
記住我








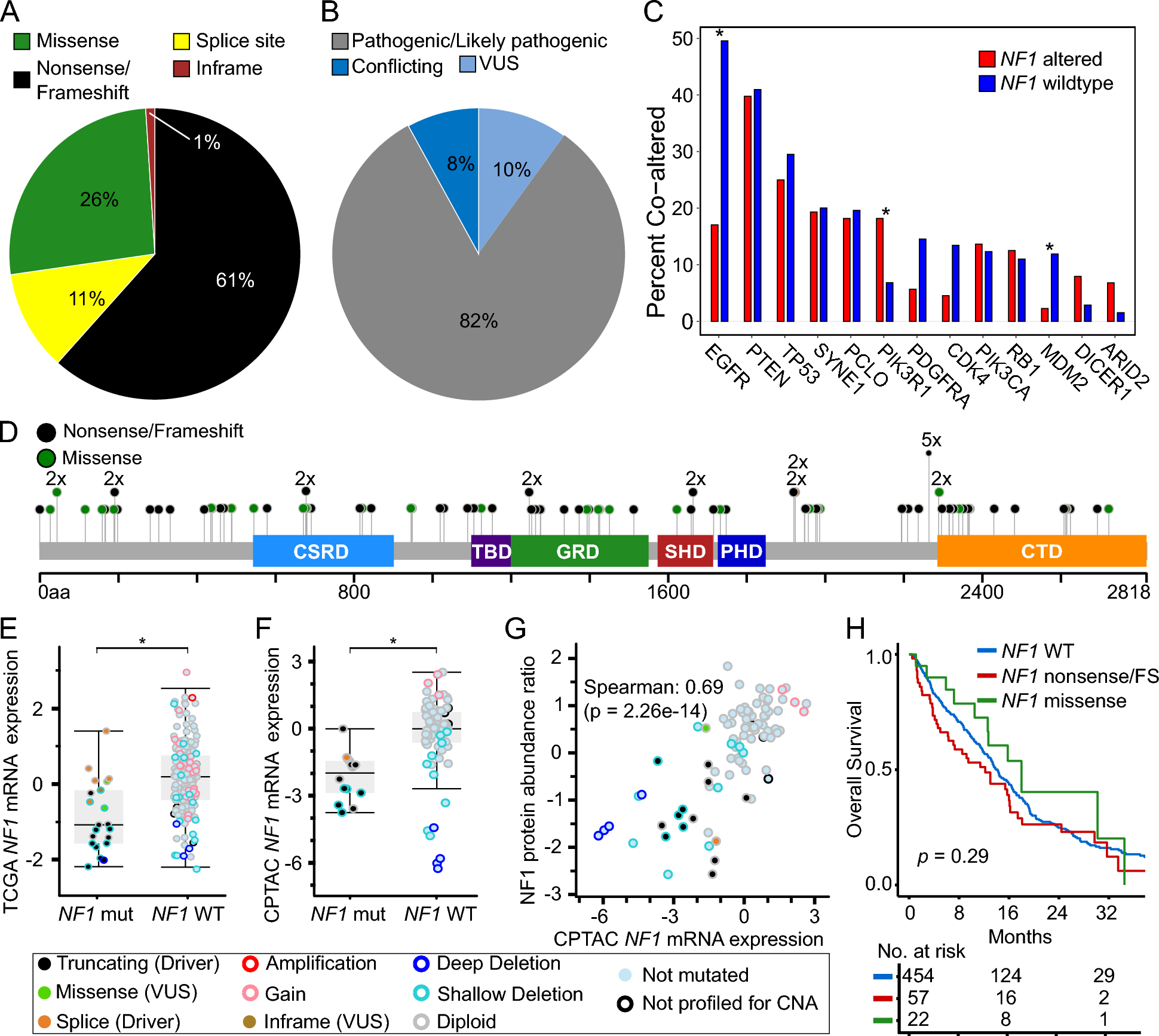
We first assessed the frequency and type of NF1 alteration among 542 IDH-wildtype GBM. One hundred fifteen NF1 alterations were identified across eighty-eight tumors (16.2%) (Supplementary Table 1). Seventy-one nonsense/frameshift mutations, 13 splice-site mutations, 30 missense mutations, and one in-frame insertion/deletion in NF1 were annotated (Fig. 1A). Protein-coding changes in NF1 were distributed throughout the entire gene, with the exception of recurrent truncations at Y2264 which were observed in five tumors. Seventy-two NF1 alterations were identified by ClinVar to be either “Pathogenic” or “Likely pathogenic”, with 16 alterations of conflicting or uncertain significance and 27 not annotated [27]. Of the 30 NF1 missense mutations detected in our cohort, seven were predicted to be cancer-promoting and the rest were predicted to be likely passenger mutations by FATHMM [28].
Fig. 1
Genomic and clinical features of NF1 loss in glioblastoma. A Distribution of NF1 alteration types and B ClinVar classification of NF1 alterations detected in JHH GBM cohort (N = 115). C Co-alteration frequencies between NF1 and other selected genes in JHH GBM cohort. Statistical significance was assessed using two-tailed Fisher’s Exact test. D Lollipop plot of NF1 alterations detected in JHH GBM cohort. Box and whisker plots comparing NF1 mRNA expression between NF1 mutant and NF1 wildtype glioblastomas in E TCGA (z-scores, U133 microarray) and F CPTAC (z-scores, log2FPKM) GBM cohorts. Statistical significance was assessed using Welch’s t-test. G Scatter plot comparing NF1 protein abundance ratio (z-scores) and NF1 mRNA expression (z-scores, log2FPKM) in CPTAC GBM cohort. H Kaplan–Meier survival curves displaying overall survival in JHH GBM cohort stratified by NF1 alteration type. VUS = variant of unknown significance, FS = frame shift
NF1 alterations were mutually exclusive with alterations in EGFR (log2OR –2.3, p-adj < 0.001), MDM2 (log2OR –2.5, p-adj = 0.05), PDGFRA, and CDK4 (Fig. 1B, Supplementary Table 2). Alterations in NF1 co-occurred with alterations in PIK3R1 (log2OR 1.6, p-adj = 0.03). After adjusting for multiple hypotheses, only the associations between NF1 and EGFR, PIK3R1, and MDM2 remained statistically significant, supporting prior observations [8]. Mutual exclusivity of NF1 and EGFR alterations in IDH-wildtype glioblastoma was further substantiated using two publicly available resources, TCGA and CPTAC, totaling 368 genomically-characterized glioblastomas after excluding samples with pathogenic IDH1/2 alterations (Supplementary Table 3–4) [21,22,23,24].
To identify whether pathogenic alterations in NF1 are associated with changes in mRNA or protein expression, we interrogated the TCGA and CPTAC glioblastoma cohorts [21, 22]. We observed significantly decreased average NF1 mRNA expression (p < 0.001 for each cohort) in tumors with known pathogenic NF1 alterations, though a subset had intact expression levels (Fig. 1E, F). We next evaluated whether NF1 mRNA expression correlated with protein expression. In 89 glioblastoma samples with both protein and mRNA, mRNA expression levels strongly correlated with protein expression (Spearman = 0.69, p < 0.001) as quantified by mass spectrometry (Fig. 1G).
We evaluated how NF1 alterations were associated with overall survival. In our institutional cohort of 542 glioblastomas, we observed that neither truncating NF1 alterations in the entire cohort nor NF1 genomic loss in the TMA cohort alone were significantly associated with decreased overall survival (Fig. 1H). This was supported by the TCGA dataset where neither NF1 alterations nor mRNA expression was associated with a difference in overall survival (Supplementary Fig. 2A-B). Our findings contrasted with the CPTAC dataset where NF1 alterations were associated with decreased overall survival, though this was not true for NF1 mRNA or protein expression (Supplementary Fig. 2C-E) [23, 24].
Tissue microarray assembly and genomic characterizationIn order to evaluate NF1 protein expression, we created a tissue microarray (TMA) from 69 representative glioblastomas from the cohort described above. Forty-four samples were NF1-wildtype and 25 were NF1-mutant on next-generation sequencing. All but one was obtained from the first tumor resection. The mutational profiles of glioblastomas included in the tissue microarray cohort are displayed in Fig. 2. The range of genomic alterations and their association with NF1 loss were similar to the overall GBM population. NF1 alterations identified in our cohort were manually assigned to two categories (pathogenic or non-pathogenic) by a neuropathologist based on whether they were indicative of genomic loss, taking into account alteration type, location in the gene, and potential loss of heterozygosity events. Tumors in the TMA were categorized as “NF1 altered” or “NF1 intact” by a neuropathologist based on their alterations. These scores were in high concordance (100%) with ClinVar annotations.
Fig. 2
Oncoprint of 69 glioblastomas included in TMA cohort displaying genetic alterations in selected genes
Immunohistochemistry for NF1 statusWe performed an exhaustive literature search for NF1 antibodies previously validated for immunochemistry in human glioma tissue. After thorough validation, we chose two antibodies: clone NFC (Sigma Aldrich, MABE1820) and iNF–07E (Infixion Biosciences) to evaluate NF1 protein expression (Fig. 3, Supplementary Fig. 3). Another NF1 antibody, clone McNFn27b (Abnova, NB300-154), was tested but not used due to high background staining in our hands. Immunostaining with the NFC antibody was negative in the tissue specimen from a patient with NF-1 (Fig. 3A), but immunoreactive in a specimen with no NF1 alterations (Fig. 3B). The NFC immunostain was immunoreactive in 26 tumors (41%), whereas 34 (54%) had minimal or absent staining, one had equivocal staining, and seven were not scored due to the absence of assessable tumor tissue. Inter-core heterogeneity was noted within tumors, with seven displaying a mixture of cores showing both retained and absent NFC immunoreactivity. NF1 immunostaining scored by a second independent neuropathologist (F.J.R.) was highly concordant (82%). NF1 immunostaining results were evaluated for each tumor in the context of NF1 genomic status (Fig. 3E). Of the 14 scorable tumors assessed to be NF1-deficient by NGS, 12 (86%) also demonstrated minimal to absent NF1 immunostaining with NFC antibody. Of the two tumors with intact NF1 immunostaining, one harbored an E2624* truncating mutation near the C terminus. This truncation preserves a portion of the NFC antibody epitope and, of note, was the most C-terminal truncation present in our cohort. Among scorable NF1-wildtype specimens, the NFC antibody was immunoreactive in 23/42 (55%), equivocal in one (2%), and minimal or absent in 18 (43%). In tumors harboring NF1 variants of uncertain significance, NF1 immunostaining was minimal or absent in four and immunoreactive in one.
Fig. 3
NF1 immunohistochemistry in patient-derived glioblastoma samples. Representative IHC shows loss of NF1 immunostaining in NF1 −/− tumor (patient with neurofibromatosis type 1) with A NFC antibody and C iNF-07E antibody. Representative IHC shows retained NF1 immunostaining in NF1 +/+ tumor with B NFC and D iNF-07E. E Plot of NF1 genomic status and corresponding NF1 IHC scores using two NF1 antibodies. F, G I Bar plots demonstrating no difference in expression of candidate surrogate biomarkers selected a priori—phospho-ERK, phospho-S6, p16, podoplanin (PDPN), ATRX, Ki-67, p53—between tumors with retained and lost NF1 immunostaining
The iNF-07E antibody was appropriately negative in the tissue specimen from a patient with NF-1 (Fig. 3C), but immunoreactive in a specimen with no NF1 alterations (Fig. 3D). The iNF-07E antibody was immunoreactive in 46 tumors (76%). Fourteen (23%) demonstrated minimal or absent immunostaining, one stained equivocally, and seven were not scored due to the absence of assessable tumor tissue. Inter-core heterogeneity was minimal with iNF-07E immunostaining, as no tumors exhibited cores with both intact and absent staining. Of the 14 scorable tumors considered NF1-deficient by NGS, eight (57%) also demonstrated minimal or absent immunostaining with iNF-07E. Among scorable NF1 wildtype specimens, NF1 immunostaining was retained in 38 (90%) and minimal or absent in 4 (10%). In tumors harboring NF1 variants of uncertain significance, NF1 immunostaining was minimal or absent in two and retained in three.
Upon comparison, the sensitivity of NFC and iNF-07E for tumors with confirmed NF1 loss was 86% and 57%, respectively (Supplementary Table 5).
Protein and genomic correlates of NF1 lossWe next evaluated potential protein correlates for NF1 loss as identified via immunostaining or NGS. We assessed the expression of proteins known to be correlated with MAPK signaling (phospho-ERK, phospho-S6) or with NF1 status (p16, podoplanin) [29, 30]. We also evaluated proteins associated with proliferation and loss of cell cycle regulation in glioblastoma (p53, Ki-67) [30]. Expression of these proteins did not correlate with NFC or iNF-07E staining. We next asked whether IHC would be associated with NF1 genomic status and found positive phospho-ERK immunostaining to be associated with NF1 genomic loss (p = 0.045, Supplementary Table 5), supporting prior observations of a similar correlation in mesenchymal GBM [21]. Unsupervised hierarchical clustering by these seven proteins of interest did not identify distinct clusters corresponding with NF1 immunostaining (Supplementary Fig. 4). We evaluated for co-occurring or mutually exclusive gene alterations and found no statistically significant associations with NF1-deficiency by IHC staining.
Survival analysis of tissue microarray cohort stratified by NF1 immunostainingWe evaluated whether NF1 loss by IHC—as assessed by two different antibodies—correlated with survival. Baseline demographic features of tumors included in the TMA were comparable between the two groups, demonstrating no differences between the two antibodies except for higher baseline Karnofsky performance score observed in patients with loss of immunostaining by iNF-07E antibody (p = 0.025) (Supplemental Table 7). Notably, patients with NF1 loss by NFC immunostaining had decreased survival compared with NF1 intact tumors (8.5 (4.5–12.6) vs. 16.4 (13.2–25.4) months, p = 0.011; Fig. 4A). This difference was not observed with the iNF-07E antibody (9.8 (6.5—18.0) vs. 11.4 (4.5—NR) months, p = 0.63). In a Cox proportional hazard regression model, we adjusted for known and potential confounding variables: age at diagnosis, sex, MGMT promoter methylation, baseline Karnofsky performance score, treatment with temozolomide or radiotherapy, extent of resection, and the presence of any NF1 genomic alteration (Supplementary Table 8). Minimal or absent NFC immunostaining remained significantly associated with decreased overall survival (HR 3.23, 95% CI 1.29–8.06, p = 0.01).
Fig. 4
Kaplan–Meier curves displaying overall survival in TMA cohort by NF1 immunohistochemistry. Curves are stratified by A NFC immunostaining, B iNF-07E immunostaining among entire TMA cohort
Protein and genomic correlates NF1 expression in NF1-wildtype subsetWe evaluated whether loss of NF1 expression in NF1-wildtype tumors was associated with other gene alterations on NGS or IHC stains. We focused our analysis on the subset of NF1-wildtype tumors that demonstrated minimal to absent staining by NFC antibody. None of the genes or proteins that we tested were significantly associated with NF1 expression in this subset (Supplementary Tables 9–10). Notably, despite the absence of associated gene alterations, minimal or absent NF1 protein by NFC remained significantly associated with decreased overall survival in patients without genomic NF1 alteration (Fig. 4B). Loss of NF1 expression remained significantly associated with decreased survival on multivariate analysis including age at diagnosis, KPS, degree of resection, prior chemoradiation, and MGMT promoter methylation status (HR 2.55, 95% CI 1.00–6.49, p = 0.049, Supplementary Table 11). On univariate analysis, age at diagnosis, KPS less than 80, subtotal resection, lack of radiation/temozolomide, and unmethylated MGMT promoter were also associated with inferior survival, but no other immunostain or genomic alteration was significantly associated with outcome.
留言 (0)