
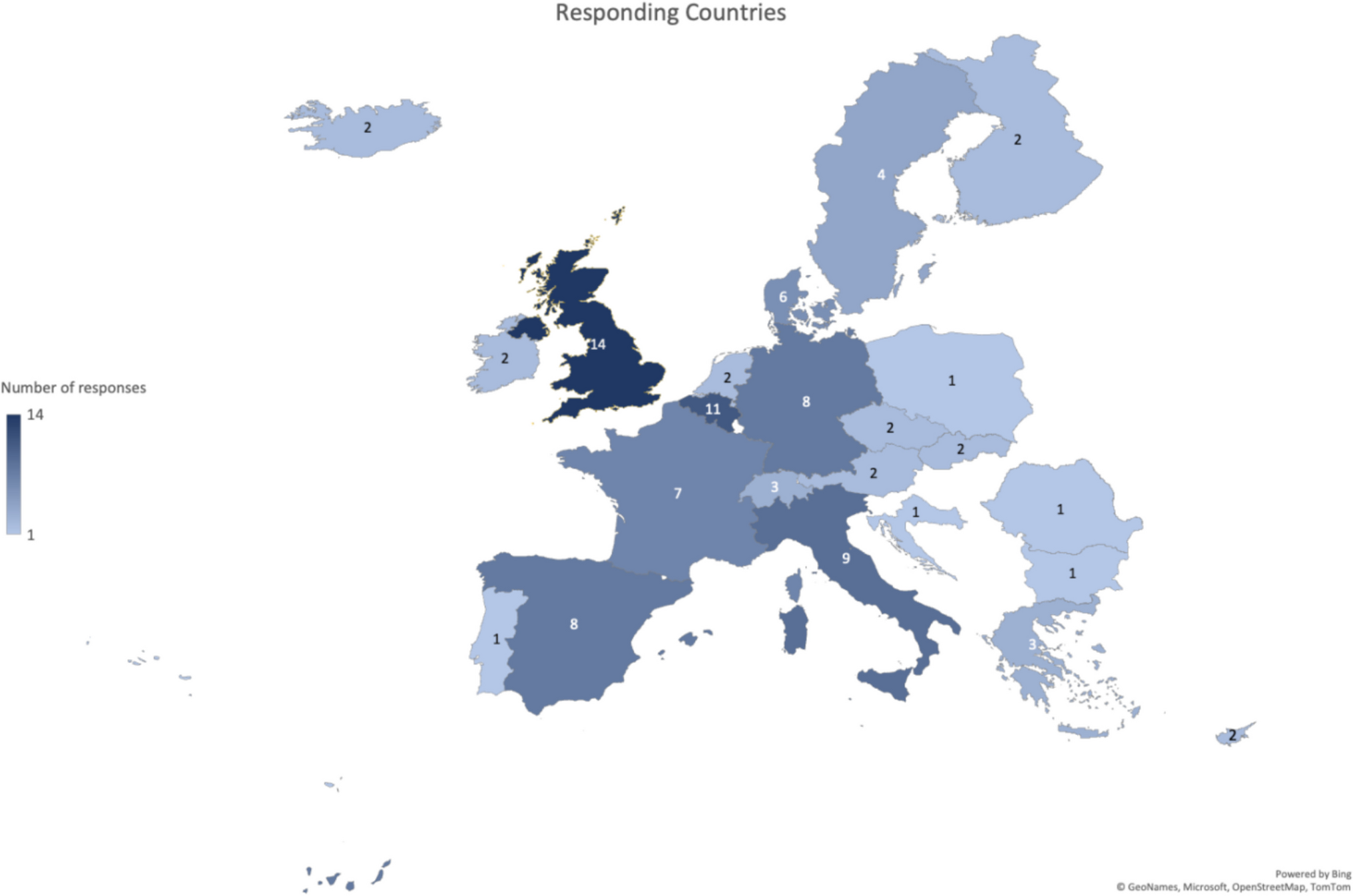
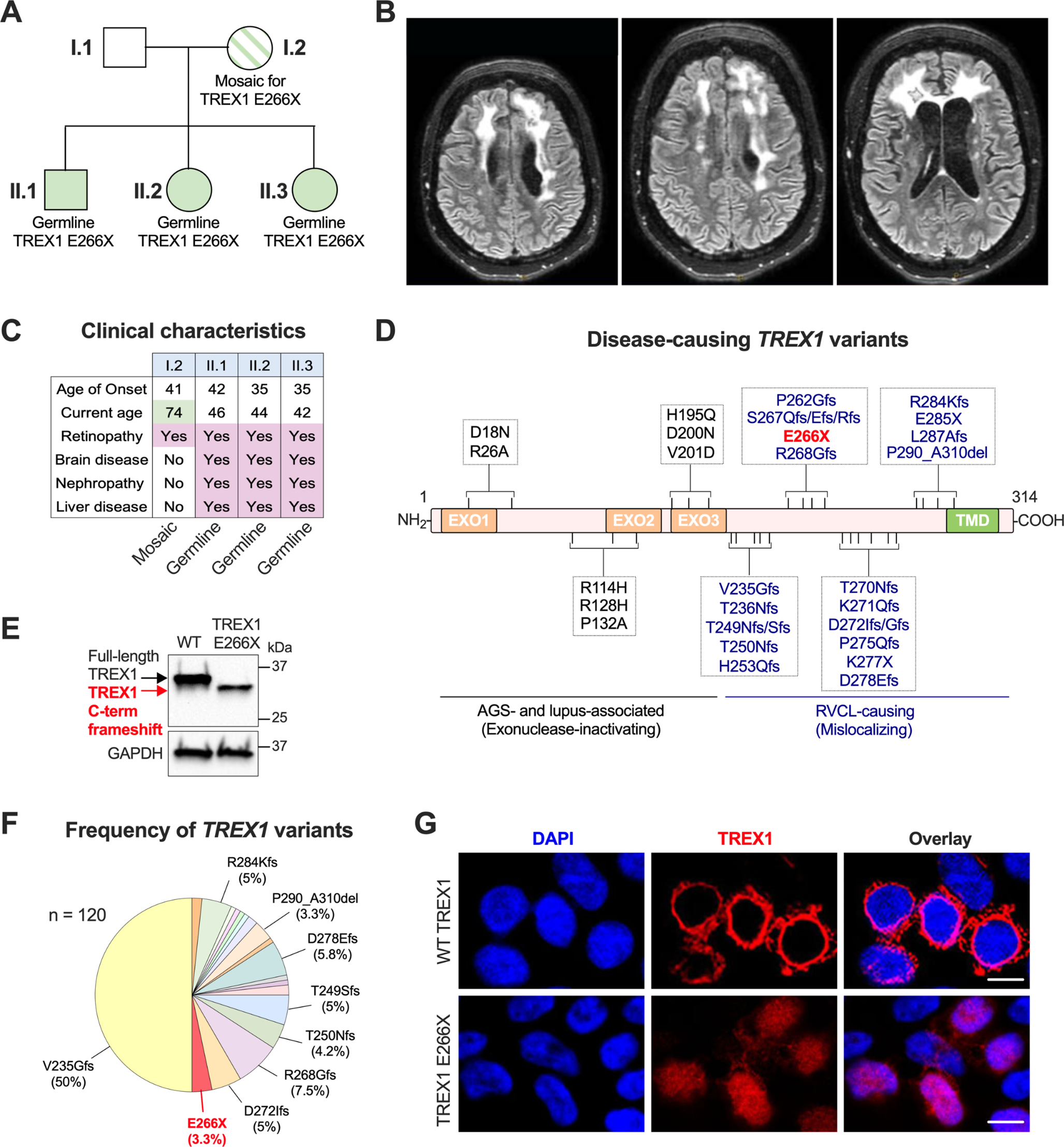
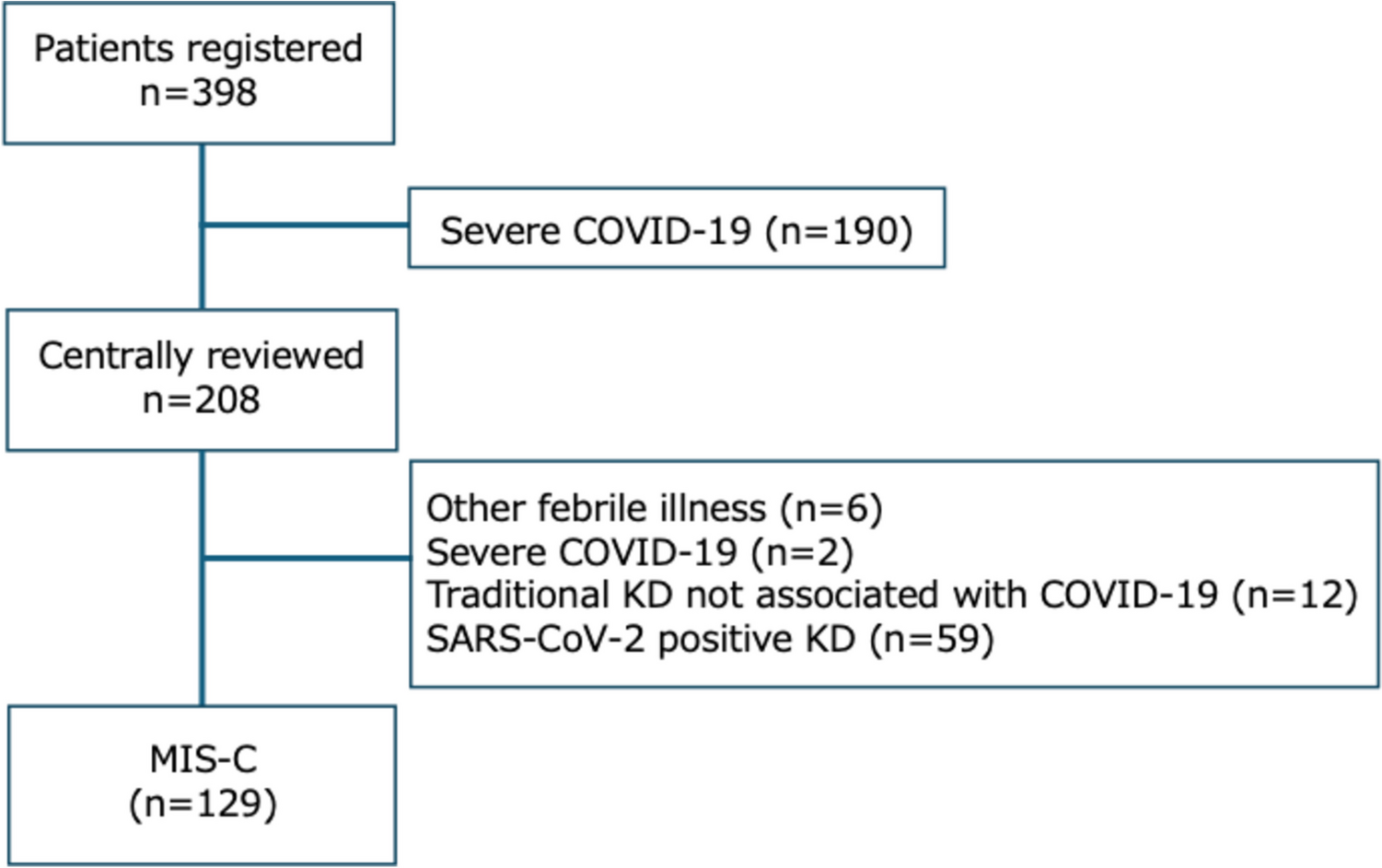
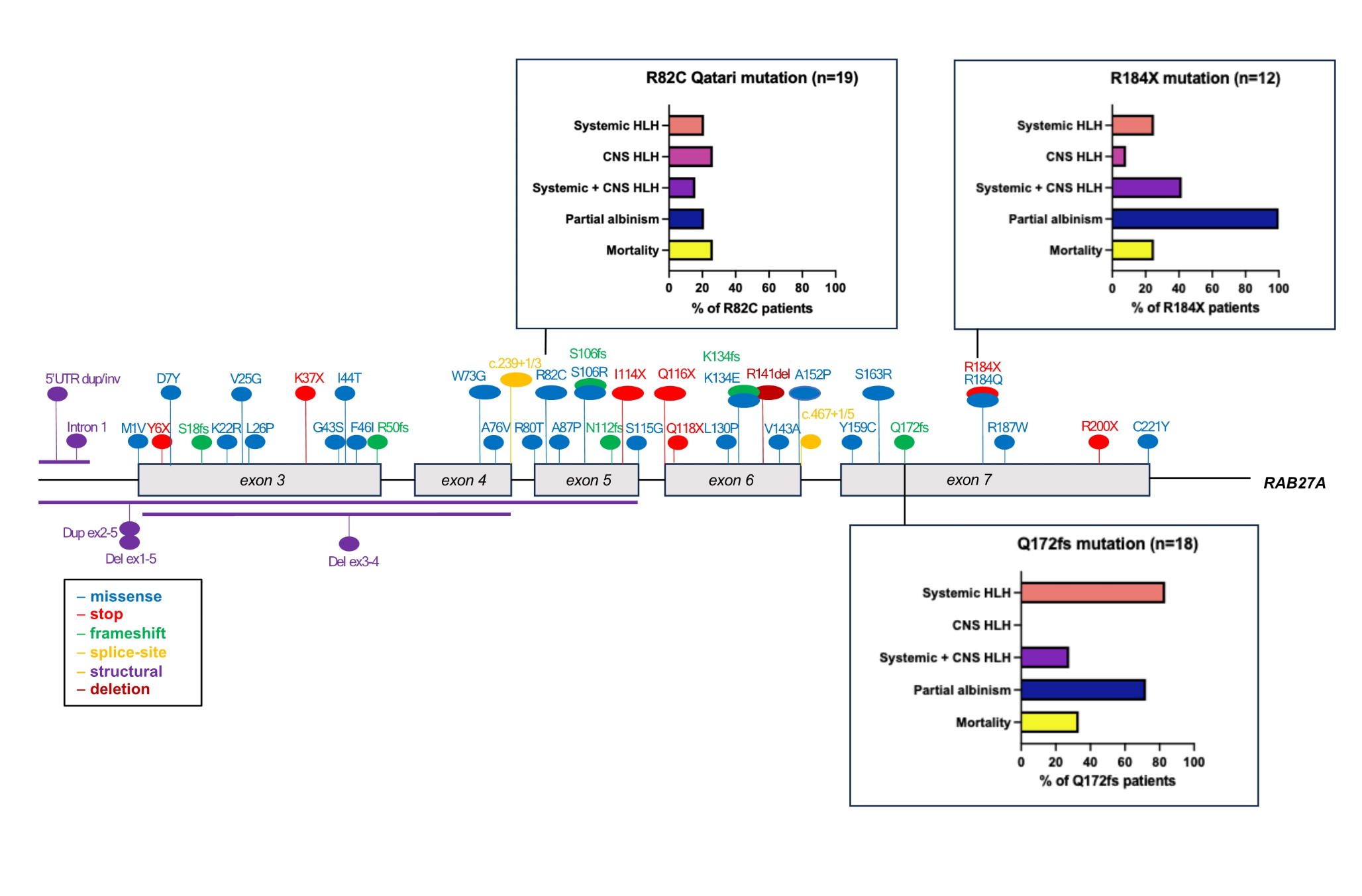
記住我


To asses histopathological differences between granulomas in CVID and those from other granulomatous diseases, we performed a blinded scoring of histological parameters on HE stained sections. The biopsies obtained from CVID patients (n = 6) were compared to biopsies obtained from sarcoidosis, PS and TB patients (n = 6 for all groups). The histological parameters examined included: granuloma size, clustered granulomas, solitary granulomas, circumscribed granulomas, confluent granuloma areas, presence and subtypes of multinucleated giant cells (foreign body giant cells, Touton giant cells, Langhans giant cells, asteroid giant cells), lymphocyte infiltration into granulomas, fibrosis in granulomas and adjacent surroundings, and necrotic areas in granulomas (Table 1; Fig. 1A).
Table 1 Histological scoring of granulomas in CVID, sarcoidosis, PS and TBFig. 1
Representative HE stains and granuloma diameter (A) 20x Lymph node HE stains of representative biopsies of CVID, sarcoidosis (Sarc), PS and TB. (B) Granuloma size per disease group based on the diameter of 30 randomly selected granulomas for each disease group. Median and interquartile ranges are indicated. Kruskal-Wallis with Dunn’s multiple comparison correction, *=P < 0.05, **=P < 0.01, ****=P < 0.0001. (C) Discriminative capacity granuloma sizes to discriminate CVID granulomas from sarcoidosis, PS or TB, shown by ROC analysis. See also Table 2
Table 2 Discriminative capacity of granuloma size for detecting CVID granulomasCVID granulomas were significantly smaller than granulomas from the other diseases (Fig. 1B). Moreover, granuloma size served as a discriminating parameter distinguishing CVID from sarcoid granulomas with a granuloma size < 191 μm highly sensitive and specific, as well as from PS or TB granulomas (Fig. 1C; Table 2). CVID granulomas were less solitary and more organized in clusters with confluent areas, were less well circumscribed, and contained less fibrosis (Fig. 1A; Table 1) as opposed to sarcoidosis, PS and TB granulomas, which presented more commonly as solitary and well-circumscribed. Multinucleated giant cells are considered a typical hallmark of granulomas and were indeed commonly observed in granulomas from sarcoidosis, TB and PS [8, 23, 25]. However, granulomas from CVID patients seldom contained multinucleated giant cells (Fig. 1A; Table 1). Overall, granulomatous disease in CVID seems to reflect a less organized cellular process compared to sarcoidosis, PS and TB, and lacks hallmarks linked to granuloma maturation like fibrosis and multinucleated giant cells.
CVID, Sarcoid, PS and TB Granulomas are Enriched for Proteins Related to Myeloid and T Cell FunctionTo assess differences in immune biology between CVID granulomas and granulomas from sarcoidosis, PS and TB, we performed digital spatial profiling for proteins associated with immune cell profiles, immune activation¸ PI3K/AKT-signaling and MAPK-signaling both inside the granulomas as well as in their adjacent surrounding (Table S1).
No distinct clustering was observed based on diseases group, region, or tissue origin (Figure S1). Comparing the relative protein expression inside the granulomas and their adjacent surroundings per disease, we observed commonalities across all diseases (Fig. 2A). CD68 (expressed by macrophages), CD11c (expressed by dendritic cells), CD127 (expressed by T cells, typically memory T cells), CD44 (expressed by myeloid and epithelial cells; involved in cell adhesion and migration), and PD-L1 (inhibitory molecule, mainly expressed by antigen presenting cells (APCs)) were significantly enriched with increased absolute counts inside the granulomas in all disease groups (Fig. 2A and B). Also the absolute counts of CD80 (expressed by APCs and activated B cells among others; ligand for CD28 and CTLA4, modulating T-cell activation and differentiation), CD40 (costimulatory receptor expressed by APCs, involved in B-cell memory development and germinal center formation), and p44/42MAPK ERK1/2 (MAPK pathway) showed significantly increased counts inside granulomas compared to its surroundings in all disease groups (Fig. 2B), even though not significantly enriched in all disease groups (Fig. 2A).
Fig. 2
Spatial protein analysis of inter disease comparison granuloma versus surrounding. (A) Relative protein expression of 72 granulomas (3 granulomas per each of 6 patients per disease group) and the adjacent surrounding. Fold induction threshold was set to 1 (red line), significance threshold at 1.2 (green line) showing uncorrected P-values. Only significantly or borderline significant targets are indicated. (B) Absolute protein counts with median of significant protein expression commonalities in the granulomas of the four diseases. (C) Absolute protein counts with median of protein targets enriched in the granulomas versus the surrounding only in a subset of the volcano plots. (D) Absolute protein counts with median of protein targets enriched in the surrounding versus the granulomas only in a subset of the volcano plots. Statistical differences only indicated for expression in granuloma versus the surrounding per disease, with statistical testing by Mann-Whitney test with * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001
PD-L2 (which inhibits T-cell proliferation, expressed by APCs among others) was enriched only in CVID granulomas versus their surroundings, and FAPα (fibroblast activating protein-α) appeared enriched in CVID and sarcoid granulomas versus their respective surroundings (Fig. 2A and C). Only sarcoid granulomas had a significant enrichment and protein count of EGFR (epidermal growth factor receptor), and of the PI3K pathway related proteins phospho-PRAs40 and phospho-tuberin (Fig. 2A and C).
For PS and TB, significant enrichment for fibronectin (an extra cellular matrix protein involved in cell adhesion and migration), CD163 (an acute phase regulated receptor expressed by monocytes and macrophages), and CD34 (an adhesion molecule expressed by endothelial cells, fibroblasts and fibrocytes among others) was detected in the surrounding of the granulomas (Fig. 2A and D). SMA (smooth muscle actin, expressed by myofibroblasts mainly), CD3 (general T cell marker), and CD8 (expressed by cytotoxic T cells) were significantly enriched in the surrounding of TB granulomas, with the absolute protein counts of SMA and CD3 also significantly increased in sarcoidosis (Fig. 2A and D). In both CVID and sarcoidosis, no relative enrichment of proteins in the surrounding versus the granulomas was observed (Fig. 2A).
The above data suggest commonalities in the immune pathogenesis of the granulomas. However, we also observed differences, as reflected by the distribution of fibroblast-associated and some hematopoietic markers between the four diseases. This could reflect differences in the local inflammatory milieu between CVID and the other granulomatous diseases.
CVID Granulomas are Enriched for CD163, CD66b and FAPα and have Reduced Expression of Fibrosis-Associated Proteins in their SurroundingWe next explored differentially expressed proteins inside the CVID granulomas or their surroundings to sarcoidosis, PS and TB. Comparing the protein expression inside the granulomas, CD163 was enriched inside CVID granulomas compared to sarcoid granulomas (Fig. 3A). Absolute protein count of CD163 was highest in CVID, but also in TB the expression was significantly higher compared to sarcoidosis. Interestingly, sarcoid granulomas exhibited a significant enrichment for proteins related to the MAPK signaling pathway when compared to CVID granulomas, including BRAF, phospho-cRAF, p44/42 MAPK ERK1/2, and borderline for phospho-MEK1 (Fig. 3A). These increased MAPK pathway related proteins were unique for sarcoid granulomas (Fig. 3B, Figure S2), even though p44/42 MAPK ERK1/2 was increased in the center of all granulomatous diseases (Fig. 2B). Additionally, SMA was significantly increased in sarcoidosis versus CVID granulomas (Fig. 3A and B).
Fig. 3
Spatial protein analysis in the granulomas of CVID versus granulomas of sarcoidosis, PS and TB. (A) Relative protein expression of 72 granulomas, comparing CVID versus sarcoidosis, or PS or TB, with fold induction threshold set to 1 (red line), significance threshold at 1.2 (green line), showing uncorrected P-values. Only significantly altered or borderline significant targets are indicated. B-D) Absolute protein counts with median of significantly enriched protein differences between CVID versus either (B) sarcoidosis, (C) PS or (D) TB. Statistical testing was performed by Kruskall-Walis with Dunn’s multiple comparison correction, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001
Compared to PS, CVID granulomas were significantly enriched for CD66b (marker for granulocyte activation, mainly expressed by neutrophils), although not detectable in absolute protein counts. Furthermore, CVID granulomas were significantly enriched for FAPα compared to PS granulomas (Fig. 3A), with absolute counts showing a significant increase in both CVID and sarcoid granulomas compared to PS (Fig. 3C). CD3 was significantly enriched inside CVID granulomas compared to TB (Fig. 3A). The absolute protein counts of CD3 were significantly lower in TB granulomas versus all others (Fig. 3D).
Comparing the surroundings revealed some proteins differently expressed in CVID compared to the other granulomatous diseases. Fibronectin was consistently more abundant in the surrounding of the other diseases than CVID, evident from both the significant enrichment and increased protein counts in sarcoidosis, PS, and TB (Figs. 2 and 4A and B). Also, SMA was significantly enriched and increased in the surrounding of sarcoid and TB granulomas as compared to CVID (Fig. 4A and B). BRAF was significantly enriched and increased in the surrounding of sarcoidosis versus CVID (Fig. 4A and B). Cytokeratin (PanCK, expressed by epithelial cells) was enriched in the surrounding of PS granulomas compared to CVID, as for the protein counts which also showed a significant increase in the surrounding of TB compared to CVID (Fig. 4A and B). CD163 was significantly increased only in the surrounding of TB compared to CVID (Fig. 4A and E).
Fig. 4
Spatial protein analysis of surrounding areas of granulomas of CVID versus sarcoidosis, PS and TB. (A) Relative protein expression of 72 granulomas, comparing CVID versus sarcoidosis, or PS or TB, with fold induction threshold set to 1 (red line), significance threshold at 1.2 (green line), showing uncorrected P-values. Only significantly altered or borderline significant targets are indicated. (B) Absolute protein counts with median of significantly depleted protein differences in the surrounding areas of CVID versus sarcoidosis, PS or TB. Statistical testing was performed by Kruskall-Walis with Dunn’s multiple comparison correction, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001
To conclude, targeted protein analysis inside CVID granulomas showed differences compared to the other granulomatous diseases, although these differences were not always unique for CVID. On the other hand, in the surrounding tissue of CVID granulomas lower expression of fibrosis-associated proteins fibronectin and SMA were observed, in line with the limited fibrosis observed for CVID in histological analysis (Fig. 1; Table 1).
CVID Granulomas Display a Distinct Distribution of Neutrophils, Myeloid Derived Cells and Fibroblasts Compared to Other Granulomatous DiseasesTo better understand the cellular organization and distribution of CVID granulomas and their surrounding tissue, we conducted three immunofluorescence (IF) multiplex assays on subsequent biopsy slides, to observe how myeloid cells (CD68+, CD163+, CD11c+, and MPO + cells) and fibroblasts-like cells (FAPα + cells) were distributed (Fig. 5). We also included certain protein targets from the previous spatial protein analysis (PD-L1+, SMA+, p-ERK1/2+) (Fig. 5, Figure S3).
Fig. 5
Cellular distribution and quantification of myeloid, fibroblast-like and lymphoid cells in the granulomas and their surroundings. (A) Heat map showing the median of positive cell counts of the cellular markers per disease group (CVID, sarcoidosis, PS or TB), normalized towards the total cells detected in that region (granuloma or surrounding). B-D) 10x representative immunofluorescence single stains together with DAPI per IF multiplex assay performed, always showing the same biopsy sample per disease of all three IF assays, complemented with the quantified positive cell counts per total cells detected of the indicated regions and markers per disease group, with violin plots showing median and interquartile range; (B) DAPI = dark blue, CD68 = aqua, CD11c = yellow, PD-L1 = white, FAPα = red, SMA = green; (C) DAPI = dark blue, CD68 = aqua, CD163 = red, pERK1/2 = white, MPO = yellow, FAPa = green; (D) DAPI = dark blue, CD3 = red, CD4 = yellow, FOXP3 = aqua, CD20 = green, PD-1 = white. Statistical analysis with 2way anova, with Tukey’s multiple comparisons test, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.001
Consistent with the digital spatial protein analysis, CD11c + and PD-L1 + cells were mainly localized inside the granulomas (Fig. 5A and B), and were significantly more abundant there than in the surrounding tissue for all diseases (Fig. 5B). FAPα + cells were mainly observed in the granulomas of all diseases groups (Fig. 5A-C), which corresponded with the trend of increased FAPα expression we observed in the spatial analysis (Fig. 2C). This difference however, was only significant for sarcoidosis in the two multiplex assays containing FAPα (Fig. 5B and C). Interestingly, we observed two different staining patterns for FAPα, that were neither disease nor organ specific; in some biopsies FAPα + cells were observed inside the granulomas, while in other biopsies FAPα + cells formed a ring surrounding the granulomas (Fig. 5B and C, Figure S3).
As observed before (Fig. 2D), SMA was increased in the surrounding tissue of sarcoid and TB granulomas compared to CVID and PS, with the majority of observed SMA + cells located around vessel structures (Fig. 5B).
In all disease groups, the number of CD163 + cells was higher in the surrounding than inside the granulomas, with the CD163 + cells inside the CVID and PS granulomas even more abundant than in the sarcoid or TB granulomas (Fig. 5C). Although MAPK-associated proteins were enriched in sarcoidosis versus CVID (Figs. 2 and 3, Figure S2), no significant differences of phospho-ERK1/2 were detected between CVID and sarcoidosis (Fig. 5C). Only a significant increase of phospho-ERK1/2 + cells in sarcoid granulomas versus their surroundings was detected (Fig. 5C).
Since we observed a significant enrichment of CD66b in granulomas of CVID compared to PS, we wanted to investigate this further (Fig. 3). Therefore, we included myeloperoxidase (MPO) as a neutrophil marker. In all disease but CVID, MPO + cell counts were higher in the surroundings versus the granulomas. In CVID granulomas, MPO + cells were clearly increased compared to the granulomas of the other diseases, being significant compared to sarcoidosis and PS (Fig. 5A and C). Similar to the distribution of CD163 + cell counts, the MPO + cell counts in the granulomas and their surroundings were comparable for CVID (Fig. 5B and C).
The IF multiplex assays corresponded with the observations from the spatial protein analysis for CD11c, PDL1, CD163, CD66b and SMA. Compared to the other granulomatous diseases, CVID granulomas contained more CD163 + and MPO + cells within the granulomas as well as in the adjacent surrounding, suggesting that neutrophils and M2-like macrophages might contribute to the pathogenesis of CVID granulomas.
CVID Granulomas and Their Surrounding Areas show a Different T- and B-Cell DistributionConsidering the importance of lymphoid cells in granuloma formation, we included CD3, CD4, FOXP3 (expressed by T-regulatory cells), CD20 (expressed by B cells) and PD-1 (Programmed Death receptor 1; innate and adaptive inhibitor, expressed on activated T- and B cells, DCs and macrophages) into the IF multiplex analysis (Fig. 5A and D). The IF stains clearly showed presence of CD3 + cells (which were mainly CD4+) in CVID granulomas, as opposed to the granulomas of sarcoidosis, PS and TB (Fig. 5D and Figure S3). In TB, CD3 + and CD4 + cells formed a ring-like structure surrounding the granulomas (Fig. 5D). Overall, in CVID the CD3 + and CD4 + cell counts were comparable between the granulomas and their surroundings (Fig. 5D). The FOXP3 + cell counts were overall low. Especially in the surrounding of TB granulomas, significantly fewer FOXP3 + cells were identified compared to all other diseases (Fig. 5D). Similar to CD3 + cells, CD20 + cells seemed to localize at the border of the granulomas in sarcoidosis, PS and TB granulomas. In CVID granulomas CD20 + cells were abundant, especially compared to sarcoid and TB granulomas (Fig. 5D). Although not significant (except for CVID granulomas versus TB granulomas), the presence of PD-1 + cells seemed more pronounced inside CVID granulomas and their surroundings compared to the other granulomatous diseases (Fig. 5A and D).
Also concerning the lymphoid cells, CVID granulomas seemed less organized. Also the presence of CD3+, CD4 + and CD20 + cells in CVID granulomas could be indicative of a lymphoid inflammatory milieu different from that in sarcoidosis, PS and TB granulomas.
留言 (0)