
記住我





DNA methyltransferase 3 A mutations (DNMT3AMT) are one of the most common aberrations in de novo acute myeloid leukemia (AML; 20–25%) [1,2,3], myelodysplastic syndromes (MDS; about 10%) [4] and clonal hematopoiesis of indeterminate potential [5, 6] and are mostly heterozygous [7]. Decreased DNMT3A function results in genomic hypermethylation with a significant enrichment of promoters of the transcription start sites of genes involved in differentiation [2, 7,8,9]. Multiple DNMT3AMT seem to be associated with shorter event-free survival and overall survival (OS) in AML [10] but little is known about the landscape of biallelic DNMT3AMT.
We analyzed 5,603 consecutive cases with myeloid neoplasia (MN) followed from 2002 to 2023 at The Cleveland Clinic for the presence of DNMT3AMT using NGS to determine the genetic and clinical landscape of biallelic DNMT3AMT. Clinical features of total patients subjected to NGS are summarized in Table S1.
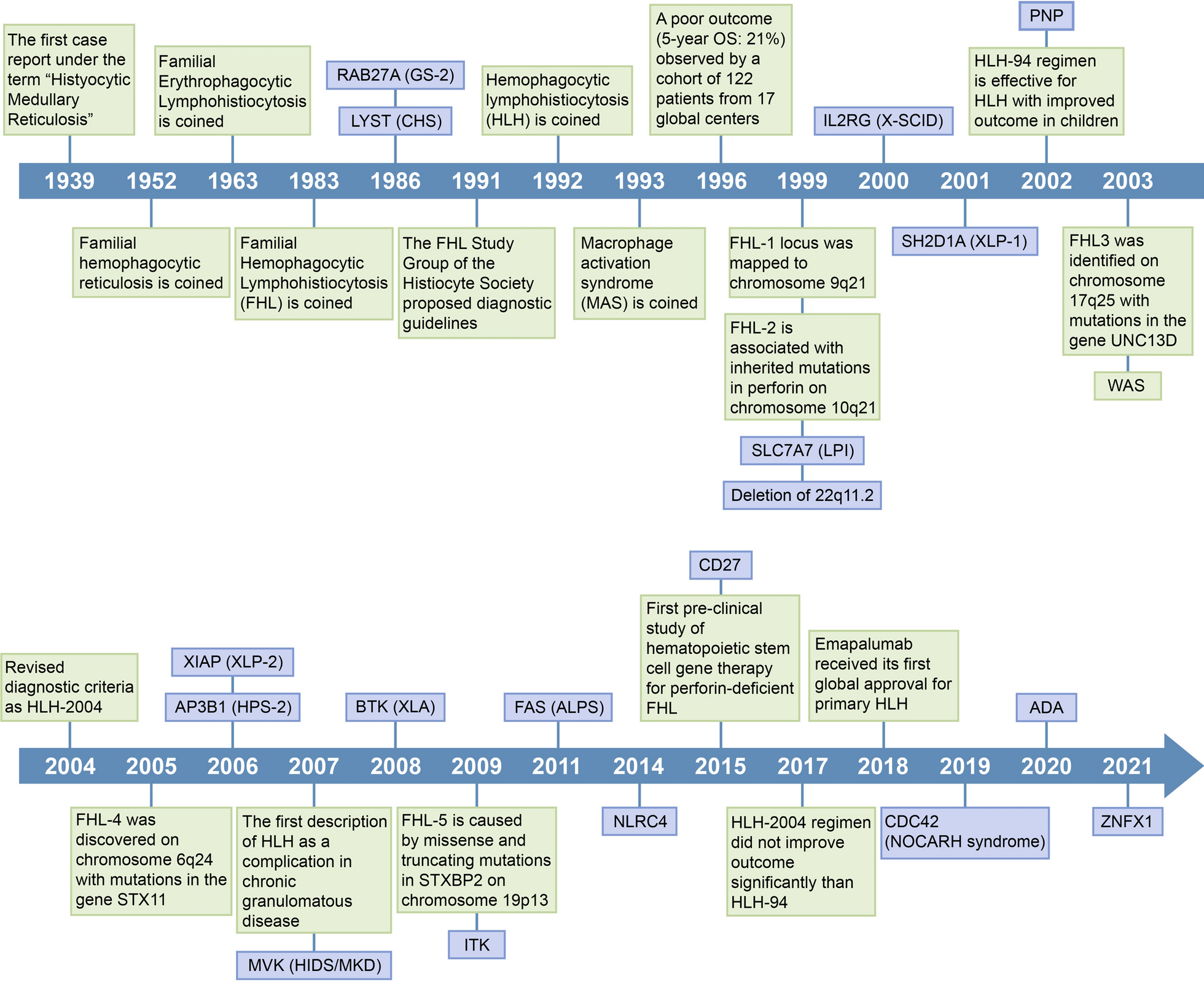
DNMT3AMT were identified in 533 patients (9.5%), of whom 488 carried single and 45 carried multiple hits (2 hits in 44 cases and 3 hits in 1 case; Fig. 1A; Supplementary Tables S2-S4). When compared with single-hit, multiple DNMT3A hits were not found in myeloproliferative neoplasm (MPN), but were encountered more often in AML than in MDS and MDS/MPN overlap (62.2% vs. 33.3% vs. 2.2%; P = 0.027). However, no significant differences in the percentage of blasts or routine blood parameters were found according to the number of DNMT3A hits (Supplementary Table S5). Patients harboring multiple DNMT3AMT carried more likely IDH2 (24.4% vs. 9.6%; OR 3.03; P = 0.005) and ETV6 (6.7% vs. 1.4%; OR 4.91; P = 0.044) mutations compared to those with single DNMT3AMT (Fig. 1B-D).
Fig. 1
Landscape of single and multiple DNMT3A mutations. (A) Lollipop plot of somatic DNMT3A mutations. Variants identified as single and multiple hits are plotted at the top and bottom. (B) Oncoplot of pathogenic mutations identified in patients with single or multiple DNMT3A mutations. (C) Comparison of frequencies of recurrent somatic mutations in patients with single vs. multiple DNMT3A mutations. (D) Odds ratios and 95% confidential intervals of genes more likely to have single (blue) or multiple (red) DNMT3A mutations. * P < 0.05, ** P < 0.01, where P is based on Fisher’s exact test. (E) Scatter plot of the variant allele frequencies (VAFs) of patients with DNMT3AMT. The VAF of first hit DNMT3AMT was plotted on the x-axis and that of the second –hit DNMT3AMT on the y-axis. Patients were categorized into 5 groups by the sum of VAFs. (F) Percentage of different types of monoallelic (upper) and biallelic (lower) DNMT3AMT. (G) Overall survival (OS) comparison between monoallelic (blue) vs. biallelic (red) DNMT3A mutants in DNMT3AMT MDS and AML patients. Kaplan-Meier Curves were compared by Log-Rank test
Next, we analyzed diverse DNMT3AMT configurations based on the sum of VAFs (Supplementary Methods; Fig. 1E). DNMT3AMT were classified into 5 groups, monoallelic (n = 265), hemi/homozygous (n = 24), biallelic (n = 28), bi-clonal (n = 10) or “undetermined” whether biallelic or bi-clonal (n = 7) or whether monoallelic or hemi/homozygous (n = 199). We identified a significant positive correlation between VAFs of two DNMT3A variants (Spearman r2 = 0.6060; P < 0.0001), suggesting that these hits occurred simultaneously. 52 MNs with biallelic or hemi/homozygous DNMT3AMT were grouped as biallelic “DNMT3A impairments” which had inactivation of both parental copies.
Monoallelic hits were missense (73%) including R882 (35%), whereas biallelic hits had a constellation of missense/missense (39%), missense/frameshifts (25%), missense/nonsense (11%), missense/splice sites (11%), missense/in-frame ins/del (7%), frameshift/nonsense (3.5%) and frameshift/frameshift (3.5%; Fig. 1F). R882 accounted for only 13% of biallelic hits, indicating that biallelicity was confined also to non-R882.
Median age was higher in biallelic than monoallelic (73 vs. 69 years; P = 0.029; Supplementary Table S4) and biallelic DNMT3A impairments were not found in MPN (P < 0.001), but more often in AML (72%) than MDS (24%) and MDS/MPN overlap (6%). Patients with biallelic DNMT3A impairment presented higher WBC in both AML (median 16.06 vs. 2.99 × 103/µL, P < 0.0001) and MDS (6.58 vs. 3.02 × 103/µL, P = 0.0032) and higher ANC (1.66 vs. 0.63 × 103/µL, P = 0.0042) and blasts (39% vs. 11%, P = 0.0001) in AML (Supplementary Table S5).
With a median follow-up of 22 months (0.1–219.0), OS rates according to DNMT3A impairment were compared in patients with MDS (n = 111) and newly diagnosed AML who received chemotherapy (n = 96; Supplementary Table S6). Those with biallelic DNMT3A impairment showed significantly poorer 2-year OS than those with monoallelic DNMT3A mutant AML (5.6% [95%CI, 0.4–22.4%] vs. 47.6% [95%CI, 34.9–59.2%]; P = 0.002; Fig. 1G). Although there was no statistically significant difference, the same trend was observed in MDS patients (45.5% [95%CI, 16.7–70.7%] vs. 60.6% [95%CI, 48.9–70.5%]; P = 0.086; Fig. 1G) Both biallelic and hemi/homozygous DNMT3AMT showed inferior OS compared to monoallelic DNMT3AMT (P = 0.007; P = 0.012), while there was no significant difference between biallelic and hemi/homozygous in AML (P = 0.790; Supplementary Fig. S1). Biallelic DNMT3A impairment (HR 2.65; P = 0.001), male gender (HR 2.05; P = 0.014) and adverse genetic alteration according to the European LeukemiaNet 2022 classification (HR1.84; P = 0.028) were deemed as independent adverse factors, while 7 + 3 based intensive chemotherapy (HR 0.47, P = 0.011) as favorable for OS in AML (multivariate analysis; Supplementary Table S7). In MDS, high IPSS risk (HR 2.63, P = 0.001) was the only risk factor for survival, while biallelic DNMT3A impairment was not a significant risk factor (multivariate analysis; Supplementary Table S7).
Since the rank of DNMT3AMT within the clonal hierarchy can be approximated using VAF for clinical purposes, we classified co-occurring DNMT3AMT into ancestral, simultaneous, and secondary mutations (Supplementary Methods; Supplementary Table S8). Among 68 variants co-occurring with biallelic DNMT3AMT, 54% were simultaneous to DNMT3AMT, followed by secondary (37%) and ancestral hits (9%). Ancestral mutations frequently occurred in chromatin regulators (BCORL1, ASXL1, EZH2) with EZH2 mutations identified as earlier events, whereas CUX1, PTPN11 and CEBPA mutations were found only as subclonalMT (Fig. 2A-C).
Fig. 2
Clonal architecture of biallelic DNMT3AMTcases. (A) Scatter plot shows variant allele frequencies (VAFs) of co-occurring mutations (y-axis) and 2nd hit DNMT3A mutations (x-axis) in biallelic DNMT3A mutant cases. (B) VAFs were used to categorize co-occurring mutations into ancestral, simultaneous, or secondary mutations compared with biallelic DNMT3A mutations. The bar graphs show the percentages of the corresponding genes. (C) The prevalence of patients in each mutational type of co-occurring mutations. (D-G) Fish plots of representative longitudinal clonal hierarchy of biallelic DNMT3AMT cases are shown. Clonal architecture of AML at diagnosis and following relapse after conventional intensive chemotherapy-induced remission (D) and allo-HSCT (E). (F) AML-MRC at diagnosis and after refractoriness to the initial therapy. (G) MDS at onset and upon progression to secondary AML (sAML). The variant allele frequencies (VAFs) of each variant are shown at the bottom
In 12 cases with biallelic DNMT3AMT, the VAFs for both DNMT3A variants tracked together with longitudinal NGS results. (Fig. 2D-G; Supplementary Fig. S2). Except for 1 case in CR post allo-HSCT and 1 donor-derived leukemia (Supplementary Fig. S2A-B), biallelic DNMT3AMT clones persisted/ expanded in 9/12 cases (Fig. 2D-G; Supplementary Fig. S2C-G) or was newly acquired at relapse post allo-HSCT in 1 case (Supplementary Fig. S2H). Furthermore, mono-allelic DNMT3AMT clones were lost and only biallelic DNMT3AMT clones dominated at relapse in 2 cases (Fig. 2E and Supplementary Fig. S2C), in which only mono-allelic DNMT3AMT clones persisted during CR (Supplementary Fig. S2C).
Our study describes the role of multiple DNMT3AMT in AML development and persistence, highlighting the likelihood that biallelic DNMT3AMT, while rare, are indeed compatible with clonal expansion and that the acquisition of a second DNMT3AMT may confer a stronger leukemogenic drive accelerating clonal progression. Given the lack of sample availability, our study is limited to estimation of DNMT3A mutational configuration using VAF-method. Further studies using single-cell DNA sequencing are warranted.
留言 (0)