies have explored the potential of VirB proteins for vaccine development and serological diagnosis. For instance, Zai et al (2021) evaluated the use of VirB8 against pathogenic Brucella species through composite reverse vaccinology. They found that VirB8 could induce specific humoral and cellular immune responses, reduce the bacterial load of B. abortus S19 in mice, and provide varying degrees of protection. Yin et al (2023) used immunoinformatics to identify antigenic epitopes of VirB8 and VirB10 from the Brucella T4SS, screening two cytotoxic T lymphocyte epitopes, nine helper T lymphocyte epitopes, six linear B cell epitopes, and six conformational B cell epitopes for constructing a multi-epitope vaccine. Several studies have also confirmed that combining VirB10 with other proteins to create recombinant vaccines can successfully induce immune responses (Tarrahimofrad et al., 2022; Rahimnahal et al., 2023). Research has shown that VirB7 and VirB9 can induce Th1 responses in mice and dogs (Pollak et al., 2015). Additionally, there is evidence supporting the potential value of VirB5, VirB10, and VirB12 for serological diagnosis of brucellosis (Tan et al., 2012; Mirkalantari et al., 2017; Pathak et al., 2018). However, existing studies have mainly focused on individual VirB proteins, and there is a lack of systematic analysis on the use of VirB proteins for serological diagnosis of brucellosis.
In this study, we used TMT proteomics technology to identify highly expressed VirB proteins from wild-type Brucella strains and successfully prepared various recombinant VirB proteins for serological diagnosis. The results demonstrated that several VirB proteins (e.g., rVirB3, rVirB4, rVirB9) exhibited high sensitivity and specificity in diagnosing brucellosis. Although their performance was slightly lower than that of traditional LPS and Rose Bengale Ag, their potential as novel diagnostic antigens cannot be overlooked, the proteins still show good results for the diagnosis of human brucellosis. Besides VirB proteins, we also identified a T4SS-related protein through proteomics, namely T4SS putative outer membrane lipoprotein BMEII0036, which also showed high sensitivity and specificity when used in brucellosis diagnosis. As key components of the Brucella T4SS, VirB proteins not only play a role in the pathogen’s virulence mechanisms but also in its interaction with host cells, making them valuable diagnostic antigens that can more directly reflect the infection status of Brucella, with significant clinical application potential (Xiong et al., 2021).
In this study, we observed some differences in diagnostic performance among the various VirB proteins. For example, rVirB3 showed the best specificity, while rVirB4 and rVirB9 had the highest sensitivity. These differences might be attributed to the specific roles and expression levels of different VirB proteins during the Brucella lifecycle. Additionally, the antigenicity of these proteins could be influenced by factors such as amino acid sequences, spatial conformation, and glycosylation modifications (Pavlenko et al., 1990; Rahman et al., 2016; Shi et al., 2024). Therefore, future research should further explore the antigenic epitopes of these proteins and how to optimize their structures to enhance diagnostic performance.
Cross-reactivity is an important factor in assessing the specificity of diagnostic antigens. This study found that although all VirB proteins exhibited some degree of cross-reactivity, the frequency and intensity of cross-reactivity were lower than those of LPS and Rose Bengale Ag. Notably, rVirB10 showed no cross-reactivity in 40 serum samples from febrile patients without brucellosis, indicating very high specificity. This suggests that VirB proteins may have an advantage in reducing cross-reactivity when used as diagnostic antigens. However, it is important to note that cross-reactivity should be assessed with a broader and more diverse sample set to comprehensively evaluate their specificity in practical applications.
Although the TMT proteomics results indicate that other VirB proteins, including VirB1, VirB2, and VirB5-VirB7, did not exhibit any significant differences between vaccine strain Brucella abortus A19 and the wild-type Brucella abortus DT21, it is essential to further investigate the potential diagnostic value of these proteins for human brucellosis in future studies. This study provides compelling evidence that T4SS proteins play a crucial role in human brucellosis infection and offers important experimental evidence and theoretical foundation for the development of new diagnostic antigens for brucellosis. However, further research and exploration are necessary to achieve widespread clinical application of VirB proteins. In summary, VirB proteins, as key components of the Brucella T4SS, show great potential in the serological diagnosis of brucellosis. Future research should continue to explore their antigenicity and diagnostic performance to develop more efficient and accurate diagnostic methods for brucellosis.
Data availability statementThe original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statementThe studies involving humans were approved by Ethics Committee of Xuzhou Medical University. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributionsQW: Methodology, Writing – original draft. CS: Methodology, Writing – review & editing. LG: Writing – review & editing. YX: Methodology, Writing – review & editing. JZ: Writing – review & editing. DY: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing.
FundingThe author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by Xuzhou Science and Technology Bureau (Grant number KC23306), the Medical Research Program of Jiangsu Commission of Health (Grant number Z2023080), Postgraduate Research & Practice Innovation Program of Jiangsu Province (Grant number KYCX23-2963), and QingLan Project of Jiangsu Province (2024). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
AcknowledgmentsWe thank the China Animal Health and Epidemiology Center for the gift of LPS and Brucella strains, and the Xuzhou Center for Disease Control and Prevention for the gift of human brucellosis sera (positive and negative).
Conflict of interestThe authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statementThe author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary materialThe Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2024.1514046/full#supplementary-material
Supplementary File 1 | Raw data for iELISA and information of febrile patients infected with other pathogens.
Supplementary File 2 | Information on highly expressed proteins of wild strains and vaccine strains.
Supplementary File 3 | Original SDS-PAGE image of recombinant proteins.
AbbreviationsT4SS, Type IV secretion system; TMT, Tandem Mass Tag; TSB, Tryptic Soy Broth; OD, optical density; AUC, area under the ROC curve; ROC, receiver operating characteristic curve; iELISA, indirect enzyme linked immunosorbent assay; CBS, carbonate buffer solution; TMB, tetramethylbenzidine; LPS, lipopolysaccharide; ANOVA, analysis of variance; HT, High Throughput; MW, Molecular Weight.
ReferencesAbdel-Hamid, N. H., Ghobashy, H. M., Beleta, E. I., Elbauomy, E. M., Ismail, R. I., Nagati, S. F., et al. (2021). Risk factors and Molecular genotyping of Brucella melitensis strains recovered from humans and their owned cattle in Upper Egypt. One Health (Amsterdam Netherlands). 13, 100281. doi: 10.1016/j.onehlt.2021.100281
PubMed Abstract | Crossref Full Text | Google Scholar
Bonfini, B., Chiarenza, G., Paci, V., Sacchini, F., Salini, R., Vesco, G., et al. (2018). Cross-reactivity in serological tests for brucellosis: a comparison of immune response of Escherichia coli O157:H7 and Yersinia enterocolitica O:9 vs Brucella spp. Vet. Ital. 54, 107–114. doi: 10.12834/VetIt.1176.6539.2
PubMed Abstract | Crossref Full Text | Google Scholar
Boschiroli, M. L., Ouahrani-Bettache, S., Foulongne, V., Michaux-Charachon, S., Bourg, G., Allardet-Servent, A., et al. (2002). Type IV secretion and Brucella virulence. Veterinary Microbiol. 90, 341–348. doi: 10.1016/S0378-1135(02)00219-5
PubMed Abstract | Crossref Full Text | Google Scholar
Chart, H., Okubadejo, O. A., Rowe, B. (1992). The serological relationship between Escherichia coli O157 and Yersinia enterocolitica O9 using sera from patients with brucellosis. Epidemiol. Infect. 108, 77–85. doi: 10.1017/S0950268800049529
PubMed Abstract | Crossref Full Text | Google Scholar
Deng, H., Xue, B., Wang, M., Tong, Y., Tan, C., Wan, M., et al. (2022). TMT-Based Quantitative Proteomics Analyses Reveal the Antibacterial Mechanisms of Anthocyanins from Aronia melanocarpa against Escherichia coli O157:H7. J. Agric. Food Chem. 70, 8032–8042. doi: 10.1021/acs.jafc.2c02742
PubMed Abstract | Crossref Full Text | Google Scholar
Fuchs, I., Osyntsov, L., Refaely, Y., Ciobotaro, P., Zimhony, O. (2016). Ritual slaughter as overlooked risk factor for brucellosis. Emerging Infect. diseases. 22, 746–748. doi: 10.3201/eid2204.151192
PubMed Abstract | Crossref Full Text | Google Scholar
Mirkalantari, S., Zarnani, A. H., Nazari, M., Irajian, G. R., Amirmozafari, N. (2017). Brucella melitensis VirB12 recombinant protein is a potential marker for serodiagnosis of human brucellosis. Ann. Clin. Microbiol. antimicrobials. 16, 8. doi: 10.1186/s12941-017-0182-4
PubMed Abstract | Crossref Full Text | Google Scholar
Moulder, R., Bhosale, S. D., Goodlett, D. R., Lahesmaa, R. (2018). Analysis of the plasma proteome using iTRAQ and TMT-based Isobaric labeling. Mass Spectrom Rev. 37, 583–606. doi: 10.1002/mas.21550
PubMed Abstract | Crossref Full Text | Google Scholar
Pappas, G., Papadimitriou, P., Akritidis, N., Christou, L., Tsianos, E. V. (2006). The new global map of human brucellosis. Lancet Infect. Dis. 6, 91–99. doi: 10.1016/S1473-3099(06)70382-6
PubMed Abstract | Crossref Full Text | Google Scholar
Pathak, P., Kumar, A., Sarangi, P. P., Bhagyawant, S., Thavaselvam, D. (2018). Cloning, expression and purification of virB10 protein of Brucella melitensis and evaluation of its role as a serological marker for Brucella infection in experimental and natural host. Protein Expression purification. 145, 53–58. doi: 10.1016/j.pep.2017.12.014
PubMed Abstract | Crossref Full Text | Google Scholar
Pavlenko, A. F., Chikalovets, I. V., Kurika, A. V., Glasunov, V. P., Mikhalyuk, L. V., Ovodov, Y. (1990). Carcinoembryonic antigen, its spatial structure and localization of antigenic determinants. Tumour Biol. 11, 306–318. doi: 10.1159/000217666
PubMed Abstract | Crossref Full Text | Google Scholar
Pollak, C. N., Wanke, M. M., Estein, S. M., Delpino, M. V., Monachesi, N. E., Comercio, E. A., et al. (2015). Immunization with Brucella VirB proteins reduces organ colonization in mice through a Th1-type immune response and elicits a similar immune response in dogs. Clin. Vaccine immunology: CVI. 22, 274–281. doi: 10.1128/CVI.00653-14
PubMed Abstract | Crossref Full Text | Google Scholar
Rahimnahal, S., Yousefizadeh, S., Mohammadi, Y. (2023). Novel multi-epitope vaccine against bovine brucellosis: approach from immunoinformatics to expression. J. biomolecular structure dynamics. 41, 15460–15484. doi: 10.1080/07391102.2023.2188962
PubMed Abstract | Crossref Full Text | Google Scholar
Rahman, M. M., Hunter, H. N., Prova, S., Verma, V., Qamar, A., Golemi-Kotra, D. (2016). The Staphylococcus aureus Methicillin Resistance Factor FmtA Is a d-Amino Esterase That Acts on Teichoic Acids. mBio. 7, e02070–e02015. doi: 10.1128/mBio.02070-15
PubMed Abstract | Crossref Full Text | Google Scholar
Shi, K., Feng, S., Zhao, L., Chen, J., Song, W., Jia, Y., et al. (2024). N-glycosylation on hemagglutinin head reveals inter-branch antigenic variability of avian influenza virus H5-subtypes. Int. J. Biol. Macromol. 273, 132901. doi: 10.1016/j.ijbiomac.2024.132901
PubMed Abstract | Crossref Full Text | Google Scholar
Tan, W., Wang, X. R., Nie, Y., Wang, C., Cheng, L. Q., Wang, X. C., et al. (2012). Recombinant VirB5 protein as a potential serological marker for the diagnosis of bovine brucellosis. Mol. Cell. probes. 26, 127–131. doi: 10.1016/j.mcp.2012.02.003
PubMed Abstract | Crossref Full Text | Google Scholar
Tarrahimofrad, H., Zamani, J., Hamblin, M. R., Darvish, M., Mirzaei, H. (2022). A designed peptide-based vaccine to combat Brucella melitensis, B. suis and B. abortus: Harnessing an epitope mapping and immunoinformatics approach. BioMed. Pharmacother. 155, 113557. doi: 10.1016/j.biopha.2022.113557
PubMed Abstract | Crossref Full Text | Google Scholar
Xiong, X., Li, B., Zhou, Z., Gu, G., Li, M., Liu, J., et al. (2021). The virB system plays a crucial role in brucella intracellular infection. Int. J. Mol. Sci. 22, 13637. doi: 10.3390/ijms222413637
PubMed Abstract | Crossref Full Text | Google Scholar
Yin, Z., Li, M., Niu, C., Yu, M., Xie, X., Haimiti, G., et al. (2023). Design of multi-epitope vaccine candidate against Brucella type IV secretion system (T4SS). PLoS One 18, e0286358. doi: 10.1371/journal.pone.0286358
PubMed Abstract | Crossref Full Text | Google Scholar
Zai, X., Yin, Y., Guo, F., Yang, Q., Li, R., Li, Y., et al. (2021). Screening of potential vaccine candidates against pathogenic Brucella spp. using compositive reverse vaccinology. Veterinary Res. 52, 75. doi: 10.1186/s13567-021-00939-5
PubMed Abstract | Crossref Full Text | Google Scholar
Zheng, M., Lin, R., Zhu, J., Dong, Q., Chen, J., Jiang, P., et al. (2024). Effector proteins of type IV secretion system: weapons of brucella used to fight against host immunity. Curr. Stem Cell Res. Ther. 19, 145–153. doi: 10.2174/1574888X18666230222124529
留言 (0)