To our knowledge, this is the first study evaluating the effect and differences between three different doses of CAF or PLA on the changes in the concentrations of CAF, PRX and PRX/CAF RATIO at different time-points in both the serum and saliva during a study protocol involving moderately trained people, additionally taking into account CYP1A2 enzyme activity and the CYP1A2 gene. We demonstrated that, in line with our hypothesis, the higher the CAF dose was, the greater the changes in the CAF and PRX concentrations in both the serum and saliva. Nevertheless, contrary to our hypothesis, the peak change in CAF concentration from pre-supplementation was elicited not 60 but 30 min after intake; CYP1A2 enzyme activity was not dose-dependent, and the CYP1A2 genotype had no interaction with enzyme inducibility.
As mentioned above, in our study, CAF and PRX concentrations were measured in two biological human matrices. We revealed moderate to strong correlations between serum and saliva CAF and PRX concentrations, indicating the possibility of the common use of less invasive saliva sampling under exercise conditions. However, with respect to CAF, the dependent variables (in saliva to serum) were explained in 79% and PRX in 61%. The good correlation observed in the literature between CAF concentrations in serum and saliva indicates that saliva analysis can be efficiently used to reliably reflect CAF concentrations in serum [28]. In a systematic analysis, Grzegorzewski et al. [14] performed linear regressions to quantify the relationships between saliva and blood-based CAF and PRX measurements. The resulting scaling factors and Pearson correlation coefficients between saliva to blood-based concentrations of CAF and PRX were 0.79 and 0.68 and 0.84 and 0.76, respectively.
Although current guidelines recommend CAF intake of 3–6 mgCAF/kgBM 30–60 min before the commencement of exercise [7], owing to its effect on the central nervous system, CAF is suspected to be the most ergogenic toward the end of exercise, when perceived effort is increased [29, 30]. Nevertheless, research has shown that after the plasma CAF concentration reaches its peak, it can remain at a steady elevated state even for several hours after intake [31].
Importantly, in contrast to our study, in the majority of research the assessment of CAF or CAF metabolites concentrations was performed solely in serum or plasma (as not the main but rather additional purpose), and blood was taken only at pre-supplementation time-point and one other time-point. Moreover, most exercise studies reported that the concentration of CAF at pre-supplementation time-point after 24 h of abstinence was usually ‘zero’; nevertheless, the measured levels may be in fact above zero [32] (ranging from 0.1 to even 5.0 µg/mL [33]). Although the participants were subjected to 24 h of abstinence, similar concentrations were also observed in our study; for this reason, we adopted the method of adjusting the values and assessing the changes in CAF and PRX concentrations from standardized zero levels at the pre-supplementation time-point. Furthermore, it has been shown that major ergogenic effects are unlikely to be induced with doses lower than 3 mgCAF/kgBM, where plasma levels are 2.91–3.88 µg/mL [34]. On the basis of only one previous study [31] analyzing CAF concentrations in plasma at three CAF doses, it was concluded that the MEDIUM CAF dose (6 mgCAF/kgBM) may increase CAF plasma levels up to 7.76 µg/mL and that the HIGH CAF dose (9 mgCAF/kgBM) up to 11.64–13.58 µg/mL after 60 min of intake. Additionally, it was simultaneously shown that the PRX level did not differ between the 6 and 9 mgCAF/kgBM, suggesting that hepatic CAF metabolism has been already saturated earlier.
Generally, previous studies, including exercise studies, have shown similar values of mean peak CAF concentrations in serum/plasma after CAF intake. The results ranged from 3.90 to 8.19 µg/mL [35,36,37,38,39,40,41] for CAF and from 1.08 to 1.60 µg/mL [36, 38, 42] for PRX. Nevertheless, studies have demonstrated that the time to the CAF peak has large inter-individual variations [43, 44]. Moreover, it was speculated that achieving peak serum/plasma concentrations prior to endurance exercise may not be the most important factor in the maximization of CAF ergogenic potential. The other possible explanation may be the CAF concentration at the site of action (in the central nervous system or muscle cells), which is currently impossible to measure. Another possible factor might be the effect of CAF metabolites or the achievement of peak concentration later in endurance exercise, when fatigue and effort perception are heightened [41].
In our study, the CAF concentration was markedly dependent on the CAF dose (the highest at HIGH dose). Interestingly, the peak change occurred at each dose from 0’ to 30’, and then, the level was maintained until 90’, suggesting serum CAF sustainability. This could be recognized as a valuable observation in the scientific and practical background. Simmonds et al. [38] demonstrated that 5 mgCAF/kgBM (in comparison with PLA) significantly elevated the CAF plasma concentration after 60 min of intake and improved supramaximal cycling in highly trained male cyclists. In a study by Sampaio-Jorge et al. [39], where recreationally trained male cyclists were supplemented with capsules containing 6 mgCAF/kgBM or PLA, significant time and treatment main effects on CAF serum concentrations were observed. However, again, blood samples were taken solely at pre-supplementation time-point and 60 min after the intake of CAF. In a study by Stadheim et al. [40] ten healthy highly trained male cross-country skiers were supplemented with 6 mgCAF/kgBM or PLA 75 min before the cross-country performance test (C-PT). Blood samples for CAF concentration analysis were taken more often - at rest, 35 min after ingestion, after the last workload on the incremental test and 1 and 14 min after the performance test. Ingestion of CAF increased plasma concentrations to approximately 7.8 µg/mL before 8-km C-PT, and, similarly to our results, it remained at a similar level throughout the exercise trial [40]. Furthermore, in a study by Mesquita et al. [45] blood samples were collected after the intake of 6 mgCAF/kgBM, immediately after the fatigue protocol and 6 h after intake, and the results demonstrated that the CAF serum concentration increased over time, with a peak level of ~ 8.51 µg/mL after 60 min of ingestion, which contradicts our results. Nevertheless, it is important to note that in this study, CAF was ingested in the form of capsules, not in powder form, as in our study. Additionally, the serum levels significantly decreased after the “fatigue protocol” (sets of 40 bilateral rebound jumps until task failure) and 6 h later. Bell et al. [46] examined the duration of the CAF ergogenic effect after the ingestion of 5 mgCAF/kgBM and registered significantly elevated CAF plasma concentrations throughout exercise. Nevertheless, this variable remained constant for trials conducted 3 and 6 h after ingestion, and its concentrations determined 1 and 3 h after ingestion were greater than those for the trial conducted 6 h after ingestion. In a more complex study, by Conway et al. [47], 3 or 6 mgCAF/kgBM (capsules) or PLA was administered 60 min before exercise, and additionally another 3 mgCAF/kgBM (where the previous dosage was also 3 mgCAF/kgBM) 45 min into exercise was given to the participants. Blood samples were drawn at pre-supplementation time-point, 30 and 60 min after the first supplementation; and additionally 30, 45, 60, and 90 min after the start of exercise and at the end of exercise (120 min after its onset). Similarly to the current work, CAF plasma concentrations increased in a dose-related manner. Its level was significantly elevated after 30 min but peaked within 90 min in the high-dose group. The lower CAF group exhibited an initial peak after 30 min, which was then followed by a slow decline until the second supplementation. Additionally, the CAF concentration was significantly higher in the higher dose group until 60 min into exercise, when the concentrations were similar [47]. In a study by Skinner et al. [41] 14 cyclists and triathletes consumed 6 mgCAF/kgBM or PLA either 60 min prior to completing a 40 km time trial or when the start of exercise coincided with the individual peak serum CAF concentration, which was determined from a separate ‘CAF profiling’ session involving monitoring CAF concentrations in the serum every 30 min over a 4 h period. Surprisingly, the CAF peak concentration (~ 6.4 µg/mL) occurred as late as 120/150 min after intake, but significant performance improvements were noted when CAF was consumed 60 min prior to exercise. It was then concluded that the ergogenic effect of CAF may not be related to the peak CAF concentration in the serum at the onset of endurance exercise. The second study by Skinner et al. [48] showed that the peak CAF serum concentration (~ 6.79 µg/mL) was also achieved as late as 120–180 min after the ingestion of 6 mgCAF/kgBM. In the third study by Skinner et al. [49] the serum was analyzed for the CAF concentration at 6 time-points over 4 h following 6 or 9 mgCAF/kgBM (fasted vs fed conditions). It was shown that this CAF time to peak occurred 60 min after ingestions for fasted trials compared with 120 and 180 min after ingestions for fed trials (carbohydrate consumption with CAF). Moreover, a higher dose resulted in a greater CAF peak in the fed condition than in the moderate dose in the fasted condition. The authors suspected that if an athlete was to consume a meal within 60 min of exercise, then a higher CAF dose may be needed to achieve the same serum concentrations as a moderate dose taken when fasted. Furthermore, Davenport et al. [30] investigated the optimal timing of a 200 mg CAF-containing supplement on exercise performance [a) 35 min before the 30 min steady-state cycling (SS); b) at the onset of SS, or c) immediately before the 15 min time trial]. Blood samples were taken at five time-points. CAF plasma concentrations increased under all experimental conditions, and there was a significant interaction effect, with peak values observed at different time-points [30]. Although the intake of 1.5-3 mgCAF/kgBM at around 60 min before the cycling time trial may improve performance [50], the same intake at approximately 180 min before the trial does not explicit similar results [51]. This finding suggests that elevated plasma CAF may not be the only mechanism responsible for improved performance but rather the optimal timing of the ingestion of a low dose of CAF.
In our study, the CAF dose increased the PRX concentration in the serum, and the change in the PRX concentration was also the greatest with the HIGH dose and increased gradually until 60’, when its peak was reached. There are only sparse studies measuring PRX concentration after CAF intake. Simmonds et al. [38] demonstrated elevated (p = 0.006) PRX plasma concentrations after 60 min of 5 mgCAF/kgBM intake in comparison with PLA. Similarly to our results, Conway et al. [47] showed that the PRX plasma concentration was significantly greater in the higher CAF dose group. However, the authors reported that the increase in the PRX concentration occurred at a slower rate, and no peak was observed, as it continued to increase throughout exercise [47]. Additionally, the PRX concentrations were greater with higher CAF intake and under fasted conditions. The authors suspected that if PRX contributes to the ergogenic CAF potential, the reduced PRX concentration observed following a pre-exercise meal may even limit the ability of CAF to enhance performance [49]. Davenport et al. [30] reported that PRX plasma concentrations increased over the duration of the trial and that there was a main effect for condition, with PRX concentrations being higher at the CAF intake point of 35 min before exercise. Interestingly, although CAF was elevated before and during the exercise test with all strategies, PRX was elevated only prior to exercise when the supplement was ingested 35 min before exercise (the most currently postulated strategy with ergogenic effects) [30].
In terms of saliva measurements, our study revealed that the change in the CAF concentration in saliva was also the greatest in HIGH dose. The peak change occurred from 0’ to 30’; nevertheless, in contrast to serum, we observed further significant differences between time-points, which suggested lower sustainability in saliva, which was probably influenced by other factors. Furthermore, the CAF dose increased the PRX concentration in the serum, and the change in the PRX concentration in the serum was also the greatest in HIGH dose. The saliva PRX peak was not achieved at the 60’ time-point for each dose, but increased gradually at least until the 90’ time-point. In another study, measuring saliva, Zawieja et al. [23] collected saliva samples at pre-supplementation time-point, 45 min post-ingestion of 5 mgCAF/kgBM and post-exercise (120 min post-ingestion) to quantify the CAF and PRX concentrations. The CAF concentration increased 30-fold and was approximately 20 times greater than that during the PLA trial at 45 min post-ingestion and decreased post-exercise. In turn, the PRX concentration increased 2.5 times 45 min post-ingestion and then increased further to the next time-point.
Notably, in our study, serum CAF concentration changes did not differ significantly between the 30’ and 60’ time-point, suggesting the possibility of attaining an earlier plateau, causing a peak in CAF was achieved before exercise. Davenport et al. [30] observed this at approximately 50 min after CAF ingestion, which is in contrast with other studies showing peak CAF concentrations between 75 and 120 min post supplementation [4, 47, 52]. Interestingly, it has been suggested that performance improvements caused by a reduction in perceived exertion may be mediated not only by CAF but also by increased PRX concentrations [30]. It may be concluded that when the plasma CAF concentration decreases, the PRX concentration may still increase, with no indication of a plateau even after 180 min [47]. Its increase in plasma occurs at a slower rate than that of CAF during the period of 60 min [32].
Additionally, we measured CYP1A2 enzyme activity via the mean time-corrected PRX/CAF RATIO in saliva and demonstrated that there was no CAF dose-dependency. Previously, a number of different biological human matrices and metrics, such as urine, plasma, serum, and saliva, have been employed, and CAF has been used as a probe for measuring CYP1A2 enzyme activity. The PRX/CAF concentration ratio at a single time-point correlates with apparent CAF clearance [12]. Metabolic phenotyping with PRX/CAF RATIO is time dependent (ratios increasing with time) and there is a clear CAF-dose dependency (smaller CAF doses increase the metabolic ratio) [14]. Although, it was demonstrated that to reliably assess CYP1A2 enzyme activity, no 24 h CAF abstinence is needed [12], in our study, this abstinence was followed by participants. The ‘gold standard’ for CYP1A2 enzyme activity estimation is considered to be the saliva molar PRX/CAF RATIO 6 h after CAF intake, owing to its best correlation with intrinsic CAF/PRX clearance [8, 24]. On the basis of these data, Urry et al. [8] developed a method enabling the adjustment of the CYP1A2 enzyme activity values to the optimal values and required 6 h post-dose sampling time-point. This novel technique accounts for the varied time intervals between CAF intake and saliva sampling. Perera et al. [12] quantified plasma and saliva CAF concentrations at different time-points after the ingestion of 100 mg of CAF. No significant difference was found between the PRX/CAF AUC0–24 ratio in plasma (0.79) and saliva (0.78) during the CAF abstinence period and during no abstinence (0.83 and 0.85, respectively). In particular, the PRX/CAF RATIO at 4 h demonstrated a strong correlation with all the reference metrics, such as CAF clearance or plasma CAF/PRX. Moreover, a study conducted in healthy males and females [53] assessing CAF as a probe for CYP1A2 enzyme activity reported no evidence of dose dependency. It was also concluded that even with high CAF consumption, steady-state concentrations are too low to saturate CYP1A2-mediated metabolism [13]. Considering the habitual intake of coffee, people who usually drink more than three cups/day usually have significantly higher normal CYP1A2 enzyme activity than individuals who usually consume less. This finding suggests that cessation of regular CAF intake before the measurement of CYP1A1 activity may have resulted in its decrease during the study; thus, it may not reflect an individual’s usual CYP1A2 activity [13].
In our study, the functional rs762551 polymorphism of the CYP1A2 gene was analyzed, but no interaction with CYP1A2 enzyme activity was found. It is a demonstrated determinant of inducible CYP1A2 activity, by which genotypes may be labeled ‘highly inducible’ (AA) and ‘less inducible’ (AC, CC) [8, 54]. On the basis of the literature, AA homozygotes may metabolize CAF more quickly and perceive a greater ergogenic effect [55]. In this case, it would appear that PRX may be a more potent adenosine-receptor antagonist, which is elevated sooner in the AA genotype [30]. Nevertheless, several authors [8, 56,57,58,59] have demonstrated that there is no significant effect of this SNP on CYP1A2 enzyme activity, as measured by the PRX/CAF RATIO or the mean time-corrected PRX/CAF RATIO, indicating that this CYP1A2 genotype is not a robust predictor of the CYP1A2 phenotype. The − 163 C > A polymorphism is correlated with RATIO only in smokers [58], since the more pronounced increase in CYP1A2 enzyme activity caused by this genetic variation is only observed in current smokers. Smokers with the − 163 C/C genotype have been shown to have 40% lower plasma PRX/CAF RATIO than those with the − 163 A/A genotype, while no influence of this polymorphism has been detected among nonsmokers [57]. Interestingly, according to Davenport et al. [30] although regular exercise increases CYP1A2 expression, it seems that ‘slow metabolizers’ may be able to overcome any genetic disadvantage with training to increase CYP1A2 enzyme activity sufficiently.
Limitations
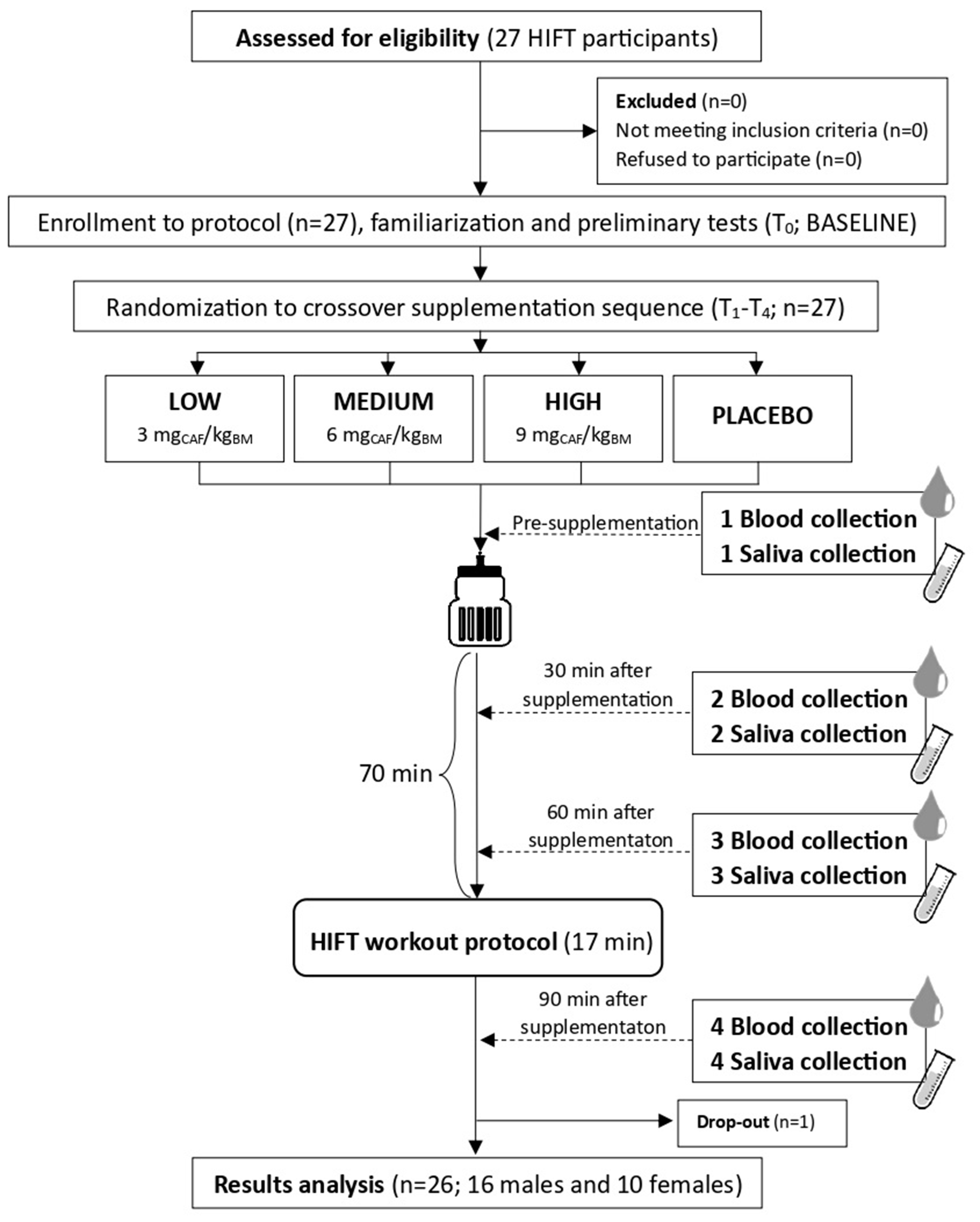
Notably, our research has several limitations. Firstly, we examined the CAF and PRX concentrations at different time-points over a relatively short period of time (0–90 min); the sampling was not constant over a long period of time, which prevented us from measuring the AUC, and we were not able to measure the possible peak of PRX, as it is believed to peak further than 90 min. Secondly, examining the PRX/CAF RATIO only at these time-points probably influenced the possibility of adequate CYP1A2 enzyme activity measurement. It is established that early and late time-sampling are least suitable for phenotyping because at these time-points, concentrations may be low, resulting in relatively high random errors and thus a low single to noise ratio. Additionally, in this early stage, the distribution phase of the substance and its absorption and kinetics may further influence the outcome of metabolic ratios. Moreover, metabolic phenotyping with PRX/CAF RATIO is strongly time dependent (the ratio increases with time), and smaller CAF doses and concentrations increase the metabolic ratio (explaining why BASE and PLA values were profoundly higher) [8, 14, 32]. Nevertheless, as we were aware of this methodological issue, we used a previously developed method to adjust the CYP1A2 enzyme activity to the optimal 6 h after CAF administration sampling [8]. Moreover, the CYP1A2 genotypes results should be accounted with caution due to relatively small sample of ’fast metabolizers’ among the studied participants. Nevertheless, in accordance with the Hardy-Weinberg principle, the frequency of CYP1A2 SNP in the population is approximately 45% for AAs, 45% for ACs and 10% for CCs [9]. Finally, although we included both males and females in the study protocol, because of the participation of both sexes in CrossFit/HIFT training, we were not able to perform sex-dependent analyses differentiating CAF effects between sexes (uneven groups).
Strengths
The unquestionable strengths of our study were its multiple crossover design protocol and the implementation of three different CAF doses, as well as the PLA, in HIFT-trained participants. Secondly, as the first study, we analyzed both serum and saliva CAF and PRX concentrations at different time-points in moderately trained people and measured their CYP1A2 enzyme activity. Thirdly, taking into account the possibility of higher than zero CAF pre-supplementation concentrations, we assessed the effects of CAF on CAF and PRX changes from the pre-supplementation time-point. Furthermore, we used a saliva to serum regression model, which to some extent accounted for changes in CAF or PRX concentrations with time in saliva (concentrations decreased over time) and serum (more constant). In addition, we calculated the required sample size, which was met by the number of participants who took part in the study. Furthermore, all study participants were habitual CAF users who maintained 24 h CAF abstinence and were not smokers. We also ensured full compliance with the intake of supplements and thorough standardization of the procedures.



留言 (0)