Ovarian TCs represent a relatively understudied aspect within the female reproductive system, with no prior investigation reported in rat ovaries to the best of our knowledge. A study conducted by Liu et al. (2016) explored TCs in the ovarian stroma of mice, utilizing electron microscopy, immunofluorescence, immunohistochemistry and flow cytometry by employing some markers such as CD34, vimentin and PDGFR-α and -β. The researchers found these cells characterized by a small body and a moniliform transitioning arrangement of branching Tps. Furthermore, it was suggested that TCs may play a role in sustaining the local microenvironment.
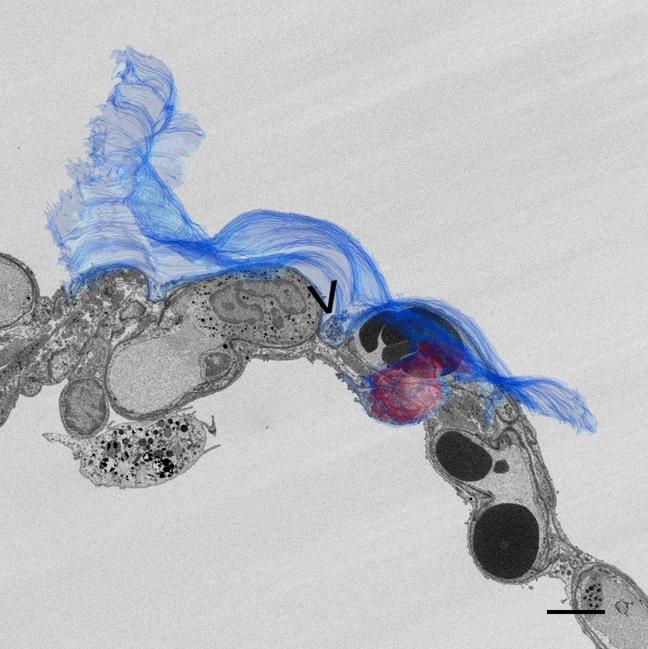
In our study, we have successfully demonstrated, for the first time, the presence of TCs in rat ovaries. Moreover, our investigation revealed a widespread distribution of TCs throughout the ovarian tissue. The observed morphology of TCs in the ovary closely resembled that reported in various other studies (Liu et al. 2016; Mazzoni et al. 2019; Mohamedien et al. 2023). Importantly, we observed an abundance of Tps and Pdms throughout the ovarian stroma, indicating a close interrelation among these structures and with other stromal cells. This supports the notion that TCs may form intricate three-dimensional networks via homo- and heterocellular connections, as previously proposed by Cretoiu and Popescu (2014b). Notably, our findings also align with existing data, as TCs in the rat ovary exhibited Tps densely populated with organelles, including RER, caveolae, Golgi apparatus, ribosomes, endocytic vesicles, lysosomes and autophagic figures. The presence of these organelles within the Tps underscores the high metabolic activity of these cells (Lemons et al. 2010; Felisbino et al. 2019). Furthermore, consistent with prior knowledge, Tps are recognized for their ability to secrete three forms of extracellular vesicles which are exosomes, ectosomes and multi-vesicular cargos implicated in paracrine signalling (Cretoiu and Popescu 2014b, a). In this study, we report that the nuclei of TCs in both the ovary and uterus consist of centrally euchromatic regions with peripheral heterochromatin. However, in various studies, there is a debate regarding the dominant presence of heterochromatin in the nuclei of TCs. Similar to our findings, some researchers propose that TCs exhibit nuclei with irregular indentations, central euchromatin and heterochromatin concentrated near the nuclear membrane. Studies also assert that TCs typically feature nuclei characterized by thin heterochromatin band and infrequently observable nucleoli (Cretoiu et al. 2012; Cretoiu and Popescu 2014b, a; Condrat et al. 2021).
Studies on TCs in fish ovaries have elucidated their role in establishing intricate connections between stromal and vascular components, forming a labyrinth network. These TCs have been shown to engage with immune cells and blood vessel endothelium, shedding extracellular vesicles in close proximity to blood capillaries, and contributing to tissue regeneration during the spawning season (Mokhtar 2019; Mohamedien et al. 2023). Additionally, research on rabbit ovaries has proposed a potential involvement of TCs in the formation and regeneration of theca interna, interstitial gland cells and theca lutein cells (Abd-Elkareem 2017).
Consistent with these findings in various species, our study observed the presence of TCs, along with their Tps, situated between the connective tissue of the theca layers. Interestingly, these TCs did not establish direct contact with granulosa cells. In contrast, TCs with their Tps and Pdms were specifically localized around the corpus luteum, in close proximity to luteinized granulosa cells as well as with other interstitial cells. Moreover, TCs within the stroma exhibited a consistent positioning around blood vessels, particularly in close proximity to pericytes and smooth muscle cells, aligning with the observed patterns in previous studies across different species as mentioned in other studies (Mokhtar 2019; Mohamedien et al. 2023).
On the contrary, our examination of uterine TCs revealed their presence in all three layers, namely the endometrium, myometrium and perimetrium. A study conducted by Salama (2013) has already demonstrated the existence of TCs in the endometrium and myometrium of the rat uterus across various reproductive stages. The suggested roles of these TCs include providing support to glands and facilitating stromal cell communication in the endometrium. In the myometrium, their proposed functions involve initiating and coordinating myometrial contractions.
In parallel with the observations in the ovaries, our investigation reveals that Tps and Pdms form an extensive meshwork throughout the stroma of all three layers of the uterus. Tps in the rat uterus exhibit variable distances when connecting to other cells, suggesting a range of cell-to-cell communication. Additionally, Tps are known to establish connections with each other through end-to-end or side-to-side contacts, but rarely through end-to-side connections in the uterus. These morphological observations provide evidence supporting the notion that Tps may not only convey signals or engage in unique communication with other TCs but also with neighbouring stromal cells (Hatta et al. 2012; Yang et al. 2016). The protrusions of the cytoskeleton driven from a stromal cell to a Tp seem to be tunnelling nanotubes or cytonemes. These types of communications are known to allow the transfer of different cargoes, ranging from ions to organelles (Abounit and Zurzolo 2012; Gerdes et al. 2013; Korenkova et al. 2020). We also noted that the Tps, especially their Pdms in the uterine tissue were abundant in organelles, displaying a similar feature observed in TCs in the rat ovary.
Furthermore, existing literature reports the presence of Tps in the uterus, positioned in close proximity to secretory glands and blood vessels in the lamina propria beneath the simple columnar epithelium. Notably, close connections between TCs and smooth muscle fibres, facilitated by gap junctions and connective tissues, have been observed (Yang et al. 2016). This aligns with observations in the human endometrial stroma, where TCs are present in both the stratum functionalis and basalis, adopting an orientation that contours the shape of the adjacent epithelial architecture (Hatta et al. 2012).
Aligned with these coherent observations, our study reveals that Tps are not confined solely to the vicinity of blood vessels; they were also identified around uterine glands. TCs were further identified in the connective tissue between the smooth muscle layers of the myometrium and in the perimetrium, mirroring the distribution observed in the endometrial stroma. Previous research has proposed that TCs could exert influence on the timing of contractile activity in smooth muscle cells, highlighting their potential key role in uterine contraction (Yang et al. 2016). In connection with this, there is a suggestion that TCs may contribute to a range of physiological functions in both non-pregnant and pregnant states of the human uterus, which is recognized for its myogenic contractions (Roatesi et al. 2015).
We found that TCs were distributed among other resident interstitial cells, including mast cells, indicating a potential involvement in immunological processes in the rat uterus. Jiang et al (2018) investigated the immunomodulatory capabilities of uterine TCs, particularly their influence on macrophages. This study revealed that TCs could directly activate these innate immune cells through the mitochondrial signalling pathway. Although the experiment was conducted on peritoneal macrophages, the findings suggest that similar processes may occur in the uterine microenvironment.
It is known that TC markers such as c-kit and PDGFR-ß (Suciu et al. 2012; Salama 2013; ES 2022), which are used in this study, can also be expressed by other cell types found in connective tissue. Despite some researchers using one of these antibodies to identify TCs based on their morphology, the double staining method is considered more reliable and is commonly employed. Therefore, we used c-kit and PDGFR-β antibodies to identify TCLCs based on their morphological characteristics. Our next objective was to assess semi-quantitatively the presence of TCLCs in both the stroma of the rat ovary and in the endometrium of the rat uterus. This assessment helped us to ascertain the concordance in the quantitiy of TCLCs designated by a single antibody, shedding light on potential correlations between these markers and the previously identified TCLCs phenotype. The non-significant results between the c-kit+ and PDGFR-β+ TCLCs support the conclusion that a significant number of the cells in the ovarian stroma and endometrial stroma of uterus can be classified as TCs, given their positivity for both PDGFR-β and c-kit. Additionally, we supported our single marker analysis with double immunohistochemistry using CD34 (a membrane marker which stains the thin Tps) and α-SMA (cytoplasmic marker). These results strongly suggest that the TCLCs found in the rat ovary and uterus are indeed TCs.
Salama (2013)’s study was also involving healthy female albino rats, divided into immature, adult non-pregnant, pregnant and post-partum groups. They performed immunohistochemical identification of TCs with a c-kit antibody on the middle one-third of the right uterine horns. The number of c-kit-positive telocytes per high-power field has been calculated and statistically examined in both the endometrium and myometrium. TCs that were c-kit-positive were found in the endometrium, specifically around endometrial glands, while in the myometrium they were oriented parallel to circular smooth muscles and longitudinal muscle bundles. It should be noted that the human myometrium comprises two crucial cell populations contributing to its contractility which are smooth muscle fibres and interstitial cells. Although the pace-making mechanism remains unidentified, it is plausible that myometrial smooth muscle cells contract in response to a signal generated by c-kit-positive interstitial cells (Cretoiu et al. 2011). Given that the female reproductive system is highly hormonally regulated, TCs have been shown to exhibit different characteristics and functions during various stages of reproduction in the rat uterus (Salama 2013). For the first time, our study demonstrates the presence of TCs in the rat ovary. Therefore, we believe that investigating TCs in the rat ovary, particularly their roles across different reproductive stages, would be of significant importance for further studies.
As a conclusion, TCs, along with their Tps, Pdms and Pds, were observed throughout the stroma of the rat ovary, closely associated with follicles, corpus luteum, blood vessels, and other stromal cells. Subsequently, a meshwork of rat uterine TCs was identified in the stroma of the endometrium, myometrium and perimetrium, in close proximity to blood vessels and uterine glands. It is essential to investigate further the roles and functions of TCs in the female reproductive system as well as TC related disorders.









留言 (0)