
記住我
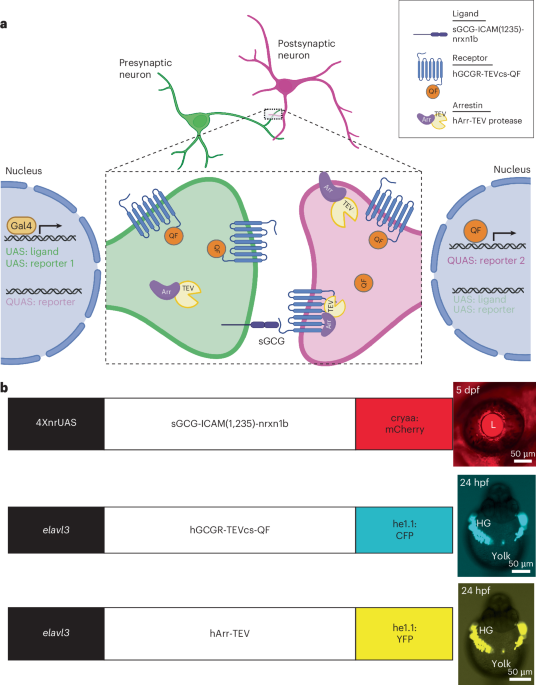
One major challenge of studying foveolar representation is minimizing eye movements. Two macaque monkeys were trained for several months to maintain stable performance with little body motion (Fig. 1a) and few saccades (Fig. 1b, left) during scanning. Only data in which fixations remained within a 1° radius window >85% of the time and the center of mass of the eye trace remained within a 0.5° radius of the fixation point were used (Fig. 1b, right). With sufficient trial numbers, consistent and reproducible fMRI results were generated across different sessions (Fig. 1c). These procedures made direct retinotopic and foveolar mapping possible (Fig. 1d,e).
Fig. 1: Acquisition stability.
a, Left, head motion in six dimensions: roll, pitch, yaw, A–P (anterior–posterior), R–L (right–left) and I–S (inferior–superior). Right, head displacement: disp, displacement relative to the initial frame (mean = 120 µm); Δdisp, difference between two sequential frames (mean = 80 µm). Data were averaged across eight sessions in monkey E. b, Eye fixation behavior. Left, eye position trace in one run. Red and blue lines represent the x and y eye position coordinates, respectively. Two red arrows denote blinks. Note: eye blinks did not cause disruptions of the BOLD signal. Right, all eye positions in four runs (250 Hz, 4 × 280 s). Eye fixations were within a 0.5° radius 94% of the time. Right, despite occasional saccades, across four runs (each trial: 20 s blank and 20 s visual stimulus; seven trials per run; a total of 280 s), the center of mass of the eye trace was within a 0.5° radius of the fixation point (central dotted red circle). With sufficient training, the monkeys maintained stable performance with little body motion and few saccades during scanning. c, BOLD signal time course. Left, time course of activated voxels during eye fixation in b (n = 348 voxels). Right, average BOLD time course. Yellow shading denotes the stimulus on period. d, Map stability. Maps obtained from 3, 7 and 28 trials. Left inset, visual stimulus: 1° and 5° eccentricity rings. Activation clusters remained stable across trials, indicating stable eye fixation. Yellow arrows, V1; red arrows, V2. Right, each dot is a measurement of the shift in center of mass between activations from 7 and 28 trials. For each of the activation clusters, six measurements were taken from six sequential slices. e, Increased trial number, from 7 (middle panel) to 28 (right panel), reveals additional visual areas. Left, section from D99 atlas. Activations in V1 (1° and 5°), V2 (yellow circles), V3 (green arrow), V4 (blue arrow) and TEO (red arrows). Same as in d. Map thresholds were significant at P < 10−3 according to a one-sided t-test. All data are presented as the mean values ± s.e.m.
Foveolar mappingTo determine the visual representation of the foveal region we conducted visuotopic mapping using very narrow 0.15° lines and arcs during excellent visual fixation (Fig. 2). Maps of VM and HM marked the boundaries dividing cortical areas V1, V2, V3 and V4 (Fig. 2a,b). We also mapped isoeccentricities using arcs ranging from 1° to 6° isoeccentricity and, within the central 1°, several finely distinguished isoeccentricities (Fig. 2c,d) (summarized for each monkey in Fig. 2e,f).
Fig. 2: Mapping visual cortical borders by mapping HM and VM and isoeccentricity mapping.
a,b, Stimuli for imaging VM and HM, in monkey E (a) and monkey J (b), comprised horizontal and vertical 0.15°-wide bands containing alternating saturated red and blue checkerboards (insets). All images were acquired within the same sessions but illustrated separately for each area (Methods). For each monkey, the four panels on the left were overlain in the rightmost images. Black and white dashed lines denote the VM and HM, respectively. LS, lunate sulcus; IOS, inferior occipital sulcus; STS, superior temporal sulcus. ECS, ectocalcarine sulcus. Monkey E, P < 10−3; monkey J, P < 10−9 (one-sided t-test). c, Isoeccentricity maps from monkey E. A paired stimulus paradigm (six sets of paired rings: 0.4° and 2°, 0.4° and 4°, 0.6° and 3°, 0.8° and 4°, 1° and 5° and 1.2° and 6°) reduced the number of stimulus conditions needed by half and further confirmed that the maps were stable across sessions (for example, similar 4° activations were obtained in 0° and 4°, 0.4° and 4° and 0.8° and 4° maps). White circle, intersection location between isoeccentricity and meridians. d, Isoeccentricity (0.8° and 4°, 1° and 5° and 1.4° and 7°) maps from monkey J. e,f, Summary of findings in c (e) and d (f).
We then used small stimuli placed at the fixation location to precisely identify foveolar locations. These comprised alternating saturated red and blue flashing spot stimuli (Methods); to examine the consistency of activation locations, three spot sizes were used (monkey E: 0.4°, 0.6° and 0.8°; monkey J: 0.6°, 0.8° and 1.0°). Given the distribution of eye positions during the fixation task, the most frequently activated locations are at the very center; thus, the foveolar center should be at the location of voxels with the highest statistical significance. Given the simple flashing visual spot stimuli used, the activation was strongest for early areas V1 and V2 and weaker at the more anterior V3, V4 and TEO locations; the maps were examined using thresholds (three P values per area) appropriate for the activation levels of each cortical area. Below, we describe how each foveolar location was identified in monkey E (Fig. 3). Findings for monkey J are presented in Extended Data Fig. 1. Division of voxels into V1/V2, V2/V3, V3/V4, V4/TEO and TEO/FST (fundus of the superior temporal visual area) regions were guided by locations of VM and HM (black and white dotted lines, respectively, shown in overlay in Fig. 3a, last column) and slice views (Extended Data Fig. 2). Below, we describe each foveolar location.
Fig. 3: Determining locations of foveolar representation.
a, Activations to foveated small spot stimuli (monkey E: 0.4°, 0.6° and 0.8° flashing saturated red and blue squares alternating at 3 Hz, shown in the first, second and third row, respectively). The activation maps to each stimulus are shown in each column (TEO/FST, V4/TEO, V3/V4, V2/V3 and V1/V2). Each panel presents significant voxels at each of three thresholds (P values according to a one-sided t-test are color-coded: orange, lowest; green, middle; blue, highest). For each visual area, the center of highest significance is consistent across spot sizes (colored dashed lines). The last column consists of the set of all foveola (colored dashed circles, overlay from four leftmost columns). Black and white dotted lines denote the VM and HM, respectively. Gyri, light gray; sulci, dark gray. b, Foveolar activations in monkey E, right hemisphere (same as response to 0.6° in a). c, Foveolar activations in monkey E, left hemisphere. d,e, Foveolar activations in monkey J’s left hemisphere (d) and right hemisphere (e) (Extended Data Fig. 1). In b–e, small colored circles denote the foveolar activations in each cortical area. Lavender dotted oval, foveolar core.
V1/V2 (fourth column)In monkey E, each of the three spot sizes activated a similar region (compare three rows in Fig. 3a). Examination of the activations at the lowest threshold (V1/V2 column, orange, 0.4° stimulus) revealed patchy activations across a centimeter-sized region; this activation region is anisotropic (more elongated along the VM axis), consistent with previously observed anisotropies in V1 and V2 because of functional rerepresentations21 and/or anisotropic representations along cardinal axes22. Although eye fixations away from the fixation location were relatively few (set by our criterion of <15% of fixations outside of a 1° radius), we cannot exclude the possibility that they contributed to some of the activation seen at lower thresholds. At higher thresholds, quite focal activations were revealed across smaller regions of the cortex (middle threshold, green: 3–8 mm; highest threshold, blue: 1–4 mm). We defined the ‘most foveal’ locus as the lateralmost (because the retinotopic center is located laterally on the macaque visual cortex) activation at the highest threshold (blue) that was consistent across the three spot size maps. The dorsal locus overlaid the V1/V2 border (red dotted lines) and another locus was slightly ventral to the HM in V1 (magenta dotted lines). Across the 0.4°, 0.6° and 0.8° spot sizes, the locations were similar (red dotted circles in overlay) and consistent with the overall topographic (HM and VM) organization.
V2/V3 (third column)There were also two loci for V2/V3. The dorsal activation fell within the banks and depths of the lunate sulcus (LUS, yellow dotted lines) consistent with previous studies23,24, had a distributed patchy appearance and was topographically consistent with a foveolar location along the V2/V3 border (yellow dotted circle in overlay). A second foveolar locus (evident for the 0.6° and 0.8° spots but below the threshold for the 0.4° spot) fell in the inferior occipital sulcus (IOS, orange dotted lines; orange dotted circle in overlay).
V3/V4 (second column)Two distinct foci were seen at the V3/V4 border, one on the anterior bank of the LUS (cyan dotted lines) and one at the tip of the IOS (green dotted lines). There were also a few patchy activations extending onto the cortical convexity (between the ends of the LUS and IOS and posterior to the superior temporal sulcus (STS)). The locations of these loci fell at the end of the V3/V4 border representing the VM (cyan dotted circle in overlay).
V4/TEO (second column) and TEO/FST (first column)The TEO is a midtier area and likely to exhibit somewhat weaker responses to simple spots25. Despite this, a locus of patchy activation is visible at the tip of the IOS (white dotted lines, V4/TEO border) and at the posterior border of the STS (blue dotted lines, consistent with activation at the TEO/FST border16).
Bilateral representationBecause the fovea is known to be represented in both hemispheres, we also examined the foveolar representation in the contralateral hemisphere. The radiofrequency (RF) coil was typically placed slightly off-center biased toward one hemisphere, such that we could capture as much of the ventral pathway as possible. The SNR on the contralateral hemisphere was, therefore, slightly lower but substantial blood-oxygenation-level-dependent (BOLD) signals could still be captured. In addition to the activation shown above (Fig. 3b, same as that shown in Fig. 3a, middle row), foveolar activations in the contralateral hemisphere, obtained from the same sessions, are shown in Fig. 3c (slice views in Extended Data Fig. 2). Each of the foveolar locations identified in Fig. 3a are indicated by circles of the corresponding color. Activation in TEO on the right side was weak. Similar data for monkey J are shown in Extended Data Fig. 1 and summarized in Fig. 3d,e.
In sum, these methods revealed precise foveolar activation locations. Foveolar activation often comprises one to a few distinct patches. The locations of these activations were stable across multiple sizes of spot stimuli and were consistent across multiple thresholds. In addition, we show that these activations were obtained with both better and poorer eye fixation behavior (Extended Data Fig. 3), indicating that the results were not an artifact of eye movements and that they were not because of edges of the spot stimuli or subtraction of the fixation cross (Extended Data Fig. 4). These data suggest that, in contrast to what was previously viewed as a singularity, there are at least eight distinct nodes of foveolar representation, one near the end of each VM and HM for dorsal and ventral V1/V2, V2/V3, V3/V4, V4/TEO and perhaps TEO/FST. This mapping is simultaneously observed in both hemispheres. Thus, the foveolar representation is not a single point or confluence and is more than one per visual area, as previously suggested. Moreover, these findings suggest that there is cortical territory bounded by these foveolar representations that is not within the visuotopically mapped regions (Extended Data Fig. 4). We provide a schematic ring (dotted lavender ring in Fig. 3b–e) around this central region and refer to it as the foveolar core (or core). This ring is not meant to be a specific border but an indication of the region of cortex bounded by the foveolar loci.
Phase encoding reveals a nonhomogeneous region and distinct foveolar core responsesAs phase encoding is the standard method for mapping retinotopy, we also conducted phase encoding for the central 3° in a fixating monkey (monkey J). Schira and colleagues15 studied the foveolar region in humans with phase encoding using very fine stimuli, high-resolution functional imaging (1.2-mm isotropic resolution) and sophisticated analysis. We attempted a similar study using a continuous sequence of very fine isoeccentric thin (0.15°) rings and narrow (5°) isopolar wedges. The resulting phase maps are shown in Fig. 4a,b (isoeccentricity and isopolarity, respectively); the foveolar core is indicated by a dashed lavender ring and foveolar loci are denoted by white circles. Consistent with phase-encoding studies10,14,15,16,26, in the isoeccentricity map, there was a systematic phase shift in V1, V2 and V3 on the lateral operculum, with the centermost (least phase-shifted) voxels (red) appearing most lateral on the cortex. This is further illustrated by the shifting time courses across regions of interest (ROIs) 1–6 along the V1/V2 border (Fig. 4d, left column; graphs 1–6, shifting red arrowheads); the regularity of the time courses and small degree of variability in these traces indicate that eye fixation was excellent.
Fig. 4: Phase encoding using fine isoeccentricity and isopolarity stimuli in visuotopic V1–V4 and in foveolar core.
a, Isoeccentricity map. Inset, isoeccentricity stimulus: continuous increasing size of isoeccentric rings (each ring: 0.15° wide; one trial consisted of 50 rings over 3°, 0.06° per ring, lasting 50 s with a 10-s ISI; fixation point: 0.05° white dot, constant over a total of 21 trials). b, Isopolarity map. Inset, isopolarity stimulus: continuous rotating checkerboard wedge (5° wedge, rotated over 50 s from 90° to −90° in the contralateral visual field; size: 3°, over a total of 21 trials). Black circles in a and b denote the locations of voxel time courses shown in d,e. White circles in a and b denote the foveolar loci identified in Fig. 3d,e. c, Histogram showing that FFT amplitude at the stimulus frequency is weaker in ‘inside core’ compared to ‘outside core’ samples. FFT amplitude: inside core, mean = 1.2; outside core, mean = 3.0; P < 0.0001 (two-sided chi-square test). d, Isoeccentricity time courses (BOLD%). ROIs 1–6 are in V1 along the VM. Red arrowheads (center of mass of peak) indicate a peak shift as ROIs approach the center. ROIs 7–15 are in the foveolar core along an anteroposterior axis through the core. ROIs 16 and 17 are along a posterosuperior axis. ROIs 18 and 19 are along a posteroinferior axis. ROIs 20–22 are along an anteroinferior axis. e, Isopolarity time courses (BOLD%). ROIs 1–9 are along three isoeccentricity rings in V1. ROIs 10–12 are in foveolar core parallel to isoeccentricity rings in V1. Note: because the dorsal cortex represents ventral fields, the phase shift because of the wedge approaching the VM in the ventral field results in a shortening latency to response (red arrowheads). Red boxes represent time courses from ROIs in the core region. Numbers in the right corner of each graph denote FFT amplitude indices (Extended Data Fig. 6). All data in d,e are presented as mean values ± s.e.m.
We hypothesized that, if the foveolar center were within the core, we would observe continuously shifting phase from the several foveolar loci toward the center. We, therefore, sampled phase response at locations through the most central (red) regions of the foveolar core starting from (1) the dorsal V1/V2 foveolar locus (ROIs 7–15); (2) the dorsal V2/V3 to the ventral V2/V3 foveolar loci (ROIs 16–19); and (3) the ventral V3/V4 foveolar locus (ROI 20–22). Anterior V4/TEO locations were outside our imaging field of view (FOV) in this experiment. As shown in Fig. 4d (three rightmost columns), we found that these samples exhibited surprisingly noisy responses that were poorly phase-locked to the stimulus.
In the isopolarity map (Fig. 4b), outside the foveolar ring, the ROIs were well modulated and showed phase shifting (Fig. 4e, top three columns, ROIs 1–3, 4–6 and 7–9 along central isopolarities, shifting red arrowheads). Again, inside the core region, sampled from a parallel arc just inside the border of the core region (Fig. 4e, ROIs 10–12) and in the central zone of the core region (Fig. 4e, ROIs 13–15), the traces were noisy and not well modulated. This was supported by significant differences in the peak amplitude of the response frequency of ‘within core’ and ‘outside core’ samples (Fig. 4c and Extended Data Fig. 5).
Thus, whereas our methods could detect phase shifting outside the core, the responses in the core appeared noisy and poorly phase-locked. If the noisy signals were because of eye movements, the foveolar traces from outside core samples would also be noisy, inconsistent with the stable traces obtained. It may be possible that neurons in the core prefer even smaller stimuli (<0.15°); however, the within core traces did exhibit responsiveness to the stimuli (large magnitude fluctuations) but had poor phase correlation. While the nature of the noisy traces remains to be examined, these data suggest that responses in the core region are qualitatively distinct from those in retinotopic regions of visual cortex. We suggest that phase-encoding maps obtained in humans also show similar organization (Extended Data Fig. 6).
Are there functional domains within the foveolar cortex?One of the defining features of visual cortical areas is their functional organization for different parameters of visual information. Well known are the stripes of V2 that range in size from 1 to 2 mm in width and the distinct sets of feature specific submillimeter-sized domains within the thin, pale and thick stripes21,23,27. While, imaging millimeter-scale or submillimeter-scale modular structures with fMRI in monkeys has been challenging17,23, we illustrate here that V2 color (thin) stripes were well mapped by color versus achromatic gratings but failed to evoke much activation in the foveolar core (Extended Data Fig. 7). We then proceeded to explore the foveolar core response using a small battery of visual stimuli, including high and low SFs, simple color and achromatic stimuli and motion dot stimuli.
High-SF domainsA defining feature of foveolar function is its high visual acuity, marked by neuronal responses to high SFs visible only to foveolar vision28. Neurons within foveolar locations of macaque V1 have extremely small receptive field sizes (predominantly 10–20 minutes of arc in size29) and exhibit peak SF tunings up to ~5–8 cycles per degree30. Peak SF tunings based on fMRI BOLD signals reach ~3 cycles per degree31. However, perceptual detection and discrimination can reach SFs well over 10 cycles per degree in humans28. Thus, a potential function of the core region may be to integrate lower-order SFs to achieve high spatial acuity28,32. In this section, we use the term ‘central’ to mean foveolar, not in the topographic sense but in the functional sense of central visual perception.
To capture central SF responses, we examined response to high-SF achromatic gratings (11, 15 and 18 cycles per degree) and, for comparison, to low-SF achromatic gratings (0.2 cycles per degree) and to small flashing color spots. Monkeys maintained excellent fixation behavior during presentation of these stimuli. As shown in Fig. 5a, activations to SF11 (yellow-green), SF15 (purple) and SF18 (orange) gratings revealed patchy activations scattered throughout the core (dashed oval). Moreover, SF11, SF15 and SF18 showed little overlap, suggesting that they are distinct functional domains (Fig. 5a,g; overlap index for all comparisons < 0.05). In sum, these high-SF stimuli appear to effectively activate the region within the core region and exhibit distinct spatial activations.
Fig. 5: Mesoscale functional domains within the foveolar core.
Color-coded responses to different visual stimuli. a, Activations to achromatic high-SF gratings (in cycles per degree). Yellow, SF11; purple, SF15; orange, SF18. b, Overlay of high-SF (yellow, SF11 + SF15 + SF18) and low-SF (red, SF0.2) activations. c, Overlay of low-SF (red, SF0.2), motion (cyan, clockwise moving dots) and color (blue, 0.8° flashing spot) activations. d, Overlay of all domains. e, Four fields of view from d showing multiple types of functional domains within a small region. f, Averaged time course of each domain type selected from 3–5 clusters within the core. All data are presented as the mean values ± s.e.m. The numbers of voxels from top to bottom are 588, 420, 387 and 385, respectively. g, Overlay indices between different populations of domains show little overlap between domains (overlap indices: 0.09, range 0–0.25). h, Domain size distribution (total number: 111; Methods and Extended Data Fig. 8).
We then compared these maps to those obtained with a low-SF (0.2 cycles per degree) achromatic grating, an SF typically used for studying surface properties such as color and luminance. The SF0.2 responsive domains (Fig. 5b, red) were largely nonoverlapping with the high-SF domains (Fig. 5b, yellow: sum of SF11, SF15 and SF18; overlap indices < 0.1). Achromatic SF0.2 domains (Fig. 5c, red) also had low overlap with domains activated by a small (0.8°) color stimulus (Fig. 5c, blue; overlap index = 0.064). For comparison, response to small motion dot stimuli (2° patch of dots rotating clockwise) (Fig. 5c, cyan) elicited activations with low overlap with SF0.2 achromatic (red; overlap index = 0.25) and slightly lower overlap with color domains (blue; overlap index = 0.17). Domains of different types appeared within small regions within close proximity to one another and did not appear to coalesce into stripes or bands (examples in Fig. 5e). Averaged time courses of different domains within the foveolar core exhibited reasonable BOLD time courses (Fig. 5f), indicating that these activations were not artifactual.
Domain sizesWe also estimated the sizes of the activation domains. The area of each activation was calculated by measuring the number of nodes within the activation when overlaid onto the brain surface mesh33 (Extended Data Fig. 8). Given the 0.6-mm in-plane resolution, we conservatively classified domains as <1 mm, 1–2 mm and >2 mm in size. Domain diameters (the longest axis) revealed that they were largely <2 mm in size (Fig. 5h, top; 82% ≤2 mm in diameter) and held for different functional domain types (Fig. 5h, bottom).
Thus, remarkably, what originally appeared as a region of cortex with little response to foveolar spot stimuli was actually a region densely populated with spatially distinct functional ‘domains’ of different feature selectivity (domain overlay in Fig. 5d; overlap indices < 0.15). Unlike the stripe and band organizations in early visual areas, each of the domain types appeared to be distributed broadly across this central zone, resulting in a mosaic-like functional architecture.
A view from optical imagingTo further examine the topographic and functional organization of the foveolar core, we conducted intrinsic signal optical imaging of this area in anesthetized monkeys. To functionally localize the foveal representation in anesthetized monkeys, determination of the foveolar location was determined by sequential, iterative imaging of large-to-small-interval vertical and horizontal bar stimuli to zero in on the precise x and y coordinates on the monitor (Methods). As shown in Fig. 6e, the lateral visual cortex was exposed, including lateral parts (central 0.5°) of V1, V2 and V4 and part of the TEO. Maps for ocular dominance revealed the V1/V2 border (Fig. 6a, fine dotted line). Mapping orientation preference (for example, horizontal versus vertical) revealed orientation maps in V1, orientation stripes in V2 (corresponding to the thick (‘disparity’) and pale (‘higher-order orientation’) stripes; regions between the yellow arrows) and larger-orientation (‘curvature’) responsive bands in V4 (Fig. 6b, outlined in green). Color versus achromatic response revealed blobs (‘color’) in V1 (pattern of dark dots), thin (‘hue and brightness’) stripes in V2 (yellow arrows) and larger bands of ‘contextual color and brightness’ response in V4 (outlined in red) (Fig. 6c). The color-responsive stripes in V2 aligned well with dark cytochrome oxidase histology (Fig. 6d, three stripes indicated by yellow arrows). Each area (V1, V2 and V4) was characterized by distinct and well-documented functional organizations34,35,36. These findings establish the quality of our optical imaging methods.
Fig. 6: Multiple foveolar representations revealed by optical imaging.
a, Ocular dominance (OD) map. Scale bar, 1 mm. b, Orientation map (45° versus 135°). Green outlines, V4 orientation bands; red outlines, V4 color bands. c, Color map (color versus achromatic). Yellow arrows, color stripes in V2 (same locations as stripes shown in d). d, Cytochrome oxidase stained stripes in V2 align well with color stripes in c (yellow arrows). e, Blood vessel map over V1, V2, V4 and TEO. M, medial; L, lateral; A, anterior; P, posterior. f, Color-coded eccentricity map (red, 0.01°; yellow, 0.10°; green, 0.20°; cyan, 0.30°; blue, 0.40°; purple, 0.50°). Lavender dashed circle, foveolar core; colored arrows, shifted eccentricities. White arrow: V1/V2 border. g, Color-coded isopolarity map (red, 0°; yellow, 15°; light green, 30°; dark green, 45°; cyan, 60°; blue, 75°; purple, 90°). Numbered white dotted circles, foveolar locations at V1/V2 (1 and 2), V2v/V3v (4), V3v/V4v (6), V4v/TEO (7) and TEO/FST (8). Locations V2d/V3d (3) and V3d/V4d (5) are within the LUS and not visible in optical imaging. Colored arrows, shifted polar angles. h, For comparison, 7T MRI of all foveola in monkey E (left hemisphere, from Fig. 3b) numbered with the corresponding locations in g. The orientation of all optical imaging maps was rotated to correspond with the MRI results shown in h.
Central 0.5° visuotopyTo determine foveolar representations, we conducted visuotopic mapping using thin (two pixels wide) (1) isoeccentricity arcs at 0.01°, 0.1°, 0.2°, 0.3°, 0.4° and 0.5° eccentricities (Fig. 6f, inset; red, yellow, green, cyan, blue and purple, respectively) and (2) isopolarity lines, from VM to HM at 90°, 75°, 60°, 45°, 30°, 15° and 0° (Fig. 6g, inset; red, yellow, light green, dark green, cyan, blue and purple, respectively). These generated vector maps (the color of each pixel reflects the preference for one of the respective isoeccentric and isopolar conditions; single condition maps in Extended Data Fig. 9). In V1, isoeccentricity of the central 0.5° mapped mediolaterally (Fig. 6f, from purple to blue, green, yellow and red) and isopolarity mapped roughly posterior to anterior (Fig. 6g, from purple to blue, green, yellow and red). V2 (which only had a 2-mm-wide region visible on surface posterior to LUS) exhibited a similar mediolateral mapping of isoeccentricity (Fig. 6f) and shared the VM representation at the V1/V2 border (Fig. 6g, white arrow). The yellow dashed circles in Fig. 6f indicate corresponding locations of 0.1° eccentricity in V1, V2 and V4. Note that, in the isopolarity color code, the foveal center responds to all isopolar conditions; this led to no preference for any single condition and, therefore, resulted in black colored pixels. Thus, the corresponding foveolar representations (foveolar loci 1 (V1/V2d) and 2 (V1/V2v)) appeared red in the isoeccentricity map and black in the isopolarity map.
In V4, where receptive fields were larger than in V1 and V2, the map was weaker in response to the fine isoeccentricity and isopolarity stimuli but still consistent with known V4 topography. The isoeccentricity topography ran mediolaterally (Fig. 6f, colored arrows; the most medial regions were purple, blue and green, with yellow pixels further lateral). Even more lateral was a large dark region (likely indicating poor response to the central 0.01° stimulus) consistent with the foveal V4 location (dashed red arrow). In the isopolarity map, V4 exhibited weak responses but an anteroposterior isopolar topography could be discerned (in Fig. 6g, colored arrows; from purple to blue, green and red). Within TEO, the topography was less clear and also weak to central stimuli.
Foveolar centers and foveolar coreTwo foveal centers at the V1/V2 border (1 (V1/V2d) and 2 (V1/V2v)) were seen as red in Fig. 6f and black in Fig. 6g. Foveolar loci 4 (V2/V3v, at dorsal lip of IOS), 6 (V3/V4v, more anterior on dorsal lip of IOS) and 7 (V4–TEOv, anterior on ventral lip of IOS) also appeared as red in Fig. 6f and black in Fig. 6g. Foveolar locus 8 (V4/TEOd) was visible on the surface and could extend into the STS (fewer red pixels in Fig. 6f because of weak response to tiny foveal stimuli). Foveolar loci 3 (V2/V3d) and 5 (V3/V4d) were not visible as they were within the LUS (Fig. 6h). The locations of foveolar loci 6, 7 and 8 were consistent with the corresponding loci shown in fMRI (Fig. 6h). In the core region (approximated by the lavender ring), there were intermingled patches of yellow, green, blue, red and purple, as well as dark regions.
In sum, the primary finding is that the optical imaging data were largely consistent with the fMRI findings. First, there were multiple foveolar loci (dark circled loci in Fig. 6g). Secondly, the visuotopic maps, most clear in V1 and V4, did not continue into the core region. Thirdly, there were patchy intermingled activations in the core region. Although these fMRI data are from different animals, these foveal centers provide support for those identified using fMRI (corresponding numbers in Fig. 6h). Following this reasoning, the lavender dashed oval region in the optical image in Fig. 6f,g indicates the approximate location of the foveolar core. Because of the folding of the cortex, the region corresponding to the foveolar core in the optical image may appear smaller than in fMRI. Thus, with a completely different imaging methodology and different spatial resolution, similar results were obtained, further strengthening these findings.
Foveolar cortical magnificationThis finding introduces new questions regarding how much cortex is devoted to foveolar representation. The CMF, defined as the millimeters of cortical area devoted to processing a degree of visual angle, for each area has been well studied
留言 (0)