
記住我

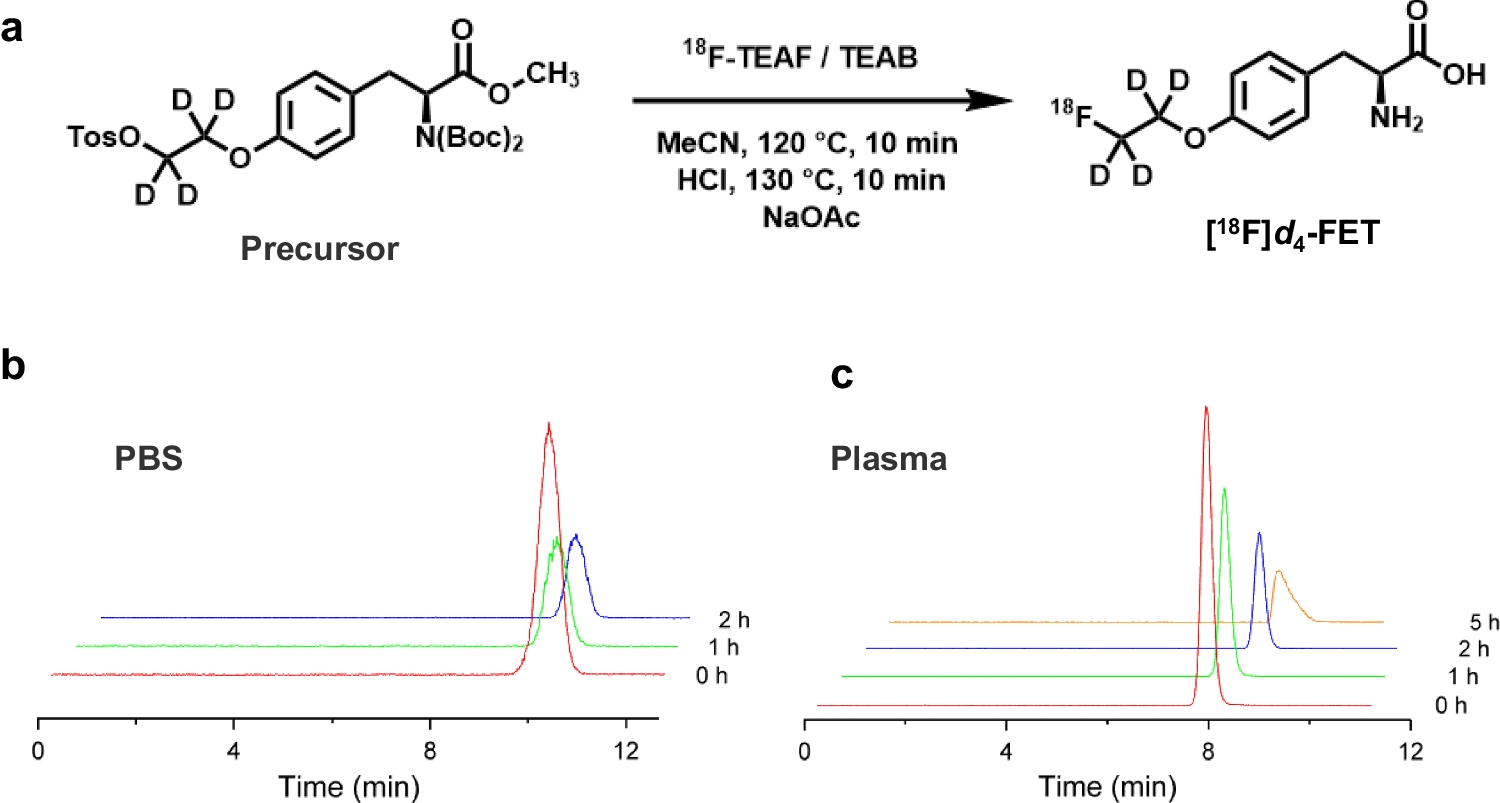



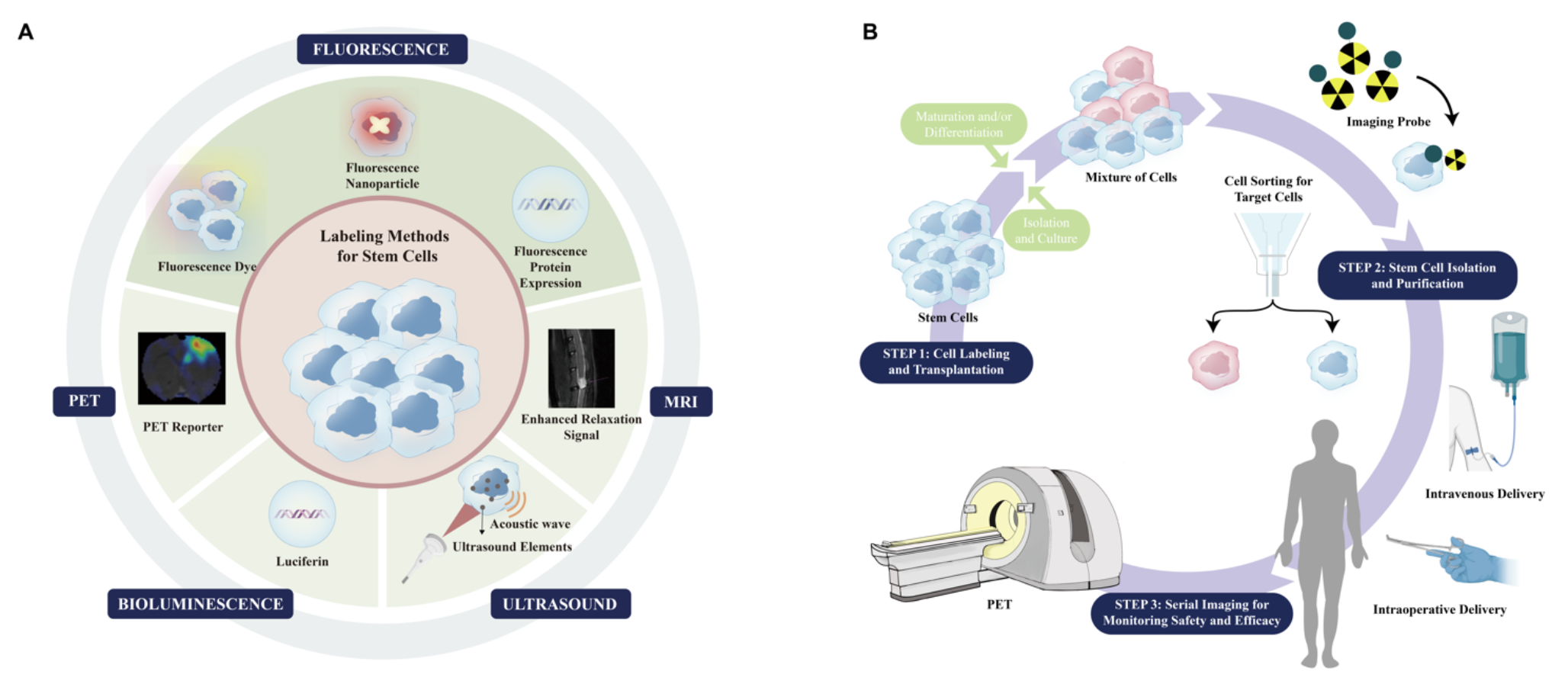
Advances in imaging, especially molecular imaging modalities (e.g., positron emission tomography (PET), magnetic resonance imaging (MRI), computed tomography (CT), ultrasound, and optical imaging means), have offered approaches to monitor various behaviors of grafted stem cells in vivo, including localization, survival, proliferation, migration, and differentiation (Fig. 1). According to the labeling strategy, the imaging approaches can be broadly classified into direct and indirect ones.
Fig. 1
Molecular imaging-based stem cell research paradigms. (A) Cell labeling strategies for stem cell imaging. The diagram shows labeling methods for tracking stem cells following in vivo implantation. Stem cells can be labeled using fluorescence (e.g., QDs, GFP and RFP), MRI (SPIONS), ultrasound (microbubbles), bioluminescence (e.g., Luciferin and Coelenterazine), and PET imaging (e.g., [18F]FDG, [18F]FESP and [18F]FHBG). (B) A step-by-step strategy for the routine application of stem cell regenerative therapy. Molecular imaging techniques represented by PET can provide spatiodynamic information for the functional recovery of dysregulated tissues. QDs, quantum dots; GFP, green fluorescent protein; RFP, red fluorescent protein; SPIONs, superparamagnetic iron oxide nanoparticles
Direct imaging strategies involves labeling stem cells with detectable markers, by incorporating fluorescent dyes, magnetic nanoparticles, or radiolabeled compounds directly into the stem cells before the transplantation. The main advantage of direct labeling is its ability to provide immediate insights into cell distribution, migration, and viability post-transplantation. For example, after the incubation with 18F-flurodeoxyglucose ([18F]FDG), it is possible to visualize the homing behavior of CD34-positive bone marrow cells in the border zone of myocardial infarction [3]. Similarly, ultrasmall superparamagnetic iron oxide nanoparticle labeled mesenchymal stem cells could be traced through MRI after transplantation in Alzheimer disease’ models [4]. It is noted that direct labeling should not affect the biological properties of stem cells (e.g. differentiation potential or proliferative capacity). Additionally, factors affecting the long-term imaging effect should be carefully assessed, such as the label dilution through cell division, the limited half-life of radiotracers, and non-specific phagocytosis of magnetic particles [5]. Despite these challenges, direct labeling remains a powerful tool for studying stem cell dynamics in vivo.
Unlike direct labeling, indirect imaging strategies track stem cells with reporter genes. By introducing reporter genes, stem cells can express receptors or proteins that bind to specific molecular probes, allowing for non-invasive imaging using PET, MRI, and fluorescence microscopy. For example, by incorporating triple fusion reporter gene (red fluorescent protein, firefly luciferase, and herpes simplex virus thymidine kinase) into the cardiomyocytes derived from induced pluripotent stem cells, the grafted cardiomyocytes can be imaged using fluorescence imaging, bioluminescence imaging, and PET [6]. The reporter gene strategy offers high flexibility and can be tailored to different experimental needs, making it a powerful tool for studying stem cell therapies and tissue engineering. Through using tissue-specific reporters, tissue-specific expression (e.g., neural differentiation) of implanted stem cells can be imaged [7]. Besides, compared to co-incubation induced direct labeling, reporter gene approaches enable long-term monitoring and lineage tracing of stem cells. By multiple genetic strategy, restrict reporter labelling to subset of endogenous stem cells is feasible, and is compatible with current intravital imaging approaches in the calvarial bone marrow, enabling the visualization of stem cell behaviors in niches [8].
留言 (0)