The focus of this article is on differences in short-term (MAs) and medium-term (Hb adducts) biomarkers of acrylamide resulting from different nutritional habits (omnivore, vegan, raw food). Due to the strict avoidance of the consumption of any warmed/heated food for at least four months (Abraham et al. 2022), the raw food eater study allows the monitoring of internal exposure to heat-induced contaminants with respect to their endogenous formation. The suitability of the study design to answer the question raised is, for example, demonstrated by the result that the urinary levels of 3-monochloro-1,2-propanediol (3-MCPD), a contaminant in processed oils and fats (Abraham et al. 2021), were under the limit of quantification in all 16 raw food eaters (article in preparation).
The RDVD study also included smoking omnivores and vegans. Their results confirm the well-known higher exposure via cigarette smoke, which were considered only for some special questions.
Difference in internal acrylamide exposure between vegans and omnivores
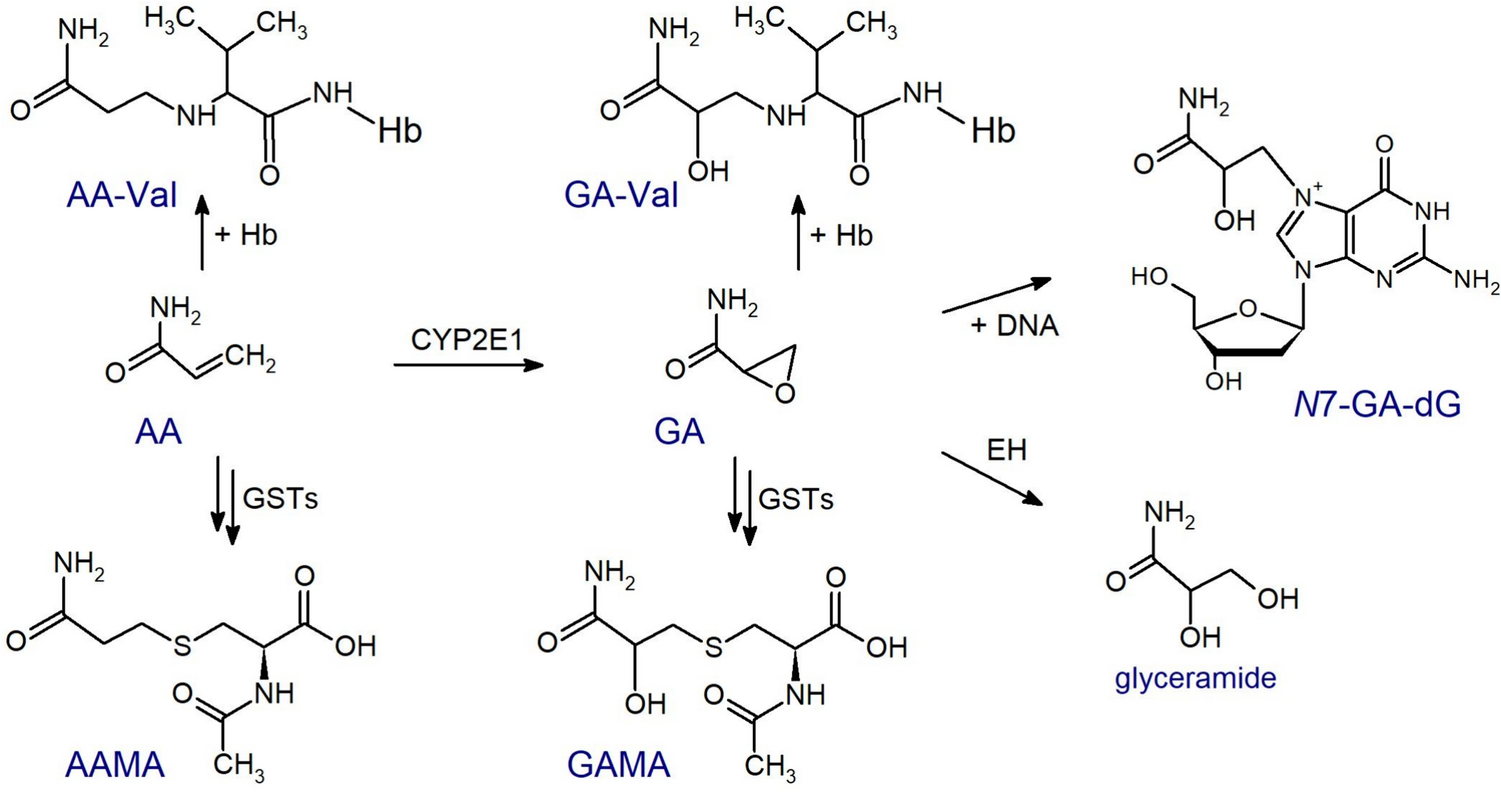
Measurements of the arylamide biomarker—the Hb adducts AA-Val and GA-Val as well as the urinary MAs AAMA and GAMA—largely confirmed the levels and ratios measured by other groups of the general population. With respect to comparison of omnivores and vegans, Goerke et al. (2019) studied the exposure comparing the intake difference in small groups of non-smoking vegans and omnivores (n = 10 with 5 females each) in a duplicate study, analyzing the acrylamide content of all foodstuffs consumed over 10 days and the daily urinary excretion of AAMA and GAMA. Vegan participants ingested more acrylamide (mean 25.2 µg/day) compared to omnivores (mean 17.1 µg/day), and had a higher mean AAMA and GAMA excretion (87.6 µg/day and 12.5 µg/day, respectively) compared to omnivores (58.8 µg/day and 10.0 µg/day, respectively). These results were confirmed by our data revealing that median excretions of AAMA and GAMA in 24-h urine of the non-smoking vegans were higher than the corresponding values in the non-smoking omnivores. Likewise, our data on levels of the Hb adducts AA-Val and GA-Val levels (43% and 30% higher in vegans compared omnivores, respectively) confirmed a higher exposure to acrylamide from food consumption in vegans.
In the duplicate study by Goerke et al. (2019), the consumption of pan-fried vegetables, meat surrogates like tofu or seitan as well as bread-based products appeared to substantially contribute to the higher intake of acrylamide in vegans. In this context, results of the German food monitoring program are interesting, revealing significant levels of acrylamide in rice waffles, hash browns, potato pancakes, and vegetable chips (BVL 2021). Furthermore, the BfR MEAL Study (“meals for exposure assessment and analysis of food”, the first German Total Diet Study) very recently published the data on acrylamide, with highest levels in vegetable crisps (1430 µg/kg), followed by potato pancakes (558 μg/kg) and pan-fried potatoes (450 μg/kg) (Perestrelo et al. 2024). As vegans have to safeguard their energy requirements with a reduced range of meals, a high consumption of these food groups in general is a plausible explanation for the higher internal exposure to acrylamide in vegans compared to omnivores.
Urinary excretion of MAs of acrylamide in raw food eaters
Based on the urinary excretion data observed in rats (levels of AAMA and GAMA in untreated animals similar to those of 14C-AAMA and 14C-GAMA after application of a single oral dose of 0.1 µg 14C-acrylamide/kg bw (Watzek et al. 2012)) and in humans after a washout phase of three to five days with an acrylamide-minimized diet (Goempel et al. 2017; Ruenz et al. 2016), an endogenous formation of acrylamide was hypothesized. In the latter studies, a significant mean urinary AAMA excretion between 0.09 and 0.14 µmol/day was observed after the washout phases. This is somewhat higher compared to the median AAMA excretion of 0.066 µmol/day (15.4 µg/day) in the raw food eaters. The difference may be due to the fact that the acrylamide-minimized diets do not reduce the amount of acrylamide in food that consequently compared to food not heated to higher temperatures than 42 °C. Compared to the non-smoking RBVD omnivores, the median AAMA excretion of the raw food eaters was about 25% (Table 1). This is comparable to the values of 32% and 27% calculated from the mean washout data of Goempel et al. (2017) for group A (washout 9 days) and group B (washout 13 days), respectively, in comparison to the mean AAMA excretions of the respective groups on day 1 (n = 6 omnivores each). These aggregated results would suggest that roughly one-fourth to one-third of the AAMA excretion in omnivores may occur independently of the dietary acrylamide intake.
As an attempt to express this endogenous acrylamide dose as a corresponding daily external dose, reverse dosimetry was applied in the past, e.g., using the data of Goempel et al. (2017) who determined a fraction of urinary 3C3D3-AAMA excretion of 41% (on a molecular base) four days after oral application of an aqueous solution with a dose of 1 µg 13C3D3-acrylamide in six adults per kg bw. As a result, estimations of the corresponding daily external doses between 0.2 and 0.4 µg/kg bw have been published (Goempel et al. 2017; Ruenz et al. 2016). However, these calculations not only assume a bioavailability of 100% [as realistic for aqueous solutions, but may be lower in case of solid foods Berger et al. 2011; Doerge et al. 2005b)], but also a metabolic fate after oral administration comparable to that of endogenous acrylamide (see discussion below).
Hb adducts of acrylamide in raw food eaters
Due to the strict avoidance of the consumption of any warmed/heated foods in the raw food eaters for at least four months, our study allowed to answer questions of possible endogenous formation of heat-induced contaminants not only using short-term biomarkers like mercapturic acids, but also using Hb adducts as more stable medium-term biomarkers. As mentioned above, the median AAMA excretion of the raw food eaters was about 25% compared that of non-smoking RBVD omnivores. In contrast, a ratio of 48% was calculated from the median levels of Hb AA-Val in raw food eaters and those of non-smoking RBVD omnivores. These data of the raw food eaters are the only available, no other data on AA-Val in people strictly avoiding dietary acrylamide exposure for at least four months (the life time of Hb adducts) have been published. Our data indicate an estimated systemic acrylamide exposure in raw food eaters nearly half as high as that of non-smoking omnivores. The difference in biomarker results for urinary AAMA excretion (25%) and for AA-Val (48%) is also reflected in the ratios of AAMA and AA-Val as well as of GAMA and GA-Val (Fig. 5), showing significantly higher ratios in the RBVD participants compared to the raw food eaters.
At first glance, this seems surprising, but may indicate a different metabolic fate of dietary (oral) acrylamide and endogenous acrylamide. Notably, after oral exposure and intestinal absorption, acrylamide undergoes hepatic first-pass metabolism, leading to significant turnover of acrylamide to glycidamide and to detoxification of acrylamide through glutathione conjugation and other pathways. This has been demonstrated in rodents (Doerge et al. 2005a, b), resulting in different internal exposure of acrylamide and glycidamide upon oral uptake in comparison to intravenous administration. The latter reflects the situation in case of an endogenous source of acrylamide. Humans are less competent to metabolize acrylamide to glycidamide than rodents, but detoxifying biotransformations, especially coupling of acrylamide and glycidamide to glutathione, are more efficient in humans than in rodents (Berger et al. 2011; Fennell and Friedman 2005; Fuhr et al. 2006; Rietjens et al. 2022). In view of our data, a relevant first-pass metabolism with relatively high glutathione conjugation is plausible and would explain the differences between the RBVD participants (vegan or omnivore nutrition) and the raw food eaters observed in our studies. Thus, a higher proportion of AAMA (and GAMA) was formed from acrylamide after oral exposure compared to acrylamide from endogenous sources. Therefore, the use of a fraction of urinary AAMA excretion determined after oral acrylamide administration (e.g., 41% on a molecular base by Goempel et al. (2017), see above) for reverse dosimetry may provide an underestimation of the corresponding external dose of acrylamide, if this re-calculation is done with urinary AAMA data from raw food eaters or people with minimized acrylamide intake for several days. In these cases, a fraction of urinary AAMA excretion determined following an intravenous application would be more appropriate, but is not available.
A generally accepted model for reverse dosimetry using the AA-Val values (as medium-term biomarkers reflecting the systemic exposure) is not available, but an estimation is possible using data of the study in humans by Fennel et al. (2005). They applied single oral doses (0.5, 1.0, or 3.0 mg/kg bw in aqueous solutions) of 13C-acrylamide to male participants (n = 5 per group), and measured 13C-AA-Val and 13C-GA-Val levels linearly increasing with the dose. On average, one mmol 13C-acrylamide per kg bw led to mean 13C-AA-Val levels of 74.7 nmol per g globin. Using this value and considerations for reverse dosimetryFootnote 2 (Fennell et al. 1992; Hays and Aylward 2008), an external dose of 0.22 µg/kg bw was calculated from the median AA-Val level in the raw food eaters of 13.3 pmol/g Hb. A dose of acrylamide in this range representing endogenous exposure would add to the external dose of dietary acrylamide, which was estimated to be 0.4 µg/kg bw per day on average in German adults of the general population (EFSA 2015).
Finally, a small note in connection with the hepatic first-pass effect: It may be confusing that smokers of the RBVD study have about the same ratios of AAMA/AA-Val as the non-smokers (Fig. 5), as the former have a relevant acrylamide exposure from inhalation of cigarette smoke not undergoing a hepatic first-pass effect. However, data in rats (Doerge et al. 2005b; Sumner et al. 2003) may indicate a pulmonary first-pass metabolism after inhalation similar to the hepatic first-pass metabolism after oral exposure (IRIS 2010).
Possible sources of internal acrylamide exposure in raw food eaters
Throughout this article, the term “endogenous” was used to describe the source of the internal exposure to acrylamide in raw food eaters, knowing that this is a simplification. Firstly, an exposure via the dermal route (e.g., by hair and skin care products (Kraeling and Bronaugh 2005)) may theoretically be possible, but evidence for a relevant exposure in the general population is missing. Furthermore, dermal bioavailability was found to be low (Fennell et al. 2005). Secondly, inhalational exposure from second-hand tobacco smoke and smoke from barbecuing and campfires (Goerke et al. 2019) is possible, but all raw food eaters were asked and denied to be exposed to this kind of smoke. Thirdly, the foodstuffs consumed by raw food eaters may contain acrylamide, even if not heated at temperatures > 120 °C. Dried fruits are known to possibly contain acrylamide without such a heat-treatment. Surma et al. (2018) reported acrylamide levels in dates (3 out of 4 brands; 20.1 to 50.9 µg/kg), in apricots (5 out of 6 brands; 13.5 to 100 µg/kg), and in plums (6 out of 6 brands; 22.1 to 141 µg/kg). The mechanism(s) of acrylamide formation during drying are not known, although two of the required conditions are met: the fruits mentioned contain high levels of the precursor asparagine (Amrein et al. 2007; Bahrami et al. 2021; Lo Voi et al. 1995), and the decreasing moisture content favors the Maillard reaction and the formation of acrylamide (Lund and Ray 2017). Amrein et al. (2007) commented that the drying process may take several days entailing temperatures reaching 70 to 80 °C, which is probably not known to all raw food eaters. Using the three-day weighed food records, the consumption of dried fruits was documented and revealed no consumption in 7 participants, a moderate consumption in 8 participants (average 70 g/day, range 10 to 179 g/day), and an extreme consumption (797 g/day) in one participant. The latter was the one with an extreme urinary AAMA excretion (but unremarkable Hb AA-Val levels), and a later analysis of “Medjool” dried dates revealed a high level of acrylamide (see results chapter; urinary AAMA excretion of this participant was replaced the results by a new 24-h urine collected on a day without prior consumption of dates). Urinary AAMA excretions of raw food eaters having consumed dried fruits during the 3-day food recordings were not significantly different from those of the non-consumers. Therefore, we would exclude a relevant contribution of acrylamide from dietary exposure in our group of raw food eaters, and consider their internal exposure to acrylamide as a result of endogenous formation.
The causes of such an endogenous formation of acrylamide in the organism is under scientific discussion. Oxidative stress is one of the hypotheses. Tareke et al. (2008) proved this hypothesis by feeding mice with substances known to induce formation of reactive oxygen species, and observed an increase of AA-Val and GA-Val levels. The picture was completed by demonstrating that acrylamide is formed at physiological conditions (37 °C, pH 7.4), when asparagine is incubated in the presence of hydrogen peroxide (Tareke et al. 2009). Rietjens et al. (2022) hypothesized that the addition of ammonia to acrolein may lead to the formation of acrylamide, and generation in the gut may also play a role. In humans, a positive association between the urinary excretions of AAMA and the biomarker for oxidative stress, 8-hydroxy-2´-deoxyguanosine, was observed in adolescents and young adults (n = 800) (Lin et al. 2013). According to the literature evaluation of EFSA (EFSA 2015), various parameters, i.e. BMI, alcohol consumption, sex, age or metabolic genotype, may have an impact on the urinary excretion of AAMA and GAMA and the formation of AA-Val and GA-Val levels in Hb.
Time trend of acrylamide exposure between 2017 and 2021 in the RBVD study
Since the discovery of relevant dietary expose to acrylamide in 2002, mitigation strategies (current regulation in the European Union: 2017/2158) led to reduction of acrylamide levels in some food classes like potato crisps, but for other food categories, no stable trends of acrylamide were observed in Europe (EFSA 2015; Mojska and Gielecińska 2024). With respect to the biomarker levels of our study, we observed significant increases of AA-Val and GA-Val in non-smoking omnivores (26% each) and in vegans (6.5% and 9.3%, respectively). These results do not support that mitigation strategies in recent years are successful. Data on urinary AAMA and GAMA excretion showed a slight decrease in vegans only. However, far-reaching conclusions should be avoided as the numbers of participants were relatively low.
Validity of AAMA/GAMA and AA-Val/GA-Val as biomarkers of individual exposure
The interpretation of our data goes hand in hand with a critical consideration about the validity and scope of the biomarkers. Daily excreted amounts of MAs are short-term biomarkers of exposure to the parent compounds; however, the individual short-term variability of exposure usually is unknown and depends on the relative constancy of the dietary habits. In this regard, Hb adducts provide a higher reliability because they represent an integrated mean level of plasma concentrations over a few months, and thus are indifferent to daily extremes of exposure (Neumann 1984; Wild 2009). Using the RBVD data of 39 participants from 2017 and 2021, we evaluated the intra-individual stability over time, a particular requirement in order to study associations between exposure and slowly evolving adverse health effects like cancer (Barregard et al. 2013; Jungert and Frank 2021). As expected, correlations of individual levels (Fig. 8) were found to be weak for urinary AAMA and GAMA excretion. These results as well as the corresponding ICCs underline that a single measurement of the daily excretion of the MAs does not adequately reflect the individual exposure at a certain time. The corresponding results of the pairs of AA-Val and GA-Val levels determined four years apart demonstrated much better correlations and higher ICCs, confirming the significance of Hb adducts as biomarkers of exposure.
In this context, the correlations between the excreted amounts of AAMA and GAMA and the respective medium-term biomarkers AA-Val and GA-Val observed in our study in non-smoking participants of the RBVD study (Fig. S2 in the Supplemental Information, data of 2017) were moderate only (rS = 0.50 and 0.58, respectively), emphasizing a high day-to-day variability and the limited capability of the MAs to replace Hb adducts as biomarkers of exposure. However, in epidemiological studies on acrylamide exposure (e.g. Poteser et al. (2022)), often only urinary AAMA and GAMA are measured in spot urine (which is available more easily than 24-h urine), and the results often are given as concentration (µg/L), but were not based on urinary creatinine levels to adjust for the hydration status [which, however, may be misleading for the comparison of vegans and omnivores (Abraham et al. 2023)]. Such data may only be applicable to compare acrylamide exposure in different populations. Compared to AAMA and GAMA, the Hb adducts AA-Val and GA-Val reflect the systemic exposure and are superior biomarkers due to their time stability and significance. For better reverse dosimetry, a controlled exposure study in humans would be needed to establish a reliable adduct level increase per dose factor, allowing the estimation the average external acrylamide exposure from AA-Val, as shown previously for glycidol and its Hb adduct N-(2,3-dihydroxypropyl)-Val (Abraham et al. 2019).






留言 (0)