
記住我

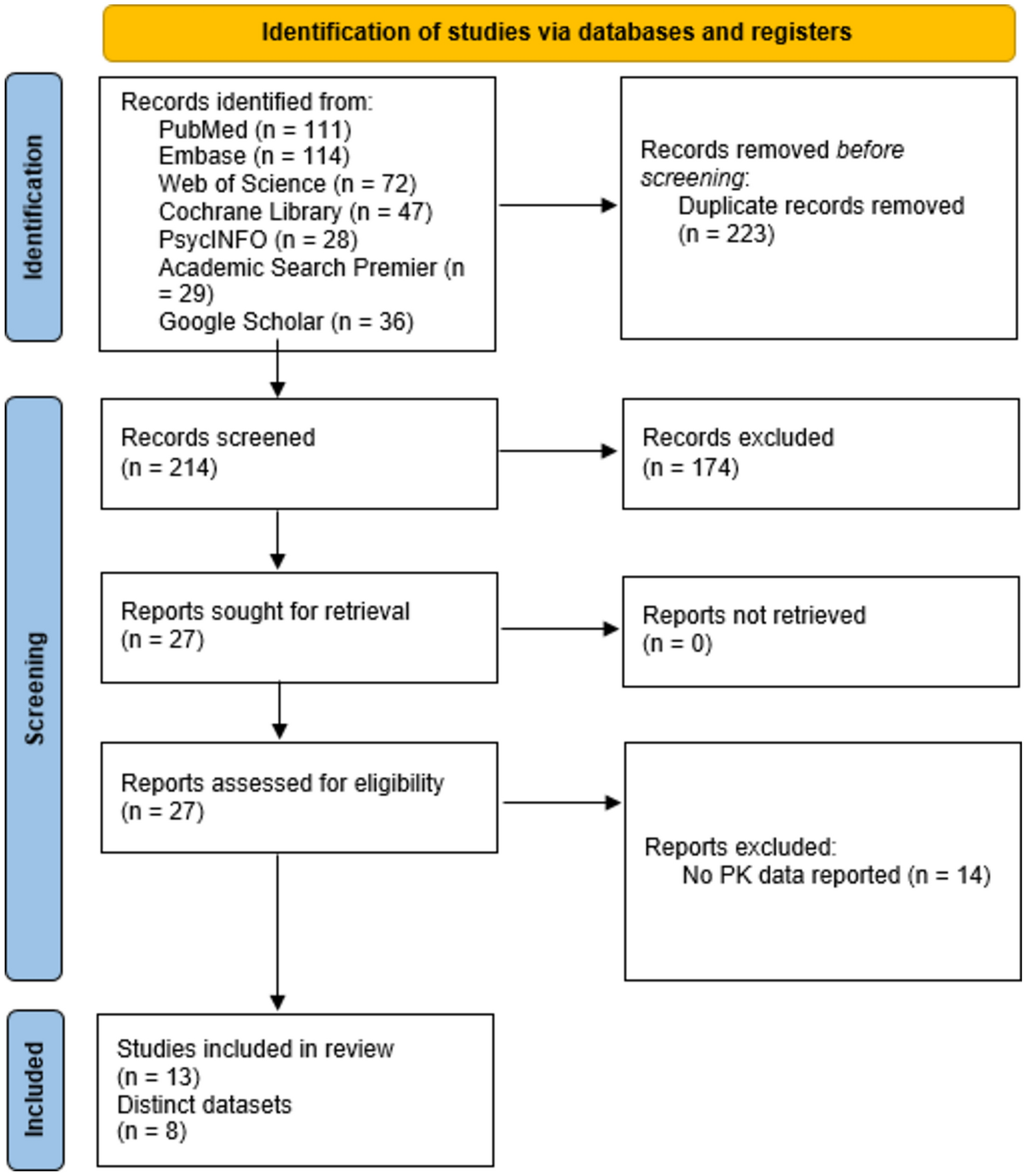




Scientists as far back as Galileo have been mystified by the relationship between the rate of physiological processes and the bodyweight of species [2]. Mathematically, the simplest relationship is a linear one. When applied to pharmacology, this would mean that the dose for a child of 10 kg would be one-seventh of the adult dose (assuming an average adult bodyweight of 70 kg). This was already shown in the nineteenth and twentieth centuries to lead to underdosing in children [3]. Non-linear relationships therefore seemed more adequate to capture the relationship between bodyweight and physiology.
The first widely adopted non-linear allometric relationship was proposed in 1838, when an animal’s BMR was found to be directly proportional to its body surface area [4]. As body surface area ∝ bodyweight2/3, it followed that BMR ∝ bodyweight2/3. Although empirical in nature, this is known as the surface area law. The surface area law has not remained free of criticism, as multiple assumptions are challenged, for instance, the assumption that the radiating capacity of animals is equal between species or that temperature of the skin is constant and equal for the entire body [5, 6]. In 1932, Max Kleiber empirically investigated the relationship between BMR and bodyweight of mammals spanning a larger bodyweight range and found an allometric scaling exponent (b) of 3/4 [7]. This empiric relationship has become known as Kleiber’s law.
Kleiber’s law did not replace the surface area law, and fierce discussions on which value is the ‘true’ allometric exponent (i.e., 2/3, 3/4, or 1) created distinct schools of thought, an observation reported as early as 1949 (Fig. 1) [8,9,10,11,12]. Regardless, a general power function for BMR was quickly accepted:
Fig. 1
Excerpt from Quantitative Relations in the Physiological Constitutions of Mammals by EF Adolph, published in 1949, highlighting the fragmentation of the scientific consensus on the scaling exponent and alluding to the existence of a distribution of exponents rather than a single universal value
$$\text=a\bullet }^,$$
(1)
where BW is the bodyweight of the species, a is an empiric coefficient, and b is the allometric scaling exponent.
2.2 West, Brown, and Enquist’s Theoretical Framework for 3/4 Power Scaling of Basal Metabolic Rate Between SpeciesMany studies sought to provide theoretical support for Kleiber’s empirically derived allometric exponent of 3/4, with the most influential being the mathematical framework by West, Brown, and Enquist (WBE) [13,14,15].
Central to the WBE framework, is that “Living things are sustained by the transport of materials through linear networks that branch to supply all parts of the organism” [14]. These transport systems are assumed to be of a fractal nature and to fill the entire volume of an organism. Two other essential assumptions are that the final branch of the fractal system is size invariant (i.e., independent of the animal’s size) and that energy minimisation (i.e., minimisation of the energy required for optimal flow through the network) is the driving force within the fractal network. For a complete mathematical derivation, the reader is directed to the original article [14]; here, we highlight the main aspects required to understand how WBE derived that b from Eq. 1 must be 3/4.
WBE use analogies to the vascular system to illustrate the theoretical concepts, for example, ‘flow’ through ‘networks’ is represented as blood flow through the vascular system (Fig. 2). The vascular system starts at the aorta and branches out until terminating at the capillaries (c). WBE define the aorta as level k = 0, which branches down in N branches. Any given branch at level k has a length (lk), radius (rk), and drop in pressure (Δpk). Each vessel branches into nk daughter branches. Blood flow (Q) in the aorta (Qo) is conserved throughout the system, so that Q0 = NkQk = NcQc. Because all nutrients required to sustain a BMR are transported via this flow, it follows that Qo ∝ BMR. Accepting the previously defined proportionality of BMR ∝ Mb, it must follow that Q0 ∝ Mb, with M referring to body mass. Moreover, as the total amount of capillaries (Nc) serve as the final supplier of nutrients for metabolic processes, Nc ∝ BMR and therefore it must follow that Nc ∝ Mb.
Fig. 2
Visualisation of the concepts and parameters used in the framework of West, Brown, and Enquist. Shown are a side view (top) and cross-sectional view (bottom) of the vascular system, which branches from the aorta (left) to the capillaries (right)
Second, the authors propose that the branching of consecutive levels of vessels can be characterised with scaling factors for the radius (r) and length (l). These scaling factors are the ratio between the radius or length of a level k + 1 and level k and are denoted β and γ, respectively. Because of the assumptions of self-similarity of the network and energy minimisation in the system, n, β and γ are independent of k.
Next, WBE define that the number of times the branches split (N) is logarithmically related to size. This is based on the relationship that the number of capillaries (Nc) is related to the number of vessels per node (n) and the number of levels (N) there are in total according to Nc = nN. Together with Nc ∝ Mb, the authors then derive that the number of levels must scale according to:
Here, M0 is defined as a “normalisation scale for M”, for which the precise definition remains unclear but, in a footnote, WBE mention that “normalisation factors, such as M0, will generally be suppressed.”
WBE subsequently derive the total volume of the system to be \(_=\sum_^__\), which, because of the fractal nature of the system, can be rewritten using the previously defined scaling parameters β and γ:
$$_= \frac^\right)}^-1}^\right)}^-1} ^_.$$
(3)
Using Eqs. 2 and 3, the assumption that Vb ∝ M, and the derivation that (γβ2)−N ∝ M, the last equation is derived:
$$b=-\frac\,n}\left(\gamma ^\right)}.$$
(4)
To show that b = 3/4, the ‘service volume’ is introduced, which is the volume of cells that a capillary provides with nutrients. All cells in a body are required to be serviced by capillaries, and it is implicitly assumed that there is no overlap in service volumes of capillaries. The diameter of a service volume on level k is defined as lk, meaning that the diameter of a service volume is equal to the length of the capillary by which it is serviced. From this, it follows that the total volume serviced by the network is approximated by 4/3π(lk/2)3Nk, which is increasingly more accurate with increasing k. From this equation and the self-preserving nature of the fractal network, it is derived that γ = n−1/3. A similar derivation for the cross section of the branches yields β = n−1/2. Filling out Eq. 4, it emerges that \(b= -\frac\,n}\left(^}*^\right)},\) which gives b = 3/4.
Of note is that the provided derivation is based on non-pulsatile systems. For pulsatile systems such as the mammalian vascular system, more mathematically intense and elaborate theoretical considerations are discussed, for which the reader is directed to the original publication by WBE [14]. Ultimately, this also yields b = 3/4.
2.3 Heated Discussions on WBE’s FrameworkThe theoretical framework by WBE to support Kleiber’s observation caused a resurgence of the interest in allometric scaling but also sparked a major debate. Other papers building on the concept of the fractal nature of biology, as well as empirical studies in favour of 3/4 scaling over 2/3 scaling, were published in the context of BMR [16, 17] but also to explain mitochondrial aerobic metabolism [18], plant growth [19, 20], rates of population growth [21, 22], and more [17, 23, 24], with all publications co-authored by one of the original WBE authors. However, others pointed out that the WBE framework was developed to explain but not predict an empirically derived exponent of 3/4 and as such provides one possible explanation but not conceptual proof. For instance, in 2016, Kwak et al. provided a theoretical framework to explain 2/3 scaling [25]. In addition to this, other topics remain on which consensus between proponents and opponents of the framework has not been reached. Table 1 summarises the core assumptions of the framework by WBE as well as a sample of counterarguments for each assumption that are further discussed below.
Table 1 Overview of the core assumptions in the theoretical framework developed by West, Brown, and Enquist (WBE) to provide an explanation for Kleiber’s law, including a summary of the theoretical counterarguments and empirical observations that disprove the assumptions2.3.1 Theoretical Critiques on the WBE Framework2.3.1.1 Service VolumesIn 2004, Kozłowsky and Konarzewski (K&K) published a paper in which they stated that “WBE’s model is mathematically incorrect” [26]. According to these authors, this bold statement follows from the definition of the service volume. An important assumption of WBE states that the capillaries are size invariant. The capillaries supply a service volume with a diameter of the length of a capillary (lc). K&K subsequently argue that, because lc is size invariant, the number of capillaries (Nc) must be directly proportional to the total body volume (W), which is directly proportional to body mass (M). So, if M ∝ W and W ∝ 4/3π(lc/2)3Nc, from the size invariance of lc, it must follow that Nc ∝ M and not Nc ∝ M3/4. K&K therefore conclude that either b must be equal to 1 or the assumption on the size invariance of lc must be violated. K&K continue by showing that, aside from this theoretical inconsistency, the derivation of b = 3/4 is incorrect. WBE make use of the relation (γβ2)−N ∝ M, which itself is based on the proportionality of total blood volume (Vb) and mass, Vb ∝ M. K&K state that, according to capillary size invariance, the blood volume present within capillaries, which is VcNc, must be proportional to M, not total blood volume Vb as suggested by WBE. Vb should increase supralinearly with M, as more fluid is present within vessels not supplying any tissues. In that case and based on numerical examples, K&K state that (γβ2)−N ∝ M4/3 and not (γβ2)−N ∝ M as proposed by WBE. Overall, the derivation of b = 3/4 based on the erroneous (γβ2)−N ∝ M therefore does not hold in their opinion.
Hereafter, we refer to WBE’s response to K&K as WBE2005 and statements from the original publication as WBE1997. In WBE2005, the authors claim that “[K&K] misread our paper, misrepresented our model, and made serious mistakes” [27]. They state that K&K interpreted the size invariance described in WBE1997 to imply that a constant volume of tissue will be supplied by a capillary [26, 27]. However, WBE2005 indicate that, in their framework, the service volume scales to body size via the relationship M1/4. According to WBE2005, K&K take size invariance to mean that Nc must scale isometrically with bodyweight. However, WBE2005 argue that the size invariance of the capillaries and the way that Nc scales are independent. Quickly after, K&K responded [28], saying WBE did not manage to accurately refute their critique and that, unless b is equal to one, a logical fallacy remains within their framework. They restate that Nc ∝ Mb (as stated in WBE1997) is needed to satisfy Qo ∝ BMR. However, Nc ∝ M1 is required to allow size invariant service volumes. According to K&K, this fundamental inconsistency can only be solved in one of three ways: (1) assuming density of tissues increases with body mass with an exponent of 1 − b (which K&K deem unlikely), (2) giving up the assumption of capillary size invariance, or (3) assuming b = 1. K&K note that solution (2) is the only one supported by empirical evidence [29,30,31]. Overall, the two sides remain unreconciled. The discussion illustrates the complexity of the assumptions introduced by WBE, and the difficulty in their interpretation.
Etienne et al. tried to offer a solution to the impasse [32]. According to these authors, the disagreement is caused by how a space-filling network is interpreted. WBE interpret this as a preservation of 4/3π(lk/2)3Nk for every level k, simplified to Nklk3 for ease of reading. According to Etienne et al., WBE assume Nklk3 to be proportional to the service volume of all vessels. WBE treat the assumption of a space-filling fractal such that the entire volume of an organism is filled with service volumes. This is also corroborated by others: “[…] network extent and structure must be such that its terminal points can service (and thus cover) the entire body volume” [33]. However, Etienne et al. argue that, if the service volume is proportional to Nclc3 and this volume fills the entire body, it will scale with body size as 4/3π(lk/2)3Nk ∝ M, as concluded by K&K. The solution to this is to forgo the assumption that 4/3π(lk/2)3Nk corresponds to the entire body volume and adopt the assumption that the quantity 4/3π(lk/2)3Nk is preserved through all levels k. The extent to which this resolves the discussion on size invariance of capillaries and service volumes is questionable, as the main concern K&K posed was related to the incompatibility of size invariance of capillaries with the service volumes, not the relationship between service volumes and total body volume itself.
2.3.1.2 Biological Relevance and Universality of the WBE FrameworkWBE present their framework to apply to a wide range of supply networks, including mammalian vasculature and respiratory systems, plant vascular systems, and tracheal tubes of insects [14]. However, K&K point out that assumptions by WBE on branching of supply vessels do not take the organism’s geometry into account [26]. For instance, for mammalian vasculature, this implies that vessels running from the heart to abdominal organs would split similarly to those running to limb extremities, the reality of which K&K question. The mesh-like organisation of capillaries is also not represented by the hierarchical branching presented in WBE. Additionally, K&K note that WBE’s assumptions do not consider differences in tissue perfusion in mammals [26]. As not all capillaries can be completely filled at the same time, because of a lack of blood to do so, K&K argue that the supplying system adjusts to cover tissue demand, rather than the supplying system limiting BMR. This explains why an animal’s metabolism increases when they become active, which would not be possible if the delivery of nutrients was rate limiting at rest. Time-varying energy consumption by organs and acclimatisation to different climates can also lead to physiological differences in tissue perfusion, therefore one can question whether the assumption of one capillary exclusively servicing one service volume is always correct. Finally, K&K highlight that the assumption by WBE of a constant number of daughter branches at each node of the network is violated by empirical findings that, depending on the intensity of metabolism, the proportion of capillaries opened to blood flow varies [28].
Regarding plant vasculature and tracheal systems of insects, K&K have raised further questions on the biological validity of assumptions in those specific contexts, disputing the proposed universality of the WBE framework. Arguments on these topics have been going back and forth [27, 28], but also here the different authors do not settle their disputes. Others have also critiqued the application of the WBE framework to plants, concluding that the framework is too simplistic to apply to vascular plants and critique it for not taking important ontogenetic changes into account [34].
2.3.1.3 Mathematical CritiquesAside from K&K, others have raised doubts about the mathematical validity of the WBE framework. Dodds et al. have analysed theoretical derivations of the 3/4 power scaling law [35] and assessed the assumption made by WBE that the networks are fractal in nature. In WBE1997, the network impedance (or resistance, Z) of pulsatile systems is minimised via a so-called Lagrange multiplier method. The fractal nature of the system is partly described via this minimisation procedure. However, Dodds et al. show that Z indeed is minimised when γk = βk = nk−1/3, but their calculations show it is not necessarily level independent and therefore does not prove that the network must necessarily be fractal.
Chaui-Berlinck thoroughly assesses the mathematical support for the assumptions made by WBE1997 in a pulsatile system [36]. WBE show that β = n−1/2 in non-pulsatile systems, which is essential to be able to derive that b = 3/4. However, Chaui-Berlinck illustrates that this pivotal relationship is not proven in a pulsatile system and that derivations of this relationship are not in line with empirical findings. Chaui-Berlinck also concludes that the relationship of Vb ∝ M has never been proven and investigates this assumption in a pulsatile system by considering energy minimisation to conclude that “WBE cannot prove their claim that a linear relationship between fluid volume and body mass is a natural consequence of geometric/impedance constraints in living beings”. Lastly, Chaui-Berlinck argues, like K&K, but through different reasoning, that, because the service volume is calculated as 4/3π(lc/2)3 and the capillaries have an invariant length, the service volumes must also be an invariant unit, meaning the relationship Nc ∝ Mb can only be true if b = 1. Much like the critique by K&K, the report by Chaui-Berlinck sparked a discussion between the author and WBE [37, 38], in which the authors do not reconcile their differing views.
In a reanalysis of two theoretical frameworks, including the WBE framework, that aim to explain b = 3/4, Makarieva et al. discuss the implications of the assumed proportionality Vb ∝ M. They find that the framework would only yield B ∝ M3/4 if M ∝ l04, with l0 being the linear size of the total volume of the fractal network [39]. A common assumption would be l0 ∝ L (body length), as in the framework by Banavar et al. [15], which also aims to a theoretical explanation for b = 3/4. This is deemed problematic because empirical observations show M ∝ L3 in mammals. However, WBE circumvent this problem by introducing l0 as independent from L. But, as noted by Makarieva et al., this creates the inherent problem that the network must stretch along the whole body (L) to supply all parts of the organism, instead of being restricted to its own spatial scale l0. This yields two linear scales within one organism, which is shown by Makarieva et al. to not be supported by empirical evidence. They show that the assumption Vb ∝ M causes the WBE framework to predict the length of the aorta to scale with M1/4, whereas available evidence shows it to scale as M1/3. Also shown is that the assumption of M ∝ l04, which is required if Vb ∝ M, would lead to the spatial volume of the network becoming increasingly large with smaller body sizes, even giving rise to volumes of the system larger than the body volume itself [39]. They conclude that “the WBE model cannot account for the scaling exponents in animals remaining simultaneously within the domains of biological and physical plausibility and mathematical coherence.”
Self-similar properties of biological transport systems have also been questioned on biological grounds by K&K. In response, Etienne et al. found that a scaling exponent of 3/4 can be derived without the assumption of self-similarity in the fractal networks [32]. However, their redefined WBE framework becomes even more theoretical in nature, including the introduction of topological quantities that “will be helpful in our derivation”, but for which it remains unclear what biological features they represent. As such, it is difficult to scrutinise the biological implications.
Apol et al. reanalysed WBE and critically discussed five assumptions. One critique is that energy dissipation has only been partially minimised by WBE. They show with further optimisations that the optimal branching ratio is 1, meaning that the optimal network predicted by the WBE framework is one large vessel that runs through the organism [40]. The scaling exponent would then be 0. By relaxing some of WBE’s assumptions, the framework predicts isometric scaling of BMR with body mass (i.e., b = 1), which they recognise to be incompatible with observed scaling exponents. They conclude that the WBE framework is not suited to ‘explain’ Kleiber’s law and suggest the reason for it may have to be sought outside of the realm of flow-limited network theory.
Finally, Savage, Deeds, and Fontana performed a thorough reanalysis of the WBE framework and a re-examination of the assumptions. They investigated network size, branching numbers, and when pulsatile flow (area-preserving branching) switches to smooth flow (area-increasing branching) and delineate how these aspects influence the scaling exponent [33]. Their main conclusion is that the 3/4 exponent derived by WBE only holds if the limits of mass, network size, and metabolic rate are approached. After finite-size corrections, a scaling exponent of 0.81 is predicted by the WBE framework.
2.3.2 Empirical Findings to Disprove Kleiber’s Law and the WBE FrameworkIn addition to theoretical discussions, empirical evidence to accept or reject Kleiber’s law and the WBE framework has also been reported. In 1982, Heusner concluded Kleiber’s law to be a statistical artefact and challenged both the derived allometric exponent of 3/4 and the assumed constancy of the mass coefficient (a in Eq. 1). He found the allometric exponent to be 2/3 for seven different species and the mass coefficient (a) to change with body mass and between species [9]. As such, Heusner concluded large and small mammals to lie on parallel regression lines with a slope of 2/3 [10].
In a thorough reanalysis of multiple datasets, including Kleiber’s original dataset, Dodds et al. found little statistical evidence to reject the 2/3 scaling law in favour of the 3/4 law [35]. Furthermore, they were unable to statistically reject 2/3 as the scaling exponent for mammals with a bodyweight smaller than 10 kg and for birds. For larger mammals, deviations from 2/3 were more common, but little evidence exists for accepting 3/4 over 2/3 as an allometric exponent.
White et al. [8] argue that Kleiber and others used datasets that unfairly represent species. For example, Artiodactyla are overrepresented, which, because of their relatively large weight, can lead to an overestimated exponent. They also argue that differences in body temperature between species should be acknowledged. Taking these considerations into account, their analysis of the correlation between bodyweight and BMR in 619 species from 19 mammalian orders yielded an estimated exponent of 2/3.
Hayssen and Lacy also critique Kleiber’s dataset, as they find it too small and unrepresentative of mammals, with 9 of 12 included species being domestic populations that do not live under natural conditions [41].
2.3.3 Lack of Universality of the Allometric Exponent for Basal Metabolic RateAlthough numerous discussions focused on whether 2/3 or 3/4 should be accepted as the universal scaling exponent, many have challenged the existence of a universal relationship altogether. For example, Hayssen and Lacy were unable to identify a single allometric exponent to describe the relationship of BMR and body mass in mammals [41]. White et al. published a meta-analysis on 127 allometric exponents for scaling BMR and found high heterogeneity of the exponent, suggesting that a universal scaling exponent does not exist [42], whereas Bokma’s scaling of BMR in fish indicated the same even for within-species scaling [43]. Heusner’s scaling of BMR within dogs resulted in a scaling exponent of 0.885, further refuting the (within-species) universality of 3/4 or 2/3 [44]. Clarke et al. found that the allometric exponent of species with a lower bodyweight was 2/3 and that it moved asymptotically to a value of around 3/4 with increasing bodyweight [45], which is in line with the results reported by Dodds et al. [35] and McNab et al. [46]. A dependency of the scaling exponent on bodyweight was also highlighted by Heusner [44] and reported by Lovegrove across 487 mammalian species [47]. Numerous other papers have proposed species characteristics that show correlations with the allometric scaling exponent for BMR, including taxonomical class or phylogenetics [48, 49], physical constraints of an organism [50, 51], zoogeographical zones, whether a species is a predator or not, and the mobility of animals [47]. In a series of papers, Kosłowski proposes how evolutionary principles can drive variability in the allometric exponent [52, 53]. In line with this view, in a recent review on the variability of allometric scaling exponents, Glazier shows the wide range of exponents that have been reported within different taxa [54]. They indicate a switch from a ‘Newtonian approach’ to allometric theory, in which physical explanations are sought for a universal law and variability is of minor importance, to a ‘Darwinian approach’, in which variability is considered of primary importance and for which evolutionary explanations can be found. As such, a shift can be seen in allometric theory no longer aiming to find physical explanations for a universal scaling law but instead explaining the variable nature of the allometric exponent due to “adaptable phenotypic plasticity and genotypic evolution,” which are impacted by biological and ecological influences [54].
2.4 Conclusions for PharmacologistsThe background of allometric theory is vast and has been summarised here to provide pharmacologists with the highlights of the efforts to concretise Kleiber’s empirical observations into mathematically based theories and of the theoretical and empirical discourse surrounding these theories. Readers interested in obtaining a more substantial understanding of this field are encouraged to read the extraordinarily in-depth reviews by Gould [
留言 (0)