
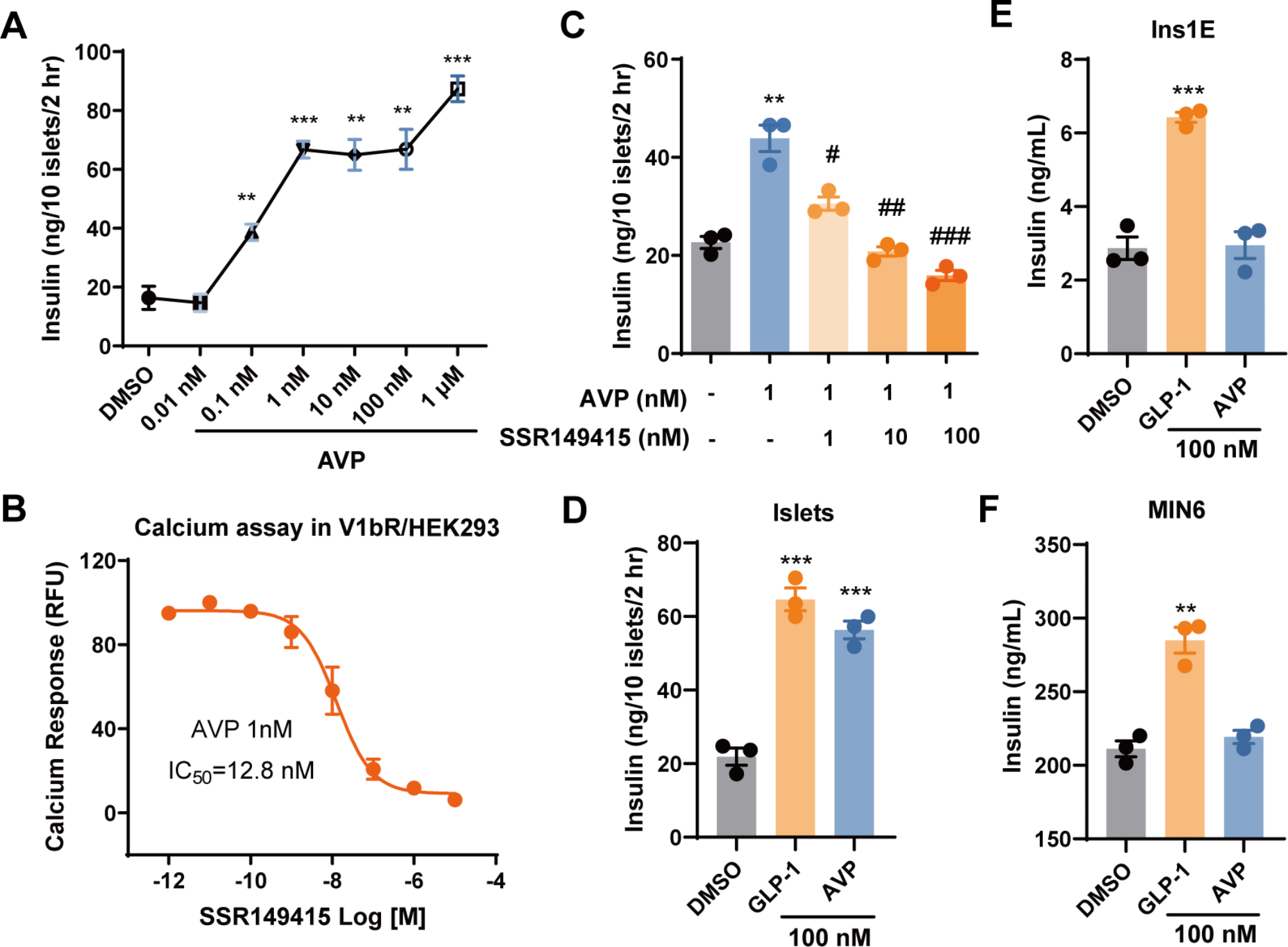
記住我






During the mammalian embryonic development, the paraxial mesoderm, situated bilaterally adjacent to the neural tube, forms during gastrulation. The anterior segment of this mesoderm epithelializes into segmented somites [1]. These somites subsequently compartmentalize along the dorsal–ventral axis into two distinct structures: a dorsal epithelial dermomyotome and a ventral mesenchymal sclerotome. The dermomyotome differentiates into skeletal muscle, brown adipose tissue, and the dermis of the back, whereas the sclerotome gives rise to the axial skeleton and tendons [2] (Fig. 1A).
Fig. 1
Differentiation of UiPSM cells into skeletal myocytes in vitro. A Schematic diagram of somite development. a. Illustration of the epithelial somite's spatial relationship to surrounding structure. b. Depiction of the differentiated somite's spatial relationship to surrounding structures. Dorsally, the somite differentiates into the dermomyotome and sclerotome. The dermomyotome subsequently gives rise to the myotome, which develops into skeletal muscle tissue. The sclerotome differentiates into osteoblasts and chondroblasts, forming the axial skeleton. B Schematic diagram of UiPSM cell differentiation into skeletal myocytes, osteoblasts and chondroblasts. C Schematic overview of stepwise differentiation of skeletal myocytes from UiPSM cells. Representative images show the morphological changes from UiPSM cells to skeletal muscle filaments. Scale bars, 100 µm. D Representative gene expression of human skeletal muscle satellite cells (PAX3, PAX7, CXCR4, C-MET) at day 15. Data are mean ± SD, n = 3 independent experiments. (*P ≤ 0.05). E Immunofluorescence of PAX3 and PAX7 during the differentiation of skeletal muscle cell form UiPSM at day 60 (left). The scale bar represents 100 µm. The values on the left represent the percentage of positive cells statistically. Data are mean ± SD, n = 3 independent experiments, each experiment counted 100 fields of view. F Representative gene expression of human skeletal muscle satellite cells (PAX3, PAX7) and skeletal myoblasts (MYOD, MYOG, MRF4) and skeletal myocytes (MYH3, MYH7) during the differentiation process. Data are mean ± SD, n = 3 independent experiments. (*P ≤ 0.05). G Immunofluorescence of MYOD, MHC, Desmin, Laminin during the differentiation of skeletal muscle cell form UiPSM at day 60 (left). The scale bar represents 100 µm. The following values represent the percentage of positive cells statistically. Data are mean ± SD, n = 3 independent experiments, each experiment counted 100 fields of view. H The UiPSM cells differentiated at day 30 and day 60 were enriched for GO terms of skeletal muscle development. I Heatmap illustrating the gene expression of skeletal muscle development related genes with dramatical change in UiPSM cell derived myocytes at day 30 and day 60
Skeletal muscle, the largest tissue by mass in the body, is crucial for movement and support. However, millions of individuals worldwide are suffering from skeletal muscle atrophy, caused a range of factors, including cachexia, sarcopenia, and muscular dystrophies. The latter factors, encompassing over 30 distinct genetic disorders, often result in paralysis and are frequently associated with cardiopulmonary complications [3]. Currently, treatments are predominantly conservative and lack definitive cures. The replacement of diseased muscle with healthy muscle fibers derived from stem cells offers a promising approach. Prior research has shown that myogenic progenitors, which arise during myogenesis, can be differentiated directly or reprogrammed from embryonic stem cells or pluripotent stem cells (PSCs) [4,5,6,7,8]. However, the incomplete differentiation of PSCs poses a clinical safety risk. The process of in vitro PSC myogenesis necessitates a transition through a mesodermal stage [9]. For therapeutic applications, the generation of abundant, engraftable, tissue-specific cells are required. Establishing a mesodermal progenitor cell lineage with specific differentiation capabilities to expand the pool of transplantable myocytes is an area warranting further investigation.
In our research, we have successfully reprogrammed human urinary epithelial cells into presomitic mesoderm progenitor cells (UiPSMs). These cells are capable of long-term expansion in vitro and differentiate into mesoderm cell types in vivo, displaying anteroposterior axis and segmentation clock features [10]. In this study, we established a UiPSM-based approach to skeletal myogenesis, enhanced by MYOD-mediated maturation. Transplantation of these cells into injured mouse muscle demonstrated sustained regeneration and repair. Additionally, we induced differentiation into osteoblasts and chondrocytes (Fig. 1B). These findings open new pathways for treating musculoskeletal diseases.
Generation of human skeletal muscle satellite cell from UiPSMMyogenesis during development contains two distinct phases: initially, the Paired Box Homeotic Gene (Pax7) and its homolog Pax3 confer myogenic fate in an early embryonic or primary phase [11]; subsequently, the cell fusion and the incorporation of myonuclear from proliferating Pax7+ progenitors facilitate secondary myogenesis [12]. Consequently, we devised a two-stage protocol for differentiating UiPSM cells into skeletal myocytes (Fig. 1C). Previous study had proved that inhibiting endogenous TGF-β (using LDN93189 or SB431542) and stimulating myogenesis-promoting factors such as hepatocyte growth factor (HGF) and insulin-like growth factor 1 (IGF1) are crucial for maximal Pax7 induction [13]. And dexamethasone (DEX) has been shown to improve myogenesis, advances muscle structure, and increases force production in engineering skeletal muscle tissue [14]. Building on this foundation, through high-throughput screening and testing of various small molecule combinations, we found that Vitamin C effectively enhance myogenic differentiation. Thus, for the initial phase, the optimal combination for maximal PAX7 induction from UiPSM cells in 15 days involved activating IGF/HGF alongside Vitamin C and Dexamethasone, and inhibiting TGF-β with SB431542. We termed this medium SM1. The cytokine receptor CXCR4, in tandem with the adaptor protein Gab1, which mediates c-Met signaling, regulates the development of migrating Pax3-positive myogenic progenitor cell [15, 16]. Due to their functional roles, Cxcr4 and c-Met enable the isolation of high-purity PAX3+ PAX7+ skeletal muscle precursors [17]. RT-PCR analysis confirmed that after 15 days of differentiation, UiPSM cells robustly expressed PAX3, PAX7, C-MET, and CXCR4, verifying the formation of PAX3 and PAX7 positive skeletal muscle precursors (Fig. 1D). Immunofluorescence assays revealed 30–50% of cells were PAX3+ or PAX7+, with 30% being PAX3+ PAX7+, indicating muscle lineage commitment (Fig. 1E).
For secondary myogenesis, Pax7+ and Pax3+ myogenic precursors derived from UiPSM cells fused to form larger fibers, expressing more mature MyHC isoforms, like fast MyHC [18]. We transitioned UiPSM cells, after 15 days in SM1 medium, to a medium supplemented with HGF, IGF-1, and horse serum for up to 2 months to promote further differentiation (Fig. 1C). During dermomyotome formation, cells begin downregulating Pax3 and Pax7, concurrently activating muscle regulatory factors (MRFs) including Myf5, MyoD (Myod1), and MRF4 (Myf6) [9, 19]. Our qPCR results showed a decrease in PAX3 and PAX7 expression by day 15, alongside increased expression of MYF5, MYOD, and MRF4 by day 30, suggested the activation of a myogenic transcription factor network (Fig. 1F). Notably, Myf5, transiently expressed in dermomyotome cells, is known to specify the skeletal muscle lineage in mouse and chicken embryos [9, 20]. Its expression peaked on day 30, indicating the onset of myogenesis (Fig. 1F). Myogenin, functioning downstream of Myf5 and Myod, controls the terminal differentiation of myoblasts into myocyte [20, 21]. The concurrent activation of MYOG at day 30 gave supported to the formation of myocytes (Fig. 1F). Afterwards, Myogenin-positive myocytes formed embryonic myosin heavy chain-positive skeletal myocytes, eventually fusing into primary myofibers [22, 23]. The increase in MYH3 and MYH7 transcripts between days 40 and 60 signified terminally differentiated skeletal myocytes/myotubes (Fig. 1F). Terminal differentiation was further evidenced by the expression of additional mature muscle markers, Desmin, Myosin Heavy Chain (MHC), and Laminin, in approximately 10–20% of cells by day 60, as shown by immunocytochemistry (Fig. 1G). RNA-seq data analysis indicated that UiPSM-differentiated cells on days 30 and 60 were closely related to muscle tissue development and muscle contraction, enriching for skeletal muscle (ACTC1, ACTA1, ACTA2), Myosin Light Chain (MYL7, MYL9, MYL4, and MYL12B), and Dystrophin (DMD) genes (Fig. 1H, I), supported a mature myocyte phenotype. Therefore, at this stage, UiPSM-derived PAX3+ and PAX7+ myogenic precursors had differentiated into proliferating MYOD1+ myoblasts, MYOG+ myocytes, and MHC+ myocytes/myotubes.
MYOD increased the maturity of skeletal myocytesGiven the modest efficiency of myogenic conversion in UiPSM cells, these cells might not be effectively activating Muscle Regulatory Factors (MRFs), which are crucial for initiating downstream muscle-specific gene expression. MyoD, a pioneering factor in myogenesis, is known for its ability to convert various cell types, including fibroblasts, pigment cells, nerve cells, adipocytes, and hepatocytes, into skeletal muscle [24, 25]. Prior studies have shown that MyoD-reprogrammed hESCs can efficiently generate myotubes by activating myogenic programs [6, 26]. Consequently, we hypothesized that MYOD might be capable of reprogramming the nuclei of UiPSM cells into a skeletal muscle phenotype.
To test this hypothesis, we evaluated the ability of UiPSM cells to undergo direct myogenic conversion in response to ectopic MYOD expression under consistent culture conditions (Fig. 2A). Remarkably, MYOD expression significantly increased the expression of myocyte-specific genes, such as MYOG, MYH3, and MYH7. The expression of the myoblast gene MRF4 and endogenous MYOD increased approximately tenfold. In contrast, MYOD expression did not notably activate early myogenesis genes PAX3, PAX7, and MYF5 (Fig. 2B, Additional file 1: Fig. S1A). This outcome is consistent with the understanding that MyoD acts downstream of Pax3, Pax7, and Myf5 in the myogenic cascade [22, 27, 28], and MYOD was also shown to be insufficient to induce Pax3-positive cells [23]. Additionally, Myf5 and MyoD are differentially distributed in the epaxial and hypaxial dermomyotome, respectively [27]. These findings suggested that MYOD can effectively induce later stages of myogenesis in UiPSM cells. The efficacy of MYOD in driving terminal myogenesis in UiPSM cells was further substantiated by both FACS and immunocytochemistry. These methods revealed a quick increase in the expression of mature muscle marker proteins—Desmin, Myosin Heavy Chain (MHC), and Laminin.
Fig. 2
MYOD promoted the maturity of skeletal myocytes in vitro. A Schematic overview of stepwise differentiation of skeletal myocytes from UiPSM with ectopic MYOD. SKM: skeletal muscle cells. Representative images show the morphological changes from UiPSM cells to skeletal myocytes. Scale bars, 100 µm. n = 3 independent experiments. B Representative gene expression of human skeletal muscle satellite cells (PAX3, PAX7), skeletal myoblasts (MYOD, MYOG, MRF4) and skeletal myocytes (MYH3, MYH7) when overexpressed ectopic MYOD during the differentiation process. Mcherry as a negative control of overexpression vector. Data are mean ± SD, n = 3 independent experiments. (*P ≤ 0.05). C Detection of skeletal muscle satellite cell-specific genes (PAX3 and PAX7) in MYOD-mediated differentiated UiPSM cells at day 15. The following values indicate the percentage of positive cells statistically (Data are mean ± SD, n = 3 independent experiments). Scale bars, 100 µm. D Flow cytometric analysis evaluating differentiation efficiency via MHC and Desmin protein expression in skeletal muscle cells at day 60 of differentiation. hESC (H9)-derived skeletal muscle cells at day 85 are used as a positive control. E Immunofluorescence analysis of MYOD, MHC, Desmin, and Laminin in UiPSM-derived muscle fibers at day 30 with ectopic MYOD. Scale bar represents 100 µm. The values indicate the percentage of positive cells statistically (Data are mean ± SD, n = 3 independent experiments, each experiment counted 100 fields of view). Scale bars, 100 µm. F MYOD-mediated differentiation of UiPSM cells into skeletal muscle cells at days 15 and 30, showing enrichment for skeletal muscle development-related GO terms. G Heatmap illustrating gene expression changes specific to skeletal myocytes in MYOD-mediated UiPSM cell-derived myocytes at days 15 and 30
Notably, the expression of these markers escalated to approximately 50% by day 30 (Fig. 2D, E). RNA sequencing (RNA-seq) data analysis provided deeper insights into the nature of MYOD-induced myogenesis on days 15 and 30. The data indicated a progression towards more mature muscle features, encompassing aspects such as muscle contraction, sarcomere organization, striated muscle tissue development, muscle filament sliding, myofibril assembly, and muscle fiber development (Fig. 2F). A notable enrichment of skeletal muscle genes, including ACTC1, ACTA1, ACTG2, and ACTN2, was observed. Additionally, the expression of Myosin Light Chain (MYL) genes (MYL4, MYL1, and MYLP) and components of the troponin complex (TNNI1, TNNT2, TNNC2, TNNI3, TNNC1, TNNT1, TNNT3) associated with the sarcomere thin filament in striated muscle was significantly enhanced [29, 30] (Fig. 2G). These findings collectively suggested that MYOD efficiently accelerates the maturation of skeletal myocytes derived from UiPSM cells within a short time (Fig. 2D).
To further assess the sructural and functional mature of the UiPSM- and iMYOD UiPSM-derived skeletal myocytes, we collected samples of UiPSM direct differentiation and exogenous MYOD-mediated differentiation at day 30 to detect myofiber diameter and the resting membrane potential (RMP). Myotube diameter is a conventional measurement of muscle mass [31, 32]. MyoD-mediated skeletal muscle differentiation resulted in an increase in diameter, and the myotube number also showed a significant increase (Fig. S2A-C). These increases suggest enhanced myogenic differentiation.The resting membrane potential (RMP) of a cell is crucial for transitioning from a resting state to an excitable state, allowing the cell to perform its proper function. Hence, we evaluated the RMP of differentiated cells at day 30. The RMP recorded from UiPSM directly differentiated cells at day 30 was relatively low, with a mean value of − 15.77 ± 4.16 mV. In contrast, UiPSM cells overexpressing MYOD (myocytes) displayed a higher RMP, with a mean value of − 60.27 ± 2.92 mV (Fig. S2D, E).Typically, the resting membrane potential of human myoblast cell lines (AB1167) is − 74.1 ± 0.8 mV [33]. Similar RMP mean values are found in human ES-derived (− 72.25 ± 4.57 mV) and iPS-derived (− 72.12 ± 2.55 mV) myocytes [34]. We detected no significant difference between MYOD-differentiated cells and the reported myocytes using a t-test (Fig. S2E). These results collectively support that MYOD promotes UiPSM cell differentiation to a mature myotube state by day 30.
Recent advancements have demonstrated MYOD's efficiency in converting hESCs and other cell types into skeletal myocytes in vitro [5, 6, 23, 35]. However, when comparing MYOD-reprogrammed hESCs (H9) with UiPSM cells, the former showed lower myogenic efficiency on day 15 (Additional file 1: Fig. S3A, B). Notably, human urine cells (UCs) failed to activate myogenesis (Additional file 1: Fig. S3D). These results suggested an inherent predisposition of UiPSM cells towards skeletal muscle differentiation, surpassing that of hESCs. PAX7, recognized for its role in generating myogenic progenitors from hPSCs [4, 36], was less effective in our studies. Overexpression of PAX7 did not significantly induce MYOD and MRF4 expression in either UiPSM or hESC (H9) cells on day 15 under identical conditions (Additional file 1: Fig. S3C, E), indicating that PAX7 alone might not be sufficient to trigger myogenesis in these cell types. In conclusion, these data suggested that MYOD is uniquely capable of efficiently directing UiPSM cells into more mature skeletal myocytes within a relatively brief period.
UiPSM induced skeletal myocytes contribute to muscle regenerationThe potential of MyoD-reprogrammed hPSCs and hPSC-derived myoblasts for engraftment upon transplantation into mouse muscle has been established in previous studies [8, 37,38,39,40]. To further explore the myogenic potential of UiPSM cells in vivo, an appropriate animal model is necessary. In our study, we transplanted UiPSM and iMYOD UiPSM derived skeletal myocytes into the tibial anterior (TA) muscle of the MITRG mouse, which is an immune-deficient strain like NSG commonly used as a recipient of human hematopoietic cells [41]. Before transplantation, both UiPSM-derived skeletal myocytes and hUCs were transduced with a lentiviral vector carrying luciferase and green fluorescent protein (eGFP) [42]. This approach allows for noninvasive monitoring of cell survival in vivo through bioluminescence imaging (BLI). To induce muscle necrosis, we injected 8 µg of cardiotoxin (CTX) bilaterally into the TA muscle of thirteen MITRG mice. Twenty-four hours after CTX injection, we transplanted UIPSM-derived, MYOD-mediated differentiation to day 30 or no MYOD-mediated differentiation to day 60 skeletal muscle cells into damaged TA muscle. This was done to assess the survival and regenerative capability of these human cells in the mouse model (Fig. 3A). For blank control group, one mouse received CTX in the right hindlimb TA muscle and saline solution in the left hindlimb. The remaining twelve mice were split into two groups. One group received injections of luc+ (luciferase-expressing) UiPSM directly differentiated skeletal myocytes into the right hindlimb TA muscles. Another group received luc+ UiPSM-derived skeletal myocytes with MYOD expression in the right hindlimb TA muscles. As a control, the contralateral TA muscles, which were also pre-injured with CTX, were injected with human urine cells (UCs).
Fig. 3
Transplantation of UiPSM and iMYOD UiPSM cells derived human myocytes in muscle injury model. A Schematic overview of the transplantation methodology for UiPSM and iMYOD UiPSM cell-derived human skeletal myocytes into the TA muscle of MITRG mice, following treatment with cardiotoxin (CTX) for 24 h. Urine cells serve as a negative control and are transplanted into the left tibialis anterior muscle. B Bioluminescence imaging (BLI) signal captured at the right tibialis anterior graft site in a representative MITRG mouse treated with CTX, 1 month after transplantation. UCs transplanted into the left TA muscle in (a), as a negative control. UiPSM derived myocytes transplanted into the rright TA muscle in (b). C Morphological characteristics of TA muscle tissue in after transplanted UiPSM-derive myocytes and UCs. H&E staining of longitudinal sections of TA muscles showed the aggregation of inflammatory factors (asterisk) could still be seen locally in the left tibial anterior muscle after transplanting UCs in (a). H&E staining of longitudinal sections of TA muscles after transplanted UiPSM differentiated into myocytes at day 60 in (b). Scale bars, 100 µm. D Morphological characteristics of TA muscle tissue in after transplanted iMYOD UiPSM-derive myocytes and UCs. H&E staining of longitudinal sections of TA muscles showed the local aggregation of inflammatory factors (asterisk)in the left tibial anterior muscle after transplanting UCs in (a). H&E staining of longitudinal sections of TA muscles after transplanted iMYOD UiPSM-derived myocytes at day 30 in (b). Scale bars, 100 µm. E TA muscle from UiPSM-derived myocytes evaluated for the expression of myocyte-specific markers. Longitudinal section showed the colocalization of Desmin (DES) and Myosin Heavy Chain (MHC) (arrows in (a.)). Transversal section showed the colocalization of Desmin and Laminin (arrows in (b.)). Transversal section showed the colocalization of human nuclei antibody (hNA) and MYOD (arrows in (c.)). Scale bars, 100 µm. F TA muscle from iMYOD UiPSM-derived myocytes evaluated for the expression of myocyte-specific markers. Colocalization of Desmin (DES) and Myosin Heavy Chain (MHC) (arrows in (a.), and colocalization of Desmin and Laminin (arrows in (b.)) were shown in Longitudinal sections. Transversal section showed the colocalization of human nuclei antibody (hNA) and MYOD (arrows in (c.)). Scale bars, 100 µm
One month after cardiotoxin injury and subsequent grafting, the mice exhibited a stable bioluminescence imaging (BLI) signal in the right hindlimb. This signal was indicative of graft survival of both UiPSM and iMYOD UiPSM cell-derived skeletal myocytes. In contrast, the left tibialis anterior muscle of the same mice, injected with luciferase-expressing urine cells (luc + UC), showed no BLI signal, thereby underscoring the robust long-term survival of the transplanted skeletal myocytes in the host muscle (Fig. 3B). The aggregation of inflammatory factors could still be seen locally in the left tibial anterior muscle by H&E staining (Fig. 3C, D), also suggested UiPSM and iMOD UiPSM cells derived skeletal myocytes repaired the damaged muscle. Further validation of engraftment and contribution to muscle regeneration was achieved through immunohistochemical analysis. Muscle sections were stained with antibodies specific to Desmin, Myosin Heavy Chain (MHC), Laminin, MYOD, and Human Nuclear Antigen (hNA) (Fig. 3E, F). The expression of these markers in the muscle tissue provided strong evidence that the UiPSM-derived skeletal myocytes were contributing to its regeneration. An important observation was the absence of tumor formation in the transplanted mice, even over a longer period of 2 months.
UiPSM derived skeletal myocytes mature after transplantationTo comprehensively analyze the cell diversity at the transplanted sites (Fig. 3A), single-cell RNA sequencing (scRNA-seq) was performed on UiPSM and iMYOD UiPSM cells-derived skeletal myocytes, as well as the control urine cells, engrafted into the TA muscles after 1 month. Considering the presence of both human and mouse cells in the grafts, the scRNA-seq data were aligned to both human and mouse genomes accordingly (Fig. 4A). Of the 4159 individual cells analyzed, the experimental group showed more than 80% genome matching with the human genome and less than 20% with the mouse genome. In contrast, the control group exhibited nearly 80% genome matching with the mouse genome and only 15% with the human genome (Fig. 4B). This data indicated that both UiPSM and iMYOD UiPSM-derived skeletal myocytes not only survived but also actively participated in muscle regeneration post-transplantation into the damaged TA muscle.
Fig. 4
scRNA-seq analysis of the cell components of the human skeletal myocytes derived grafts. A Schematic overview of the process involving transplantation of UiPSM and iMYOD UiPSM-derived human myocytes and urine cells into injured mouse TA muscles. One month after transplantation, TA muscles were dissociated into single cells for RNA sequencing. Sequencing data were then mapped to the human (hg38) and mouse (mm10) genomes, respectively. B Quantification of cell numbers and mapping rates from the sequencing data. C UMAP representation of scRNA data from Xi et al.(2020) and our studies, including eight sources of myogenic. D t-SNE plot showed the projection of 2137 cells mapping to human genome (hg38). These cells could be divided in 11 color-coded clusters. The circles, triangles and diamonds represent the control group (urine cells), and UiPSM and iMYOD UiPSM derived human myocytes, respectively. E Gene ontology (GO) analysis revealed gene expression profiles of the groups (H1, h5, h7, h8, h9, h10) that are highly associated with skeletal muscle development in the above 11 clusters. p value < 0.05. F Violin plot showing the expression and ratio of typical mature skeletal muscle cells related genes in each cluster. G Dot plot of expression levels for titin (TTN) and different Myh genes. Color indicates expression level in each cluster; Dot size percent of cells expressing selected genes in each cluster. H Violin plot of expression levels for calcium-handling gene PVALB, Calcium-Transporting ATPase Sarcoplasmic Reticulum (ATP1A2, ATP2A1), Calmodulin (CALM1, CALM2) and troponin (TTN, TNNI2, TNNT3, TNNC2). I Calculating the percentage of cells in skeletal myocytes related clusters (H1, h5, h7, h8, h9, h10) on all sequenced cells of the grafts from UiPSM and iMYOD UiPSM derived human myocytes, when mapping with human genome (hg38)
To gain a comprehensive view of UiPSM-derived myocytes close to human skeletal muscle cell ontogeny, we evaluated the scRNA-seq data of UiPSM derived myocytes after transplantation and reported human myocytes, including SKM cluster in scRNA-seq data of human limb muscle tissues from embryonic (week 5–8), fetal (week 9–18) as well as postnatal juvenile (year 7–11) and adult (year 34–42) stages [43], most of the cells in our lab overlapped with embryonic (week 7–8), fetal (week 9) myocytes (Fig. 4C). Typically, the primary generation fibers with central nuclei are present in the human skeletal muscle at week 8 of gestation [44]. Human fetal skeletal muscle (week 9) enriched for muscle contraction, mitochondria and oxidative phosphorylation (OxPhos) as well as calcium signaling [43]. These suggested UiPSM derived mature skm state after transplantation. At the same time, we also compared the data of UiPSM derived myocytes after transplantation with SkM cells generated from hPSCs in representative protocols [22, 35, 43, 45], we only found a large group of cells coincide with MS protocol during week 6–7 differentiation, and a small group of cells match to SKM cells in HX protocol at week 4 (Fig. 4C). A small subset of terminal differentiating myoblasts-myocytes (MBMC) were observed in MS protocol at week 6–7, and half of SKM cells were PAX7-GFP+ populations in HX protocol at week 4 [43]. These suggested UiPSM derived terminal differentiating after transplantation, and retained a group of PAX7 positive populations. Furthermore, a small group of cells neared to adult (year 34–42) myogenic cells (Fig. 4C), confirmed a mature SKM state in UiPSM derived myocytes after transplantation.
Upon detailed examination, the scRNA-seq data from human skeletal myocyte grafts could be categorized into 11 distinct clusters when mapped to the human genome (hg38). Moreover, the data UiPSM and iMYOD UiPSM cell derived skeletal myocytes showed similar distribution (Fig. 4C, D), suggested skeletal myocytes in different levels of maturity showed an almost uniform ability to differentiate in the body in vivo. Cluster H1 enriched for muscle contraction and muscle system process related genes, TNNI1, MYL4, DDX17, TPM3, KRT8 and KRT18. Cluster H5 was specifically associated with myoblast differentiation and proliferation, highly enriched for DDX17, TPM3, KRT8 and KRT18. Actin filament bundle assembly related genes, TNNC1, TPM3, BMP2,
留言 (0)