Bacillus species are known for their adaptability in extreme environments, especially in conditions with multiple stress factors, due to their ability to produce endospores. Endospore production has a high energy cost; under mesophilic conditions with a single stress factor, the percentage of Firmicutes present in the environment, including Bacillus, is relatively low. In terms of the total population, places with multiple stress conditions provide the highest proportion of spore-forming bacteria [27]. The strains from Mina B. cereus MSF4 and the Chichonal strains B. cereus S07C and P. dendritiformis S07D can be useful because of their unusually high alkaline tolerance, indicating broader environmental adaptability than typically observed within the pH range of 6–8 for Bacillus species. This makes the tested strains potentially better suited for bioreactor conditions, reducing the risk of viability loss [12]. Bacillus inaquosorum in particular also offers particular advantages as a PHB producer compared to previously reported bacteria. One notable strength of tis species first isolated from the Mojave Desert is its capacity to tolerate desiccation [28]. This trait is particularly valuable in solid-state fermentation systems, which are low-cost and require minimal water usage compared to traditional liquid fermentation. Solid-state fermentation systems, which utilize agro-industrial residues as substrates, can significantly reduce production costs, and the ability of B. inaquosorum to thrive in these conditions makes it well-suited for this type of production.
Compared with our other strain, P. dendritiformis S07 D, which had an exponential phase at the seventh growing hour, followed by a short stationary phase before declining around to hour 22, reports from Paenibacillus describe it as a species with quick growth times [29], which is consistent with what we observed in this work; quick growth times and high stress tolerance are two desirable factors for a metabolite producer.
The PHB synthesis process is traditionally attributed to carbon source mobilization and storage for further nutritional needs. Other physiological cell functions of PHB provide protection mechanisms against protein aggregation, reactive oxygen free radicals, and heat shock [30], as corroborated by the observed bacteria, as all the PHB-producing strains tested in this work were isolated from hostile environments where resistance to abiotic stress is needed, particularly heat shock resistance, as both the desert and the volcano were exposed to high temperatures. The strains isolated from Mina Nuevo León generally grew at the upper end of the temperature range of Bacillus spp. and were able to grow at 45 °C.
The strains B. cereus S07C and P. dendritiformis S07 D isolated from “El Chichonal” along with the B. cereus strain MSF4 from Mina had higher tolerances to alkaline conditions than did most of its members, which are already known for their high adaptability to a wide array of conditions [30]. In addition, the ecological distribution of Bacillus is strongly determined by ecological adaptations, in addition to other adaptations, including heat shock, osmotic shock by salts and different ranges of acidic and alkaline pH resistance [31], enabling the discovery of the environmental adaptations of P. dendritiformis S07D. The metabolic adaptability of these strains under stress conditions such as limited nutrients and variable temperatures indicates a robust physiological profile that may enhance PHB yield in non-optimal environments. This robustness not only lowers maintenance requirements but also potentially increases production efficiency by reducing the need for strictly controlled conditions. As result, these organisms could be a promising candidates for industrial-scale PHB production, particularly in economically viable setups where resilience and adaptability are key factors. Strain S07C of B. cereus in contrast had a slower growth rate under most conditions, suggesting that it was particularly adaptable to its microbial mat community, which opens the possibility of using both bacteria as consortium instead of pure cultures.
Among the seventeen isolated strains, seven tested positive for PhaC via PCR (41%) with the designed oligonucleotides, and a statistical report by Hassan et al. [7] revealed approximately 74 producers among 153 isolated bacteria (48%), indicating that approximately half of the wild-type bacteria can be PHB producers among cultivable organisms of the Bacillus genus. Hassan author along with Singh, et al. [5] reported that B. subtilis and B. megaterium are PHB producers; however, the NCBI database reports only the cereus group sequence of PhaC, although the functional region, in theory, should be preserved to maintain protein functionality. Paenibacillus dendritiformis is a bacterial species that has not been previously characterized as a potential PHB producer until this research; to our knowledge, one study reported it to be PhaC positive, but no further evaluations were made [32].
PHB production ability was detected in the strains from the two sampling locations, as confirmed by Sudan black staining on minimal medium agar when the strains were growing under stressful conditions (Fig. 2). The variation in the amount of PHB production between strains was significant even among members of the same species. The PHB production values obtained in this study were relatively high in the tested strains grown on orange peels, with 0.43 g/kg of PHB at the highest value for B. cereus MSF4 isolated from desert soil (Fig. 3), compared with the reported value of 0.025 g/L of strain OK2 isolated from commercial fermented soybean natto. According to the reports of Sukan et al. 2014, B. subtilis OK2, even when nitrogen starvation on the medium is a well-documented influencing factor on the PHB synthesis pathway, a certain amount of nitrogen is needed, pure orange peel is low in nitrogen, making it relatively unoptimized. The two strains of the same species had different outputs despite the same substrate being utilized. Most studies highlight producer strains grown on liquid media, including Bacillus subtilis NG220, which was isolated from sugarcane fields and was observed to accumulate 51.8% (w/w) of PHB via sugar industry wastewater supplemented with maltose and ammonium sulfate; Bacillus cereus SE-1, with an accumulation of 40% (w/w); and Bacillus sp. CS-605, with an accumulation of 33% (w/w) collected from a garbage dumping site and a lake, both of which were grown on minimal media supplemented with 1% dextrose [33]. and B. cereus NDRMN001, which were isolated from the soil of a polluted lake, with a reported PHB% production of 91.48% (w/w) when grown on selective media [6]. While those represent a higher PHB production compared to our organisms liquid substrates tend to be more expensive than solid-state fermentation alternatives because of the greater volume and water needed for submerged bioreactors [34], other potential advantages of using agro-industrial subproducts include improved cytocompatibility and biodegradability [35]. This finding also reinforces the great variability observed in the amount of PHB produced between strains. The strains isolated from environments with adverse conditions tended to have higher PHB production, with MSF4 from deserts being a better producer than OK2 and the highest overall producer coming from contaminated soil; in all cases, soil-isolated bacteria were observed to outproduce their liquid-born environment counterparts across the board. Stress sources rarely occur during isolation, heat often results in water scarcity, and the organisms that adapt to parallel or sequential threats are naturally selected to survive [36], as PHB confers protection to multiple stressors; at the same time, it should be overproduced in locations that warrant it as a survival mechanism. Protection from ultraviolet light, in particular, can be a determining factor for why isolated soil strains tend to be better producers than isolated liquid strains and are exposed to more direct sunlight.
Genome analysis from the DNA extracted from the Mina, Nuevo León, MSF4 strain, showed an average nucleotide identity of 99% to the B. cereus strain J2, identifying the species of MSF4 strain as B. cereus, likewise the S07C strain, isolated from the volcano had an ANI of 97% with B. cereus strain ATCC 14,579, other identified species include MSD1 having an ANI of 98.97 with B. inaquosorum A65 and S07D with an ANI of 98% with P. dendrtitiformis 2022CK-00834, all formed appropriately similar synteny plots (Figs. 4, 5 and 6) The presence of single copies of PhaC, PhaB, and the repressor PhaR downstream was confirmed in the genome of B. cereus strains, the group of genes phaQ, phaP, and phaJ were also found upstream forming two operons on both B. cereus MSF4 and B. cereus S07C(Figs. 7 and 8), these genes on that same configuration are present in on most Bacillus cereus sensu lato species like B. mycoides, Bacillus thuringiensis, and Bacillus anthracis with the same configuration, while Priestella megaterium another species know to have PHB producing strains like NBRC15308, lacks the phaJ gene [9].
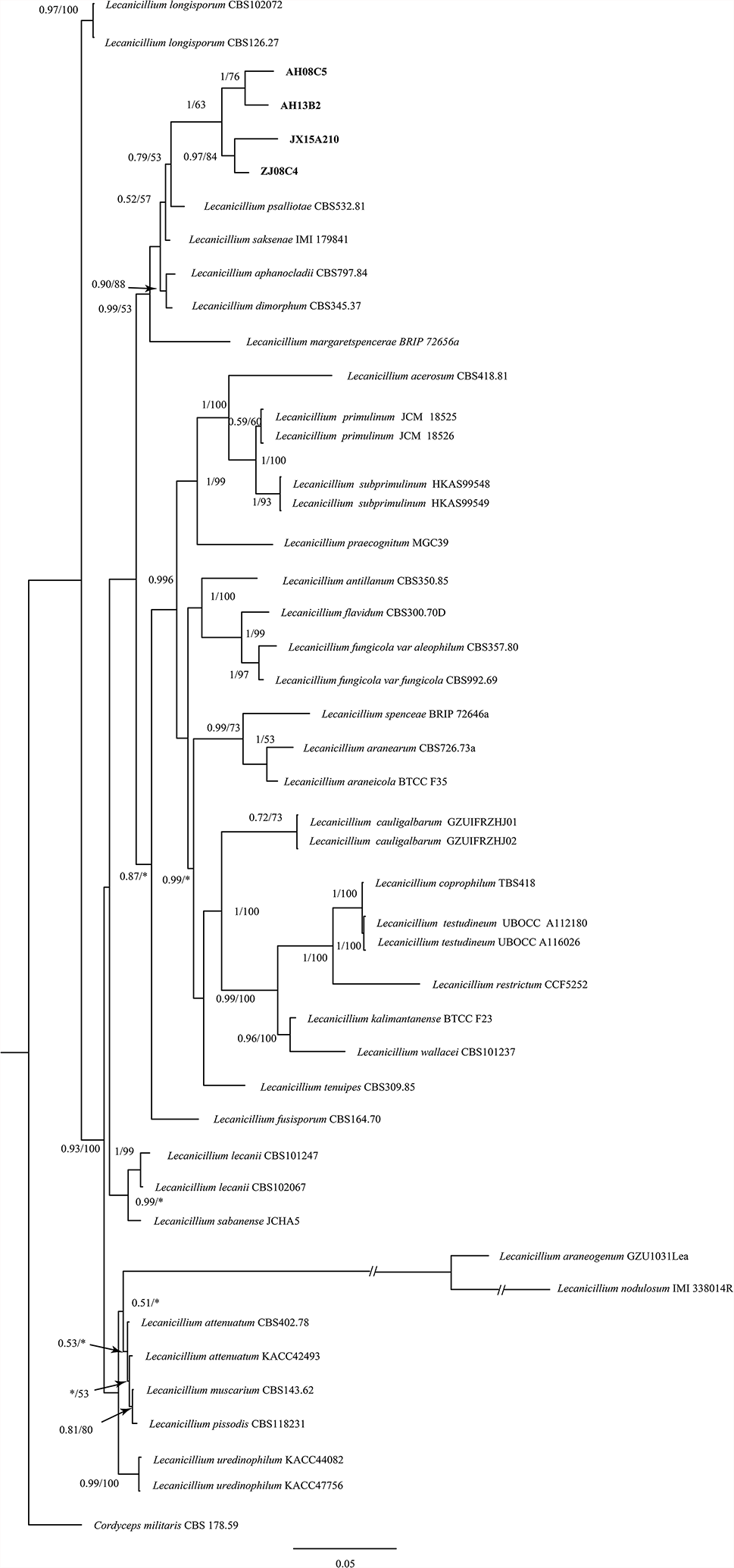
The phylogenetic tree generated via the sporulation control gene YteA (Fig. 9), which is a highly conserved gene in sporulation bacteria [16], generated well-defined branches for the cereus, priestella, subtilis and inaquosorum branches. PhaC additionally divides all the species into subgroups representing the PhaC class of the PhaC operon [37]. The gram-positive endospore species types of PhaC are Class IV for Bacillus, Class III for Haloferax, Class II for Pseudomonas and Class I for the golden standard of industrial production Ralstonia, which can be highly variable between strains of the same species. Despite being the usual way to perform phylogenetic identification, as attested by multiple authors cited in this paper, the 16 s marker proved to be ineffective in identifying individual species from the cereus group in this work owing to the extreme conservation levels between them resulting in a branch without observable separation between the different species under it, which has been noted by some authors, such as Okinaka in 2016, who reported that the group Bacillus cereus sensu lato, which includes B. anthracis, B. thuringiensis and other less known members such as B. mycoides, initially had enough similarity on the basis of early molecular approaches in which a taxonomic reclassification was discussed [38]. In contrast to the B. cereus group, the MSD1 strains of B. inaquosorum and the S07D strain of P. dendritiformis reached more than 98% ANI with respect to their respective reference strains and were separated into their respective branches on both the YteA and 16 S trees.
PHB operon comparison revealed a close relationship between PhaR and the citric acid cycle, and no direct interaction with nitrogen metabolism was found, with no direct utilization of nitrogen found in the function of the immediate PHA cluster genes. The effect of nitrogen was previously reported by Kumar and collaborators in 2009 [39], with this genomic study observing the interconnected pathway in silico and its interactions involved in the process reinforcing the hypothesis about the relationship between route activation and nitrogen starvation, which suggests an indirect mechanism regulated outside the studied operon (Fig. 10).





留言 (0)