
記住我


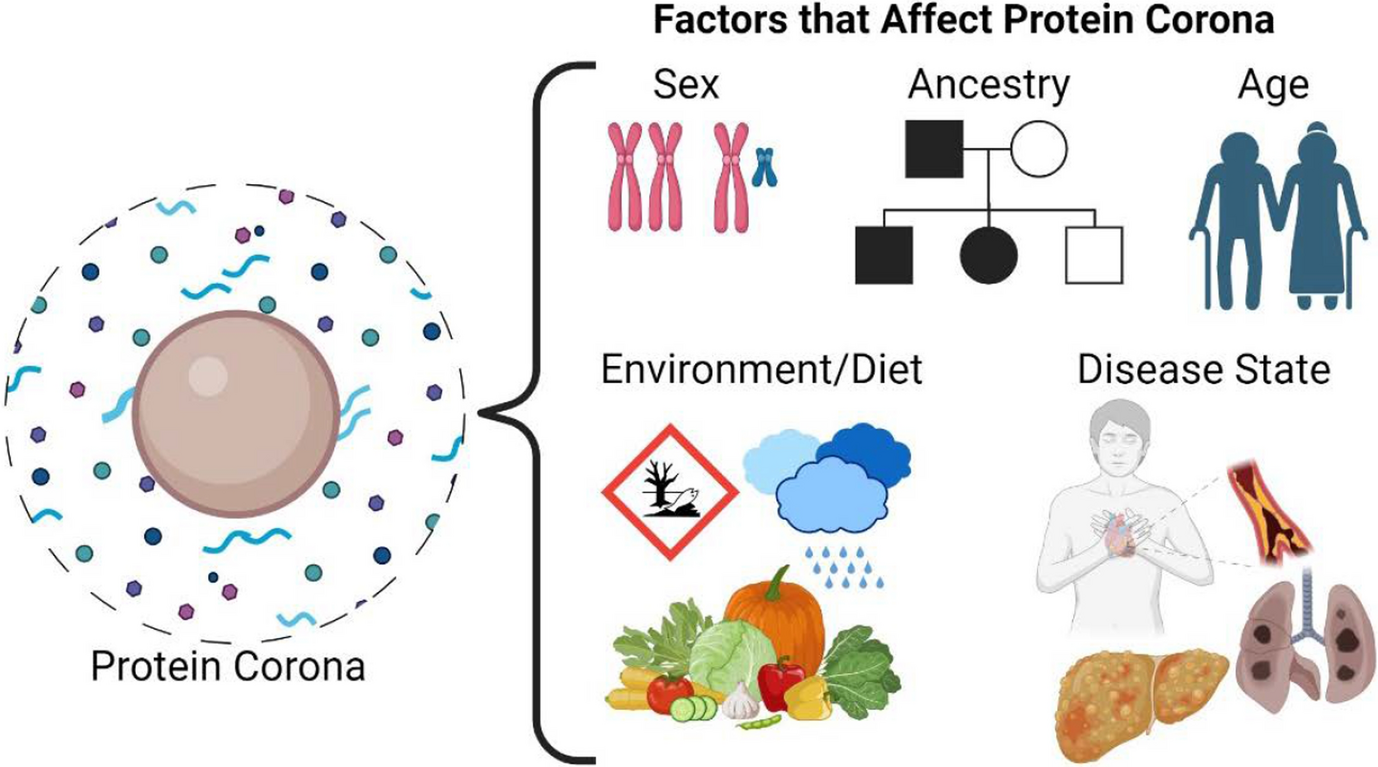




Sex and gender are often used synonymously; however, it is important to make distinctions between these two terms as they are both known to contribute to health from etiology to outcomes [15, 16]. On the most basic level, sex is a biological factor related to genetics, physiology, and anatomy and can be attributed to differences that arise on cellular and molecular levels [5]. Biological differences can include the presence/absence of the Y chromosome, bone shape and density, metabolic regulation mechanisms, and presence/absence of reproductive organs. Gender is a social construct and refers to an individual’s identity, expression, and presentation as man, woman, or non-binary [17]. Gender is shaped by sociocultural factors such as prescribed gender roles, power dynamics, and relationships. The interaction of gender and medicine can be seen through help-seeking behavior, disease perception, decision making, and use of health care [18]. Conflation of sex and gender, both inadvertently and intentionally, is more harmful than neutral to health research. Such aggregation leads to incomplete analysis and can lead to adverse health outcomes and gaps in innovative medical treatment and policy [19, 20]. While there are arguable differences between sex and gender, it is important to note that they are both fluid. Sex is on a spectrum and can be culturally influenced as well. Variations in chromosomes, hormone levels, and reproductive organs can result in more than two sexes [17]. Similarly, the concept of gender varies as a result of institutional and cultural change [17].
Nanotechnology and sexSex-based differences arise on a spectrum from variables including differences between the X and Y chromosome, X-inactivation, sex steroid hormones, reproductive organs, fat distributions, and more. In the past, these differences have led to different drug metabolism rates, and lack of inclusion in drug trials led to higher hospitalization rates and adverse effects in women [21]. Investigational New Drug Applications often neglect sex as a variable, but the ones that include it have shown up to 40% differences in pharmacokinetics [22]. These differences include properties such as drug absorption, metabolism, distribution, and excretion.
These differences can also affect the performance of nanotechnology. Researchers have begun to investigate sex-based differences which has revealed variability between sex in the biodistribution of NPs in different tissues. For example, females have greater accumulation of NPs in the kidneys as compared to males. Conversely, males have greater accumulation of NPs in the liver as compared to females [5]. Besides biodistribution differences, therapeutic effects between sexes can differ as well. An inflammation model in mice using macrophage-targeted nanotheranostics revealed sex-based differences in COX-2 inhibition, an enzyme driving inflammation, indicating greater therapeutic success in males versus females [23]. Furthermore, in an in utero study, only hypoxic female mice responded positively to MitoQ NP treatment where pro-angiogenic factor Vegfa and growth factor Igf2 expression increased [24]. Possible explanations for differences in biodistribution and therapeutic efficacy can be delineated through natural physiological processes. For instance, the menstrual cycle effects have also been shown to affect both biodistribution of NPs and therapeutic efficacy for applications in ovarian and breast cancer in a study by Poley et al. They found that gadolinium NP accumulation was the highest in the ovaries and uterus during ovulation. Based on this finding, they delivered therapeutic PEGylated liposomal doxorubicin NPs, it was found that efficacy was highest during ovulation when treating ovarian cancer. Simultaneously, when mice were treated for breast cancer during the same period, NPs continued to accumulate in the reproductive system which led to decreased efficacy in treatment [25]. Differences in biodistributions seen over the menstrual cycle in this study can be explained from an anatomical perspective; increased vascularity at different stages in the menstrual cycle can lead to differential drug trafficking causing some organs to accumulate therapeutics. Glaringly, males do not have this physiology. Therefore, it is important that NPs designed for females be modified to account for these differences. To further improve NP formulations, investigating protein coronas could provide a starting point into how performance of NPs differ with respect to biological sex.
Protein corona and sexSex differences in serums are well-documented in medical literature. These differences include statistically significant variability in concentrations as well as presence/absence of proteins and other analytes in metabolic activity [26,27,28,29,30,31]. As a result of sex-linked differences in serum compositions, sex has been implied as a critical factor that affects NP performance with respect to sex-specific protein coronas [5, 32, 33]. Sex-specific protein coronas can affect the performance of an NP since differential proteins are recruited which can affect its size, surface charge, shape, and chemistry. The modification of an NPs physicochemical characteristics, in turn, affects the NP-cell interactions [34].
Sex-based differences in protein corona formation in non-human speciesMost research in investigating sex-based differences in protein corona has been more exclusive to non-human species. In a pioneering study by Hayashi et al., 70 nm SiO2 NPs were used to test whether biological identity from protein corona affected uptake into immune cells from zebrafish. In all the serums tested, apolipoprotein A-I was detected in the hard protein corona of the NPs validating the plausibility of this study [35]. Vitellogenin and fetuin were found in abundance in NPs soaked in female and male plasma, respectively, revealing sex-specific biological identity in the protein corona. Vitellogenin is an egg precursor protein present in zebrafish oviparous reproductive systems which accounts for the difference in protein aggregation onto the NP surface. When testing NPs pre-soaked in either male or female serum, it was found that those soaked in female serum were preferentially uptaken into both myeloid and lymphoid cells isolated from whole kidney marrow cells. However, it should be noted here that the authors did not report the sex of the cells which were used for these studies, and this could be a relevant factor as well. Sexual dimorphism in cells should be a variable that is tested or controlled for since there is well-documented sex-dependent gene expression and differential cellular responses due to hormonal exposure [36,37,38,39,40]. While the biological mechanism in preferential uptake requires further investigation, the authors offer a suggestion that proteins on the surface of the NP prompt innate immune responses since they may have aided in NP recognition [35].
Based on this study, an extension of this study by Gao et al. showed similar results overall as Hayashi et al. in smallmouth bass using polyvinylpyrrolidone-coated silver NPs (PVP-AgNPs) [6]. 50 nm PVP-AgNPs were soaked in female and male plasma and showed an increase in size by about 8 nm. Zeta potential measurements showed greater instability and changes in PVP-AgNPs in male plasma versus female plasma. Furthermore, less than 40% of the proteins in the protein corona were common between sex. Here, both zona pellucida (ZP) and vitellogenin, proteins taken up in developing follicles, accumulated in the protein corona of PVP-AgNPs. With vitellogenin and ZP accumulating preferentially, the combination with PVP-AgNPs can cause localization to the ovaries in fish. This study also showed that proteins commonly associated with the immune system, notably immunoglobulins and complement proteins, accumulated more abundantly on the surface of PVP-AgNPs [6]. Previous work using PVP-AgNPs in fish have shown adverse effects in offspring and follicular development. Based on this work, NPs could be more toxic to female fish due to potential accumulation in the ovaries, an organ that is absent in males. Further experiments in delineating NP organ accumulation would definitely be an interesting extension of this study. Another interesting extension could be extracting blood from specific organs, identifying proteins specific to each sex, and testing their effect on NP accumulation in the organ. Figure 3 shows the amount of shared protein in the corona when PVP-AgNPs were incubated in male or female for 1 h or 24 h.
Fig. 3
(Copyright 2017 American Chemical Society)
Proportion of shared proteins in protein coronas between male (M) and female (F) fish and incubation time (1 h or 24 h) along with a summary of key findings from Gao et al. [6]. Reprinted with permission from Gao et al. [6]
In a study by Ashkarran et al., silica NPs (SiNPs) of varying particle size and porosity were soaked in mice plasma to test sex-based differences in protein corona recruitment [41]. Initial characterization of the NPs showed that the diameter of the all NPs increased by 8–35 nm and zeta potential slightly decreased (more positive) indicating slightly lower colloidal stability. In mesoporous SiNPs, NPs containing numerous small pores, at approximately 100 nm size, 17 proteins were found to be significantly greater in SiNPs soaked in male plasma versus female plasma. Resistin, a protein involved in glucose uptake suppression and inflammation, was the protein found to be significantly higher in male protein corona. Other identified significantly different proteins include complement factors C8a, C8b, and C8g which are proteins responsible for innate and adaptive immunity. In Stöber SiNPs (non-mesoporous) at 100 nm size, male urinary protein 1, a protein binding pheromones from male urine, was found to be significantly greater in SiNPs soaked in male plasma versus female plasma validating the approach of this study. Here, proteins related to inflammation and tumorigenicity were found to be significantly greater in male soaked SiNPs than female soaked SiNPs including Krt16, Ig-heavy chain V region MOPC 173, and carboxylesterase ID. Similar results were observed in Stöber SiNPs (non-mesoporous) at 50 nm size where Rab27b, Ig-heavy chain V region 5–84, H4c1, and Ig-heavy chain V region MOPC 173 were all found to be significantly in male soaked SiNPs than female soaked SiNPs. Table 1 shows a summary of proteins found at significant levels in the protein corona of NPs with varying porosity and size.
Table 1 Proteins measured in protein coronas at significant difference based on sex for each NP type from Ashkarran et al. [41].Based on the differential proteins adsorbed onto the mesoporous and the Stober 100 nm and 50 nm particles, it was shown that size and porosity play a role in protein corona formation [41]. Notably, in the mesoporous SiNPs’ protein corona, there were significantly more unique proteins versus the non-mesoporous SiNPs. Porosity increases the amount of surface area relative to volume allowing for increased protein absorption onto the NP. An extension of this work on size could be the addition of a polymer to “standardize” protein corona recruitment to measure the same identity and similar concentrations of protein between male and female plasma. Additionally, protein coronas formed from male plasma showed significantly greater protein recruitment than female plasma implying that biosensing capacity was greater for the males versus females. An interesting extension of this work could be to use the SiNPs to test protein recruitment for mice that have a known disease. This could reveal and quantify diagnostic efficacy for nanotechnologies as well as clarify any sex-based differences in clinical biomarkers that may arise in practical use. Table 2 shows a summary of sex-based differences in NP parameters in different species.
Table 2 Summary of parameters from literature using non-human species for sex-based difference studiesWhile there has been progress in the field of sex-based nanomedicine in non-human species, it would be interesting to see experiments using primates to identify proteins more applicable to humans. Testing nanotechnologies in harvested blood/tissue samples from specific primate organs, specifically the GI tract, liver, and kidneys, could provide valuable insight into what proteins promote clearance, uptake, cytotoxicity, and immune responses in each of these organs so that scientists can improve formulations to yield better clinical trial results.
Sex-based differences in protein corona formation in humansThere is limited research discussing biological and molecular mechanisms of sex-based differences in humans, as shown in Table 3. However, a notable study by Vulpis et al. revealed sex may be an important factor in nanomedicine [42]. In their study, DOTAP lipid NP uptake was tested in six distinct immune cells differentiated by sex. It was found that uptake in natural killer (NK) cells, lymphocyte cells critical in innate immunity and immune response regulation, was significantly greater in male cells. These results suggest that the difference in uptake between the male and female NK cells lead to disparate immune system performance. The researchers then repeated the uptake studies by pre-treating the NPs with human plasma to test the effects of protein corona. By pre-treating with human plasma, cell-NP interactions are able to be controlled better. The pre-treatment of the NPs eliminated the sex differences shown in NK cell uptake and dramatically reduced uptake in all immune cells tested. Hence, these results suggest that pre-forming a protein corona can help to mitigate and standardize immune responses in patients regardless of sex [42]. This study introduces a unique strategy to eliminate sex-based differences in nanomedicine; a standard “cocktail” of human plasma could be developed to attenuate the effects of sex in nanomedicine. To expand upon this study, researchers could consider how hormones and other sexual dimorphisms could affect uptake and immune response.
However, a challenge faced by researchers attempting to evaluate biological sex in nanomedicine is interference from hormonal variations. In a study by Serpooshan et al., human amniotic stem cells (hAMSCs) were used to circumvent this issue, as hAMSCs represent one of the earliest stages of somatic stem cells since they have sexual dimorphism without the initiation of hormonal changes [34]. Hence, the differences in NP performance should relate to genetic and structural dimorphisms. In their study, cytokine release in hAMSCs was compared before treatment with quantum dots. Prior to treatment, it was found that 14 cytokines were found to be at significantly higher levels in male hAMSCs versus their female counterparts. With respect to cytokines, NP-cell behavior could be altered with respect to immune response; the recruitment of certain cytokines could induce an immune response or trigger pathways in one sex versus the another.
Sex-specific hAMSCs and primary fibroblasts were then treated with quantum dots (QDs) controlled for size and surface chemistry. Flow cytometry results revealed greater accumulation of QDs in female hAMSCs as opposed to male hAMSCs. To partially explain this difference, QDs were incubated with serum and the zeta potential was measured. The initial QDs’ surface charge was − 22 ± 1 mV. After exposure to serum, the QDs’ surface charges were − 15 ± 1 and − 11 ± 1 mV for female and male serum, respectively. A more negative zeta potential indicates greater colloidal stability which partially explains why there was higher uptake into female cells. This finding provides empirical evidence into how the physicochemical characteristics of the protein corona can affect NP-cell behavior; here we see how the protein corona profile can affect uptake of quantum dots [34].
The cytoskeletons were then analyzed to examine structural differences to further explain differences in quantum dot uptake. Via the use of stochastic optical reconstruction microscopy, actin filaments were found to be organized differentially between sex-specific hAMSCs implying that there could be differences in uptake and trafficking of QDs. The results indicate that filaments are arranged differentially with respect to bundles, density, and direction. After treatment with quantum dots, it was found that quantum dots localized to actin filaments, structures which help to facilitate endocytosis. Sex differences were then tested in the uptake of quantum dots in clathrin-coated vesicles, proteins that coat vesicles that are endocytosed via the plasma membrane to aid in endocytosis. From this experiment, the results suggested male hAMSCs accumulated more clathrin than female hAMSCs. To test whether cell type affects QD uptake in sex-specific cells, the experiments were repeated in primary fibroblasts from a male and female patient of similar age which showed that male fibroblasts had greater uptake instead concluding that sex-based differences were present. The differences in QD uptake and clathrin-coated vesicles in different cell types largely boiled down to the fact that the male fibroblasts had a greater amount of actin filaments allowing for increased uptake into cells [34].
However, this did not hold true when the fibroblasts were treated with gold NPs (AuNPs). The results suggest that AuNP uptake is non-specific meaning that uptake can be mediated by endocytosis or direct penetration through the plasma membrane. Unlike the QDs, the NPs were not localized to the actin filaments. Instead, they were more diffusive throughout the cytosol [34].
This work highlights many factors that can affect nanotechnological efficacy as a result of sex including protein secretion, cytoskeletal differences, and physicochemical characteristics. This work could be improved by incorporating more variables known to affect health outcomes including geographical/genetic ancestry and age. For instance, the cytoskeleton can undergo remodeling during natural aging which can affect the cell cycle, polarity, and cell migration [43]. This work could also benefit from including size data for the QD experiments to support sex-differences in uptake claims. The size of the NPs/QDs are known to affect protein corona recruitment which, in turn, affects uptake into cells [44].
Table 3 Summary of parameters from literature using human species for sex-based difference studiesConsidering biological sex in future nanomedicinesThe field of sex-based nanomedicine in humans is in its infancy; there has not been nearly enough progress in this field. More in vitro and ex vivo experiments need to be conducted to test sex as a biological variable. Echoing a previous statement, harvesting samples from a specific organ could provide insight into what proteins and biomolecules promote accumulation, immune response, and cytotoxicity. Future studies testing biological sex as a variable should also consider de-coupling sex and gender which can be done by testing embryonic cells for sex differences. An interesting way that confounding variables have been controlled for is by generating isogenic induced pluripotent stem cells from a Klinefelter syndrome patient. iPSCs containing XX and XY cells were made from the XXY parent cells to isogenically study sex differences [45]. Theoretically, these cells should contain identical genetic makeup from ancestry to disease state other than biological sex.
The effects of genetic ancestry on protein coronaDifference between race and ancestryRace and ancestry are often convoluted much like sex and gender. However, it is important to delineate between the two since both are known to affect health outcomes in distinct ways. Race and ethnicity are social constructs based on societal perception and biases and were a means to justify slavery, segregation, and socioeconomic inequities [46, 47]. Notably, race and ethnicity have been shown to have no biological basis [48]. However, it is irrefutable that structural and institutional racism continues to persist as a factor in determinants of health [48]. For example, Black and Hispanic populations experienced significantly higher prevalence, hospitalization, and mortality rates due to COVID-19 than White people underscoring the need to address and improve health inequity [49].
Ancestry refers to one’s family origin and heritage [46, 47]. Furthermore, ancestry focuses on one’s biological/genetic background and geographic origin rather than skin color in order to identify relevant markers that show significant differences in frequencies between populations [47]. Therefore, ancestry is a more inclusive and clinically relevant factor that can be used to improve accuracy and efficacy of medicines. Research suggests variance in drug metabolism between different ancestral groups. For example, dosing for eltrombopag olamine was reduced by 50% for patients of East Asian descent due to a 15% incidence of hepatobiliary plasma abnormalities compared to 5% incidence in Caucasians [50]. However, it was undeterminable if genetic predisposition was the cause of this disparity. Investigation into ancestral/genetic determinants of health could help improve efficacy of medicines.
Nanotechnology, ancestry, and the protein coronaTo our knowledge, there is no research investigating ancestry as a factor in the performance of nanotechnologies nor the protein corona. Therefore, here, we describe variances in proteome analysis between different ancestral backgrounds which could influence the protein corona.
Sjaarda et al. evaluated the impact of ancestry on an admixture of 237 serum biomarker concentrations in diabetes patients via the use of variance component modeling [51]. Results indicated that ancestry affects 19% of biomarkers with 5% having significant phenotypic variance by more than 10%. The region at rs4149261 was associated with increased levels of C-peptide, and increased risk of diabetes and insulin resistance. This was found to be an effect of African ancestry; however, it was not specified what region of Africa was tested. Further hypothesis testing found that 5% of the biomarkers were affected by ancestry with 30% showing significant differences. The findings from the researchers’ models suggest that “biomarkers harbor true biological inter-ancestry differences in contraction that are genetically determined” [51]. With these differences in biomarker concentrations, one can expect differential protein corona absorption when administering a nanotechnologically-based therapeutic for diabetes treatment due to variances in C-peptide concentration and other biomarkers. This, in turn, can affect the performance and uptake of the therapeutic due to physicochemical modifications to the surface of the NP including size, zeta potential, etc.
Many studies have investigated protein quantitative trait loci (pQTL) to look at genetic regulation of proteome circulation and how it relates to disease susceptibility and predisposition. Xu et al. identified pQTL to identify any genetic control in proteome circulation in Han Chinese in addition to providing insights into disease susceptibility between groups of East Asian descent and European descent [52]. Results indicated that 45 out of 60 protein-phenotypes were not previously identified in Europeans. Out of the 41 pQTLs in both European and East Asian populations, genetically determined body mass index (BMI) was positively associated with 28 proteins and negatively associated with 2 proteins in European. Analysis in the East Asian population indicated 34 non-significant negative associations with BMI implying that the two populations may have varying genetic predisposition to obesity. Different proteins were also identified between East Asian and European populations that may suppress BMI effects on type II diabetes and coronary artery disease. Haptoglobin, alpha-1-antitrypsin, heparin factor 2, factor H, and C4b-binding protein alpha chain were suggested to suppress the effect of BMI on type II diabetes in European populations. Haptoglobin, heparin factor 2, factor H, inter-alpha-trypsin inhibitor heavy chain H3, and kininogen-1 were also suggested to suppress BMI effects on coronary artery disease in European populations. These proteins were not observed to have the same effect in the East Asian population. This cross-ancestry analysis revealed ancestral-based differences in proteomics which could point to disease susceptibility and prevalence between populations with different geographical origin [52]. In nanotechnology, disease-specific protein corona is known to form on the surface of NPs which can affect the uptake, performance, and efficacy of the therapeutic. Further discussion of disease state and the protein corona can be found in “The effects of disease state on protein corona” section.
Kachuri et al. took a similar approach in looking at pQTLs in African Americans, Puerto Ricans, and Mexican Americans [53]. They revealed greater genetic variance in whole-blood gene expression in African Americans followed by Europeans and Indigenous Americans. Additionally, they also found that 30% of heritable protein-coding genes are ancestry specific. These genes are very rare or do not exist in other ethnic populations, and this finding stays consistent when the defined ancestral boundaries were relaxed.
Zhang et al. developed models for plasma protein imputation for European and African populations to pinpoint drug targets for gout [54]. Their analysis revealed that 30% of the sentinel pQTL in the African populations was nonexistent or extremely rare in European populations. Conversely, 10% of the sentinel pQTL in the European populations was nonexistent or extremely rare. When applying these models cross-ancestry, it was found that applying African models on to the European populations performed better than the converse [54]. Again, this highlights the effect that geographical ancestry can have on plasma proteome composition due to the fact that the protein imputation models perform differently across ancestry. NPs that are designed to target a specific protein may not develop the appropriate protein corona in order to have their intended effects on different ancestral groups since proteomes may not be common. Hence, this work also highlights the need to develop specific models for different ancestral groups to further develop formulations for nanotechnologies.
Zhou et al. determined that increased levels of 2ʹ-5ʹ-Oligoadenylate Synthetase 1 (OAS1), an antiviral protein that facilitates innate immune responses against RNA viruses, is strongly associated with decreased rates of very severe COVID-19, hospitalization, and susceptibility in European populations [55]. Populations outside of Sub-Saharan Africa are known to harbor protective alleles rs4767027-T (OAS1 pQTL) and rs10774671-G (OAS1 sQTL) which differ due to evolutionary history; the rs4767027-T allele was derived from Neanderthal lineage whereas rs10774671-G allele is preserved from Neanderthal lineage. Additionally, the rs10774671-G allele regulates alternative splicing which increases levels of the OAS1 p46 isoform and has greater activity against the p42 isoform. The findings indicate that p46 isoform levels and protective effects on COVID-19 outcomes are positively associated; however, this finding is exclusive to the European population. Data is lacking on if the p46 ancestral allele preserved in Sub-Saharan Africans also offers protection. Here, it was shown that differences in plasma proteome has a genetic basis stemming from Neanderthal ancestry which offers benefit in COVID-19 risk modulation further emphasizing ancestral-based differences in proteome composition. The OAS1 forms prevalent in proteomes of different ancestral groups may offer variable immune responses when treated with nanotechnologically-based therapeutic. The increased potency of p46 against p42 could provide a genetic and biological basis into differences in disease susceptibility as well as drug metabolism for viral illnesses [55].
Considering genetic ancestry in future nanomedicinesThe field of nanomedicine could benefit from more research regarding the heritability of the proteome; there is very limited research investigating groups of similar geographical origin. Further research could include in vivo studies looking at heritability of proteome in mice. In addition, mice could be treated with NPs from generation to generation whilst observing changes in the protein corona and immune responses. Ex vivo studies investigating the protein corona around NPs could also be conducted with primary human plasma of similar geographical origin to continue to identify common genes linked to the proteome. This could also help diversify the proteome profile for ancestral groups and help refine models such as those used by Zhang et al. Lastly, a vast amount of these studies fail to consider that populations from different regions in Africa can vary widely in genetic ancestry, but are commonly grouped together in studies.
The effects of biological age on protein coronaDifference between chronological age and biological ageChronological age and biological age represent intertwined yet distinguishable concepts. It is crucial to unravel their intricacies, as both chronological and biological age exert unique influences on health outcomes, shaping our understanding of aging and its associated determinants. Chronological age, as defined by the amount of time elapsed since birth, serves as a fundamental determinant of health outcomes, chronic diseases, and mortality [56]. Biological age on the other hand, represents an individual's physiological state and is influenced by various factors, including genetics [57,
留言 (0)