
記住我
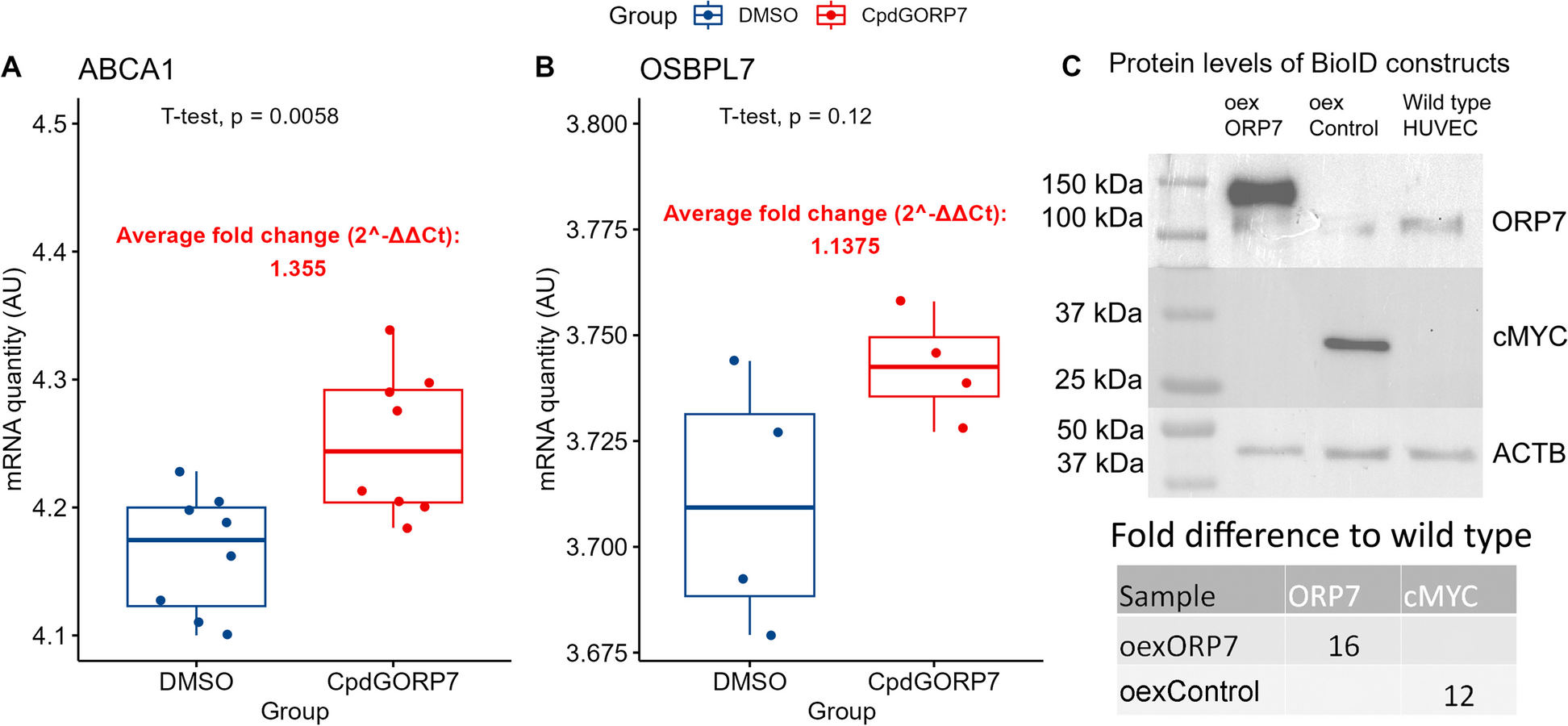







As the mobility of the honeybee queen has been suggested to play a critical role in the transmission of the queen pheromone and the rearing of new queens [5, 18, 33], we first investigated how the queen’s mobility patterns influenced how broadly the queen pheromone spreads through the colony. Previous studies have found that the movement of workers in several species of social insects [29], and also of honeybee queens [14] can be classified into two states: one characterised by local, area-restricted movement, and another characterised by wide-ranging movement. To identify these states, we used the moveHMM package [34] for R to fit a two-state hidden Markov model (HMM) to each of the 7 daily trajectories of the 10 queens. Across all 70 daily trajectories, the HMMs consistently revealed that queens frequently switch between two states, one associated with slow movement and large turn angles, and another associated with more rapid movement and smaller turn angles, which we labelled ‘stationary’ (S) and ‘travelling’ (T), respectively (Fig. 1a-b, Additional File 1: Fig. S1, Table S2).
Fig. 1
Quantifying the influence of queen behavioural state on pheromone transmission. a An example 24-hour queen trajectory on one side of the hive; travelling segments are shown in green and stationary segments in red. The black polygon indicates the boundary of the broodnest and the corner marked with a red arc represents the nest entrance. The blow-up shows the queen’s movement during an example 10-minute travelling bout (focal bout). Crosses indicate encounters with workers. b Behavioural state sequence of the queen during the same 24 hours, inferred by applying a HMM to the queen’s trajectory. The black line highlights the focal bout. c Queen movement speed during the focal bout (units are square-root-transformed body-lengths/sec). d Transmission sequence during the first minute of the focal bout. Each line on the y-axis represents a different bee, with the queen at y=0, and with the workers positioned according to the transmission order. Red and grey arrows respectively depict queen-to-worker and worker-to-worker pheromone transfers. e Individual-level load dynamics for the entire bout. f Snapshots of the spatial coverage of the informed workers on both sides of the hive. Points show locations of informed workers, lines show pairwise distances
We next investigated how the queen’s behavioural state influenced her physical encounters with workers. As the encounter rate of an agent within a population generally depends upon its speed [35], we expected that queens would experience a higher encounter rate when travelling than when stationary. Consistent with this expectation, and with previous results [19, 24], the encounter rate of queens was higher in the travelling than in the stationary state (Additional File 1: Fig. S2; Linear Mixed-Effect Model (LMM) with day & replicate as random effects, effect of state on encounter rate: d.f.=1, \(\chi ^2\)=123, p<0.0001). Furthermore, encounters were significantly shorter in the travelling than in the stationary state (Additional File 1: Fig. S2; LMM, effect of state on encounter duration, d.f.=1, \(\chi ^2\)=116, p<0.0001).
To explore how these state-dependent encounter patterns influence queen pheromone transmission, we developed an individual-based model in which queen pheromone spreads via both direct (queen-to-worker) and indirect (worker-to-worker) physical encounters (see Methods). The model was inspired by models of information flow [36] and disease transmission [37] over time-ordered contact sequences, and was parameterised using published empirical data on queen pheromone transmission [5, 16, 38]. In this model, we assumed that (i) there is a single inexhaustible and constant source of pheromone: the queen [16], (ii) pheromone transmission occurs during contacts in which a receiver bee antennates and/or licks the body of a pheromone-carrying bee (queen or pheromone-carrying worker), (iii) the amount transmitted depends on the contact’s duration and on the proportion of antennation versus licking carried out by the receiver, and (iv) the pheromone attenuates through decay [39] and ingestion [16]. A full description of the model and its parameterisation can be found in Supplementary Information (Additional File 1).
We then ran the transmission model on all bee-to-bee contacts that occurred between the start and end of each queen movement bout (Fig. 1c-e), recording the growth in the ‘audience’ of informed bees over time. A worker was defined as an audience-member if its current pheromone load exceeded a sensitivity threshold [38], regardless of whether the pheromone was acquired from the queen or from other workers (see Methods section). The audience size grew faster for travelling bouts than for stationary bouts (Fig. 2a). Furthermore, measuring the pairwise Euclidian distances between all bees in the audience (Fig. 1f) revealed that area covered by the audience increased more quickly for travelling than for stationary bouts (Fig. 2b). On the other hand, during stationary bouts the audience typically received a higher pheromone load than during travelling bouts (Fig. 2c). Considering that queens allocate roughly similar times to each state (time stationary, grand mean ± standard error = 58±2.4%, calculated across the 10 colony means), these results raise the possibility that queen’s frequent switches between bouts of stationary and travelling behaviour may help balance the size of their audience against the quantity of pheromone transferred to each audience member.
Fig. 2
Short-term dynamics of queen pheromone transmission. Panels show the growth in a audience size (proportion of the colony that are informed), b average spatial separation between informed workers, and c average pheromone load of informed workers, for stationary versus travelling queen bouts. Lines & shaded areas show the cross-colony grand mean & standard error. Each colony contributes a single value to the grand mean (n=10)
Colony-wide pheromone spread via direct and indirect transmissionTo evaluate how the queen pheromone spreads through the colony over longer timescales, we ran the transmission model on the full daily contact sequences. As young nurse workers are thought to be responsible for provisioning the queen, detecting the loss of a queen and raising new queens [14, 20, 40,41,42], we aimed to test whether different worker groups are more or less exposed to the queen pheromone. To do so, we applied a ‘soft’ community detection algorithm (FacetNet [43]) to the daily bee-to-bee contact networks [28]. This revealed that the contact networks contained three partially overlapping communities (Additional File 1: Fig. S3), namely, two nurse groups (\(N_A\) & \(N_B\), corresponding to the two patches of broodnest cells on either side of the hive [29]), and the foragers (F). As FacetNet uses scores in the range 0–1 to quantify the affiliations of each individual to each community, we defined nurses as bees that scored \((N_A + N_B) ^/_\), and foragers as those with \(F ^/_\). We then tracked the proportions of each group in the ‘informed’ state over the course of the day. Averaging across these daily growth curves revealed that after an intial phase of rapid growth, the proportion of nurses in the informed state plateaued at around 40%, whereas that of informed foragers never exceeded 10% (Fig. 3a, blue versus red lines). This indicates that queen pheromone is heterogeneously distributed across workers, with nurses – individuals that are the most involved in queen-related tasks – being more exposed than foragers.
Fig. 3
Indirect transmission enhances queen pheromone spreading, and nurses are more exposed than foragers. Growth curves show the proportion of informed bees as a function of time for daily contact sequences including both direct and indirect transmission (a, \(Q \rightarrow W + W \rightarrow W\)), or in the absence of indirect transmission (b, direct transmission only, \(Q \rightarrow W\)). Lines and shaded areas show the cross-colony grand means and standard errors, respectively. Each colony contributes a single value to the mean (n=10). Dashed coloured lines indicate the decay in the proportion of informed workers after the simulated removal of the queen at 12:00. The histograms in panel (a) show the post queen removal half-life distributions for informed nurses (blue) and foragers (red)
Previous work on how bees respond to the loss of the queen has focused on either worker behaviour (e.g., changes in aggression, mobility, and acoustic signalling) or colony-level processes (e.g., construction of replacement queen cells), and consequently the existing reaction time estimates vary from under an hour [5, 17], to close to a day [44, 45]. To evaluate whether our model produces realistic estimates for the dwindling – and eventual disappearance – of the informed workers after queen loss, we performed an in-silico queen removal experiment. Daily time-stamped contact sequences were edited to remove all contacts between the queen and workers that occurred after midday, and the transmission model was then run on these edited sequences. This showed that queen removal precipitates a rapid decay in the population of informed workers (Fig. 3a, dashed lines). Furthermore, there were differences in the half-lives of the population of informed bees for the two task groups: the population of informed foragers took 36±5 minutes to shrink to half its initial size (mean±standard error, n=10), whereas the informed nurses took 61±3 minutes (paired t-test, nurse versus forager half-life, d.f.=9, t=4.9, p=0.0009; Fig. 3a, histograms). As well as reinforcing the notion that changes in worker behaviour following queen loss are a direct response to the sudden reduction in queen pheromone flow over the contact network, the close agreement between these half-life times and previous reports of workers reacting in under an hour [5, 17] suggests that our simulation model generates reasonable approximations of real-world queen pheromone transmission.
Finally, to assess the importance of indirect transmission (or messaging) by workers in advertising the queen’s continuing presence, we created daily contact sequences in which only direct transmission was possible (Q\(\rightarrow\)W contacts only) by removing all of the worker-to-worker contacts. We then ran the transmission model on these reduced contact sequences, and compared the audience growth curves with those from the original, full contact sequences (Q\(\rightarrow\)W + W\(\rightarrow\)W contacts). These comparisons revealed that the absence of indirect transmission led to a reduction in the size of the queen-informed audience from \(\sim\) 25–40% (Fig. 3a, black line), to less than 10% of the colony (Fig. 3b, black line). Furthermore, in the absence of indirect transmission (Q\(\rightarrow\)W contacts only), the proportion of informed nurses dropped to \(\sim\)10%, whilst the proportion of informed foragers was close to zero (Fig. 3b, blue and red lines). This difference indicates that indirect transmission via workers plays a fundamental role in disseminating the queen pheromone throughout the colony, and thus that pheromone-carrying workers play a role akin to relay nodes in communication networks.
Workers orient towards high-load messengersAs our simulation results point to a potentially important role of messenger workers in spreading the queen pheromone, we next investigated the responses of receiver bees to pheromone-carrying workers. As the formation of the queen retinue is based on the ability of workers to sense and orient toward the queen [5, 22, 39], we expected to detect similar orientation responses towards workers carrying high pheromone loads. However, as we could not directly measure the actual load carried by each worker, we used two load proxies. The first was the time before and after each encounter with the queen; if queen pheromone is transferred during such encounters, then workers should orient more strongly toward nestmates that recently encountered the queen than toward the same bee in the moments just before it encountered the queen. As the following analyses concern how receivers orient towards individuals that are about to encounter the queen (i.e., enter the retinue), or that have recently encountered her (i.e., left the retinue), we refer to such individuals as ‘pre-retinue’ and ‘post-retinue’ bees respectively. We used the Rayleigh test statistic \(\rho\) [46] to quantify the orientations of receiver bees within two body-lengths of a pre- or post-retinue nestmate, and then to visualise how the queen encounter caused these orientations to change we constructed ‘difference maps’ comparing, for example the orientations in the first minute after the queen encounter with the last minute before the encounter (Fig. 4a-c).
Fig. 4
Workers orient towards nestmates that recently encountered the queen, or that have high queen pheromone loads. a-b Difference maps showing the change in the orientation strength of workers around a messenger bee (M). Worker orientation strengths are quantified using the Rayleigh test statistic, \(\rho\). Panels a & b show respectively the post-pre differences in receiver orientation strengths for the first and fifth minutes either side of the queen contact (i.e., \(\Delta \bar_ = \bar_ - \bar_\) & \(\bar_ - \bar_\)). Arrow plots show post-pre difference vectors, \(\Delta \vec v\), for areas to the front, the side, and behind the messenger. c Angular transect of the post-pre orientation strength differences as a function of the position of the receiver worker. Positions are measured clockwise relative to the heading of the messenger. Dots and error bars represent means & standard errors. d-e Difference maps comparing worker orientation strength toward a post-retinue messenger (M) with a given queen pheromone load, versus all post-retinue bees irrespective of their pheromone load (i.e., \(\Delta \bar_\)). Panels d & e show respectively the difference maps for messengers with a load in the top, and the second deciles (i.e., \(\rho _ - \rho _\) & \(\rho _ - \rho _\)). f Angular transect of the differences in receiver orientation strengths as a function of the position of the receiver
The difference map comparing the first minute after the queen encounter, with the last minute before it (Fig. 4a) contained a crescent-shaped zone around the head of the focal bee. Bees in this zone displayed a greater tendency to face toward a post-retinue bee than a pre-retinue bee (i.e., \(\Delta \rho _>\) 0). The minute-one difference map also contained a smaller zone around the abdomen in which nestmates tended to orient less toward a post-retinue bee than toward a pre-retinue bee (i.e., \(\Delta \rho _ <\) 0, blue arrows, Fig. 4a). These effects declined as the time since the queen encounter increased (Fig. 4b-c).
Our second load proxy was the dynamical pheromone load estimates for individual workers, produced by running the transmission model on the daily contact sequences. If bees react to pheromone-bearing nestmates in the same way that they react to the queen herself [39], then we expected that workers should orient more strongly towards post-retinue bees with high simulated pheromone loads, than towards post-retinue bees with low simulated loads. To test this, we used the dynamical estimates of the queen pheromone loads of each bee to assign post-retinue bees to one of ten groups (i.e., the 1st, 2nd, ..., 10th load deciles), and then calculated the difference between workers’ orientation strengths toward bees belonging to a given load decile, versus toward all bees. These difference maps showed that bees tended to face toward the head of a post-retinue nestmate with the very highest loads, that is, only when the load was in the top 10% of all bees in the colony (Fig. 4d-f). As with the previous time-based load proxy, the attraction was greatest for workers positioned around the head of the post-retinue bee. These behavioural assays are consistent with worker messaging, as workers preferentially reacted to nestmates that recently encountered the queen, and reacted more strongly the higher the pheromone load of the messenger.
Queen encounters induce temporary worker excitationWe next tested whether post-retinue workers exhibit increased mobility and increased long-range movement, which would enhance the dispersal of the queen pheromone throughout the hive [14, 23]. To do so we used individual-level comparisons of the movement of bees just before (‘pre-retinue’) and just after (‘post-retinue’) they encountered the queen.
Workers that had recently encountered the queen displayed increased mobility, moving significantly faster and straighter than they did before the encounter (Fig. 5a-b; Generalised Additive Mixed Models (GAMM) with day, replicate and worker ID as random effects, effect of ‘Post’ vs. ‘Pre’, instantaneous speed: \(\beta\)=0.007±0.00005, t=151, p<0.0001; unsigned turn angles, \(\beta\)=−0.024±0.0005, t=−47, p<0.0001). These differences persisted for \(\sim\)4 minutes after the end of the queen contact (Fig. 5d-f). Furthermore, compared to pre-retinue bees, post-retinue bees displayed a slightly higher probability of switching from one side of the frame to the other (Fig. 5c; GAMM, effect of ‘Post’ vs. ‘Pre’ on switching rate: \(\beta\)=0.000045±0.0000055 \(\text ^\), t=8.2, p<0.0001), although this effect disappeared after \(\sim\)2 minutes (Fig. 5f). Taken together these results show that queen encounters cause workers to exhibit both increased mobility, and an increased tendency to undertake wide-ranging excursions throughout the nest, consistent with the suggestion that pheromone-carrying workers act as messengers that actively modulate their behaviour to advertise the presence of the queen [14, 23].
Fig. 5
Physical contact with the queen induces worker excitation. a-c Worker movement before a physical encounter with the queen (‘Pre retinue’, negative times), and after it (‘Post retinue’, positive times). d-f Differences between the movement of pre- versus post-retinue workers. Differences were calculated by subtracting the observation at a given time before the queen contact, \(T=-t\), from the observation at the same time after the contact, \(T=+t\). Black crosses indicate cross-colony grand means & standard errors. Each colony contributes one value to the grand mean (n=10). Solid red lines indicate fits from the GAMMs described in the text. Blue arrows in d-f indicate times when the pre- and post-retinue GAMM fits were significantly different (at p<0.01)
Post-retinue worker excitation boosts queen pheromone transmissionA fundamental assumption of the worker messaging hypothesis is that the behavioural changes exhibited by workers after encountering the queen should boost the spread of the queen pheromone [14]. To test this, we ran the transmission model on all bee-to-bee contacts that occurred within a five minute window following each encounter between the queen and a worker, recording the growth in the number of informed bees over this period (Fig. 6a). Then we compared those post-retinue growth curves with growth curves obtained by running the model over the time-reversed [47] pre-retinue contacts, that is, those that occurred during the five-minute window immediately preceding the queen encounter. In agreement with the messaging hypothesis, the growth curves derived from the post-retinue contacts outstripped those for the pre-retinue contacts, and this was the case in all 10 colonies (Fig. 6c). Hence, encounters with the queen induce behavioural changes in workers which enhance the onwards transmission of the queen pheromone, for example via increased contact rates with uninformed nestmates.
Fig. 6
Comparison of forwards versus reverse-time transmission. a Example contact sequence covering a five minute pre- and post-retinue period. Individual bees occupy fixed positions on the y-axis, and physical encounters between pairs of bees are indicated by curved links. The queen is indicated in red. b Growth curves obtained from running the transmission simulation on the five-minute post-retinue contacts (‘forward’), and on the time-reversed five minute pre-retinue contacts (‘reverse’). c Relative differences between the audience size growth curves for the original post-retinue contact sequence versus the time-reversed pre-retinue contacts. Coloured lines indicate individual colony means. The black line and the shaded area indicates the grand mean & standard error, towards which each colony contribute a single value (n=10)
Messaging follows a stereotyped developmental trajectoryAs social insect worker behaviour typically follows a noisy, but stereotyped developmental trajectory [26, 28, 48], we next tested whether the expression of behaviours associated with messaging also exhibit predictable changes with age. To capture the multi-faceted nature of messaging, we defined a suite of six measures closely tied to messaging (‘messaging syndrome’), and used dimensionality reduction to quantify the developmental trajectory of these traits. As the most straightforward definition of ‘messaging’ requires that a worker frequently encounters the queen, our first two measures were (i) the active (i.e., beyond random) attraction of individual workers towards the queen [49], and (ii) the number of encounters between each worker and the queen. Queen attraction rapidly increased over the first few days of age, peaked at 3–4 days, and gradually declined thereafter (Fig. 7a). Similarly, newly-eclosed callow workers (0-days old) had few queen contacts, but over the first few days of life the number of contacts rapidly increased, reaching a peak at 3–4 days, and steadily decreasing thereafter (Fig. 7b).
Fig. 7
Expression of the messaging ‘syndrome’ follows a stereotypical developmental trajectory. a-b Active attraction of workers to the queen and contact rates between the queen and workers as a function of worker age. Points represent colony means, and point colours indicate colony identity. Solid lines indicate general additive models, fitted to the colony means. c-d Community affiliations in the daily contact networks as a function of age. e-f Spreading roles in the daily relay network according to age. Out-degree \(k_\) represents the number of nestmates an individual donates queen pheromone to, and degree difference \(\Delta k\) represents the out-degree minus the in-degree (number of nestmates an individual receives queen pheromone from). g-h Dimension reduction reveals that workers follow stereotypical developmental trajectories as they age. g Principal component analysis of the six behaviours shown in panels a-f. Background points represent a given individual on a given day. Foreground points show the mean for a given age cohort on a given day. Point colours represent colony identities. h Blowup of the rectangular area in g. Coloured arrows show developmental trajectories for each colony. The black arrow shows the ‘global’ trajectory. Coloured points show the inflection point for each colony. Points marked with an ‘x’ indicate the 3–5 day-old workers, which contribute most to feeding the queen and raising new queens. i-k The daily contact network (i-j) and the daily pheromone relay network (k) for colony 18 on 29/8/2016. All networks used the same layout, obtained by applying a force-directed layout algorithm to the contact network. Nodes represent individual bees, and the queen is indicated by the white node. Node size indicates age. In (i-j), edge thickness represents the number of pairwise encounters. In (k) edge weight & direction indicates pheromone flow. In (i), the nodes are coloured according to the community scores. The network is partitioned into three overlapping communities; a foraging community consisting mainly of older workers (red nodes), and two nurse communities consisting mainly of younger workers (blue and green nodes). In (j) nodes are coloured according to the nurse bridging score, \(H^\prime (N_A,N_B)\). Bees positioned at the overlap of the two nurse communities have high bridging scores. In (k) nodes are coloured according to the out degree. l The coarse-grained flow network. Node labels indicate the age in days. Weighted and directed edges indicate the net pheromone flow between cohorts. The donation hierarchy is indicated by the vertical positions of each node, as defined by the dominance ranks
As the social networks of honeybees are modular [29, 50, 51], the topological position of a bee, and in particular, its proximity to the queen, could influence how effective it is at spreading the queen pheromone. Given that queens were always most strongly affiliated towards one of the two nurse groups identified by the community detection algorithm, our third measure quantified how workers’ combined nurse affiliations (\(N_A + N_B\)) varied with age. Workers under 4 days of age were entirely affiliated to the nurse groups, but from \(\sim\)5 days of age, they began to transition into the forager group (Fig. 7c,i), with the S-shaped form of the transition closely resembling that seen in ant workers [28].
Given that nurses that were equally affiliated to both nurse groups might be well placed to act as messengers, we next used the entropy across their two nurse scores to calculate a ‘bridging score’, quantifying the extent to which each bee was positioned at the overlap between the nurse groups (\(H^\prime (N_A,N_B)\) , Fig. 7d,j, see Methods). Newly-eclosed workers had low bridging scores because they exhibited strong spatial fidelity to one of the two broodnests [14, 29]. However, over the first few days of life the bridging score rapidly increased, reaching a peak at \(\sim\)4–5 days, and declining thereafter. Therefore, 4–5 day old workers are best placed to convey the queen pheromone to those responsible for raising replacement queens.
As messengers should play a major role in disseminating the queen pheromone, the fifth and sixth measures quantified workers’ spreading roles over the daily pheromone relay networks (Fig. 7k, see Methods). Newly-eclosed workers donated to few nestmates (i.e. had out degree, kout∼100) and were net pheromone receivers (i.e., \(\Delta k < 0\), Fig. 7e-f). However, over the first few days of their life the number of nestmates to which they donated rapidly increased, so much so that that by \(\sim\)
留言 (0)