
記住我

The space race set in motion by the Cold War intensified in the 1960s, with the Soviet Union first paving the path to manned space flight in 1961 (1). In high relevance to astronauts' mission readiness (2–5), experiments conducted in this decade, contrasting persons with functioning and bilaterally non-functioning labyrinths, solidified the contributions of the vestibular inputs to production of spatial perception and motion sickness during exposure to passive motion or microgravity (6). However, all forms of labyrinthine deactivation are not created equal. For example, patients may undergo total vestibular nerve section as a treatment for vertigo due to eighth nerve neuroma or Ménière's disease (7, 8); others may for varying causes sustain selective damage within the vestibular end-organs while retaining functioning nerves, as may be the case for candidates of vestibular implants (9, 10). Selective inactivation of otolithic signals may even be induced environmentally under microgravity in space. Examining information processing in the central vestibular mechanism known as velocity storage in various contexts of labyrinthine inactivation may shed further light on health and perceptual anomalies during or following space flight.
What is velocity storage?Activated by head rotation, large-field visual motion, or proprioceptive cues for continuous rotation, velocity storage is a central neural circuit that sustains a rotational velocity estimate of ongoing self-motion (11–16). The mechanism serves a working memory-like function of self-motion and spatial orientation to shape ocular and postural reflexes in the brainstem and presumably perception in the cerebral cortex (13–15, 17–19).
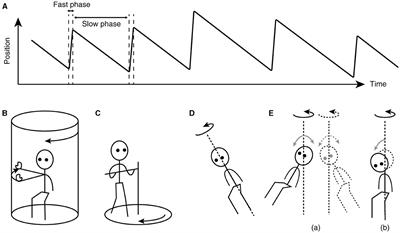
The key to understanding the working of velocity storage has been in eye movement measurement. Nystagmus, such as occurs during the vestibulo-ocular reflex (VOR), is an automatic eye movement made in response to signals that indicate self-rotation relative to the surrounding world. Nystagmus facilitates the acquisition of visual information with the eyes moving in opposition to and in compensation for the detected rotation (slow phases) but for quick “resetting” interruptions (fast phases; Figure 1A). The presence of a common, sub-cerebral cortical mechanism for multimodal sensory integration had long been presumed based on experimental observations such as: (a) coordinated shaping, rather than a simple superposition, of nystagmus of different sensory origins takes place during combined stimulation (11); (b) the nystagmic reaction to either visual (optokinetic) or vestibular stimulation is subject to a “central inertia,” demonstrated with a prolonged response or after-response (11); (c) optokinetic nystagmus (OKN) can be induced in animals with visual cortical lesions (11, 20); and (d) bilateral vestibular nerve sections degrade OKN and abolish optokinetic after-nystagmus (OKAN) (21, 22).
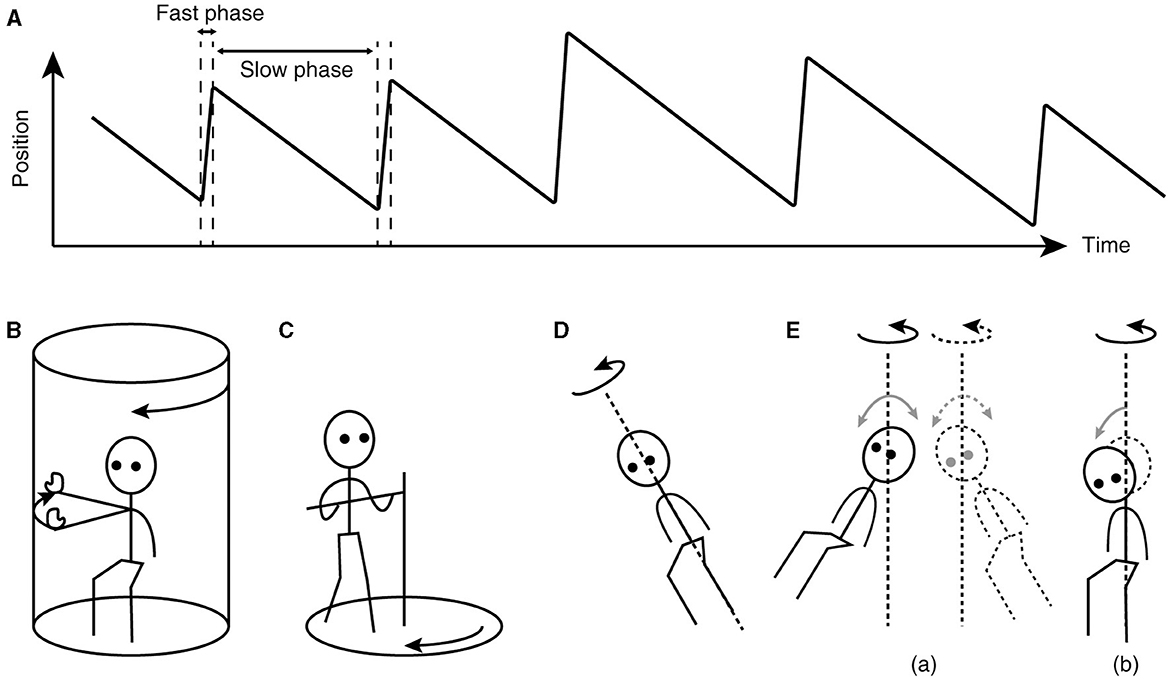
Figure 1. Characteristics of nystagmus and examples of unusual stimuli that can induce nystagmus. (A) Sawtooth pattern of eye position change over time during nystagmus. Slow-phase movements are directed against, thus in compensation for, the detected head rotation, whereas fast-phase movements facilitate maintenance of eye position within the oculomotor range. (B) Arthrokinetic stimulation. (C) Circular treadmill locomotion. (D) OVAR. (E) Whole-body sinusoidal PWR (a). As a variation in human testing, a disorienting sensation and motion sickness may be induced with a discrete, rather than continuous, head tilt while seated in a rotating chair (b). Note that in (B, C), the head stays spatially stationary, and when conducted in darkness, the stimuli can generate nystagmus without labyrinthine or visual input. In (D, E), at steady state, the semicircular canals do not signal the rotation continuing about the axis indicated by the dotted line.
Animal experiments in the 1970s demonstrated that spiking activity in certain second-order vestibular neurons correlated with the strength of either vestibularly- or optokinetically-induced nystagmus (23–25). With this discovery, the functional operation of the central processing system that soon came to be known as velocity storage became a valuable focus of rigorous model-based testing of behavioral and physiological data (13–15, 26, 27), These neurons belong to the vestibular-only (VO) class, so-termed to differentiate from other secondary vestibular neurons that display saccade or eye position sensitivity (28). Importantly, while the decay time constant of the primary afferent activity after the cessation of head angular acceleration, contributed by the semicircular canal endolymph's inertia, amounts only to 3–6 s, the activity of VO neurons is typically characterized with a time constant of 10–40 s, corresponding to the aforementioned “central inertia” (25, 29–32).
Besides vestibular or optokinetic means, and indeed in the absence of actual motion or visual motion, an illusion of body rotation (circular vection) and corresponding compensatory nystagmus can be induced in a stationary person when a rotating surround is passively followed with an extended arm (Figure 1B) (12) or when the person steps along a fictitious, circular trajectory on a counter-rotating floor (Figure 1C) (16). Compensatory nystagmus is also generated in the absence of vision or steady activation of coplanar semicircular canals when the subject is continuously rotated about a tilted axis (off-vertical axis rotation—OVAR; Figure 1D) (33–35) or when sinusoidally pitched or rolled during earth-vertical axis rotation (pitch/roll while rotating—PWR/RWR; Figure 1E) (36, 37).
That nystagmus can be generated and sustained without coplanar optokinetic or semicircular canal activation indicates that the velocity storage mechanism actively reconstitutes self-motion signals from multimodal sensory inputs. Additionally, the stored estimate of self-motion can be dynamically transformed such that the outcome nystagmus tends to align to the gravito-inertial field. For example, horizontal OKN induced while tilted laterally gives way to OKAN with a vertical component (38–40). Similarly, the per-rotatory nystagmus in response to off-center rotation with forward or backward tangential motion develops an out-of-plane, vertical component as the centripetal acceleration tilts the gravito-inertial field (41). Thus, velocity storage is also equipped with orienting properties to act as a “neural gyroscope” (40, 42, 43), making it more fitting to be identified as “working memory-like” rather than mere storage.
The properties of velocity storage are nearly exclusively characterized by the dynamics of slow-phase eye velocity of nystagmus. However, many of so-characterized properties have also been demonstrated in the activity of VO neurons (29, 44, 45). Insight into the working of velocity storage has been conversely derived from VO neuron activity as well. For example, while most VO neurons receive convergent inputs from different semicircular canal and otolith afferents, many show preferences to activation near or orthogonal to the plane of a specific push-pull canal pair (29, 46). Thus, VO neurons appear to collectively code the three-dimensional action of velocity storage using a coordinate system consistent with the geometric alignment of the semicircular canals and extraocular muscles, which is common across lateral- or frontal-eyed species (29, 47). Further, while vestibular afferents cannot differentiate active from passive motion, VO neurons reportedly show different responses (48–51). Accordingly, velocity storage may be activated and controlled differently during active and passive motion.
Velocity storage is under the inhibitory control of the cerebellar nodulus and, likely, the adjacent caudal uvula. Damage to these areas destabilizes velocity storage, triggering periodic alternating nystagmus and elongating the time constant of the decay of the VOR nystagmus during a rotational test (52–55). Such damage also results in loss of compensatory nystagmus during OVAR and sinusoidal PWR/RWR (56, 57) as well as in a compromised ability to reorient eye velocity to the gravito-inertial field (55, 57, 58). Reversible unilateral inactivation of the nodulus induces spontaneous nystagmus in darkness with contralaterally-directed slow phases and abnormal eye velocity orientation to gravity (59). The nodulus and caudal uvula target a wide variety of cell types in the vestibular nuclei, including VO neurons (60). Nodular micro-electrical stimulation results in shortening of the decay time constant of the VOR or OKAN with ipsilateral slow phases (60, 61), but may also yield further different effects depending on the duration and exact location of the stimulation (59, 62).
Lastly, velocity storage is malleable. For example, repeated rotation in darkness shortens the VOR decay time constant (decreased “central inertia”) in a long-retained effect known as vestibular habituation (63–65). The spatial orientation properties of velocity storage, such as observed in eye movements outside the stimulus plane during the VOR, can also be modified (66, 67), possibly to facilitate adaptation to a new gravito-inertial environment (68–70). Relatedly, the spatial tuning of VO neurons can be modified with prolonged tilt (71). As the cerebellum plays a major role in motor learning and calibration (72, 73), the malleability of velocity storage likely depends on the nodulus and uvula (64). Mal de débarquement syndrome, a chronic illness primarily characterized by a persistent illusory perception of self-motion, is thought to result from a failure in velocity storage to readapt to a normal acceleration environment after adapting to passive motion (74–76).
Correlates of velocity storage perturbationStudies conducted in humans and animals with bilaterally non-functioning labyrinths, including those from the 1960s, found that these subjects were immune to motion sickness when exposed to otherwise highly provocative vestibular tests, such as PWR/RWR, OVAR, and other unusual motion stimuli (6, 77–79). Such immunity merits re-examination in parallel with patterns of velocity storage activation (Table 1). Differences and similarities in perceptual experiences are also considered when illuminating information is available.

Table 1. Exemplar effects of localized vestibular-related functional inactivation.
Vestibular habituation, identified with a shortened velocity storage time constant, reportedly reduces susceptibility to motion sickness (101, 102). The GABA-B agonist baclofen has been indicated to also reduce the velocity storage time constant and motion sickness susceptibility to RWR, but reversibly (103, 104). Given that bilateral vestibular nerve sections result in loss of OKAN (21, 22), and that loss of or impaired vestibular functions due to ototoxicity or other causes significantly reduces OKAN (80–83), the immunity to motion sickness to provocative motion stimuli in labyrinthine-defective individuals may be explained by their loss of or reduced ability to store velocity signals. However, motion sickness and circular vection can still be induced with head movements in a rotating visual environment after vestibular neurectomy (7). The effect of baclofen on visually induced motion sickness is not known.
Neuronally, while the ability to store velocity signals appears to depend on the wellbeing of the primary afferents, there is no evidence that VO neurons undergo anterograde transneuronal degeneration after vestibular nerve sections (105, 106). Neuronal activities ascribed to velocity storage functions that survive labyrinthine deactivation remain to be identified. Behaviorally however, nystagmus and circular vection can be induced in labyrinthine-defective individuals during fictitious circular stepping around in darkness (Figure 1C) (16), providing strong evidence that reconstitution of self-motion velocity signals and their storage are separate processes.
Motion sickness is typically induced in a context of passive rather than active motion (107, 108). Likewise, VO neurons are reportedly more sensitive to passive than active head movements (48–51). Participation of these neurons in the vestibular-autonomic circuits is not known (109–113), but motion sickness susceptibility is likely not directly increased as a simple consequence of increased VO neuron activation. For example, while congruent optokinetic and vestibular inputs can synergistically activate VO neurons and incongruent ones antagonistically (25–27, 29, 114), vision of a stationary surround (i.e., optokinetic input congruent with physical motion) protects against motion sickness and perceptual disorientation during PWR/RWR or OVAR (79, 115–120). Curiously, under microgravity in space or during parabolic flight, active head movements with vision of a stationary surround is more provocative than the same movements without vision (3, 96).
Also curiously, while PWR/RWR is highly provocative of motion sickness and perceptually disorienting on earth, it is not so under microgravity in space or during parabolic flight (3, 94, 95). Mirroring this environmental inactivation of otolithic signals, after bilateral utricular nerve section and saccular macula destruction, previously susceptible squirrel monkeys reportedly become immune to motion sickness while unrestrained inside a rotating cage (89), a condition that likely generates provocativeness through a PWR/RWR-like mechanism. By contrast, semicircular canal deactivation precipitated by ototoxicity, sparing otolithic functions, has also been shown to induce immunity to motion sickness in unrestrained squirrel monkeys in a rotating cage (121). Thus, the provocativeness of PWR/RWR appears to arise from central integration of both the semicircular canal and otolithic signals. By parallel, the ability to generate nystagmus during sinusoidal PWR/RWR is lost in macaque monkeys after selective deactivation of the semicircular canal function with canal plugging (36, 85). The counterpart effect of selective deactivation of the otolithic input does not seem to have been experimentally tested, but nystagmus generation during sinusoidal PWR/RWR is also thought to require both otolithic and semicircular canal inputs (57, 67, 85).
The involvement of the cerebellar nodulus and uvula provides another parallel between velocity storage and motion sickness. The nodulo-uvular involvement in motion sickness production has long been suspected based on observations that nodulo-uvular lesions rendered experimental animals immune to motion sickness (97, 98). Doubts raised against this view (99, 100) may be partially addressed by considering the acute vs. chronic effects of such lesions and the presence of parallel vestibular-cerebellar circuits that can produce counteracting autonomic effects (52, 53, 111, 113, 122, 123). It is presumably within such complex neural interactions that learning takes place so that evoked symptoms of motion sickness and future susceptibility diminish even when the provocative situation is unchanged (4, 70). Motion sickness is said to be most severe when the orientation and autonomic regulation systems are undergoing rapid recalibration (4). Whether understanding the neural basis of velocity storage malleability improves the predictability of motion sickness susceptibility to specific stimuli or situations remains to be seen. The physiology of and the circuitries that control and are controlled by VO neurons likely serve a focal point of future studies (19, 113), an approach thus far possible chiefly in animal-based experiments.
ConclusionClues from various forms of labyrinthine and central loss of vestibular function connect spatial perception, motion sickness, and velocity storage as parallel phenomena. Similar partially overlapping parallelisms from different perspectives have previously been suggested (19, 113). A possible pitfall of such thinking is that these phenomena may be just that—parallel but unrelated. No matter, the paths to fill the knowledge gaps are largely unpaved and promise abundant scientific opportunities. Continuing development in space exploration and technology behooves us to advance the field.
Author contributionsJM: Writing – original draft, Writing – review & editing.
FundingThe author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interestThe author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's noteAll claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
AbbreviationsOKAN, optokinetic after-nystagmus; OKN, optokinetic nystagmus; OVAR, off-vertical axis rotation; PWR, pitch while rotating; RWR, roll while rotating; VO, vestibular-only; VOR, vestibulo-ocular reflex.
References3. Graybiel A, Miller EF, Homick JL. Individual differences in susceptibility to motion sickness among six Skylab astronauts. Acta Astronaut. (1975) 2:155–74. doi: 10.1016/0094-5765(75)90051-X
PubMed Abstract | Crossref Full Text | Google Scholar
6. Graybiel A. Functional disturbances of vestibular origin of significance in space flight. In:Bjurstedt H, , editor. Proceedings of the Second International Symposium on Basic Environmental Problems of Man in Space. Vienna: Springer (1967). p. 8–32.
PubMed Abstract | Google Scholar
9. Sluydts M, Curthoys I, Vanspauwen R, Papsin BC, Cushing SL, Ramos A, et al. Electrical vestibular stimulation in humans: a narrative review. Audiol Neurootol. (2020) 25:6–24. doi: 10.1159/000502407
PubMed Abstract | Crossref Full Text | Google Scholar
10. Strupp M, Brandt T, Dieterich M. “Bilateral Vestibulopathy,” Vertigo and Dizziness. Cham: Springer International Publishing (2023). p. 99–118.
11. ter Braak J. Untersuchungen über optokinetischen Nystagmus. Arch Neerl Physiol Homme Anim. (1936) 21:309–76.
PubMed Abstract | Google Scholar
13. Cohen B, Matsuo V, Raphan T. Quantitative analysis of the velocity characteristics of optokinetic nystagmus and optokinetic after-nystagmus. J Physiol. (1977) 270:321–44. doi: 10.1113/jphysiol.1977.sp011955
PubMed Abstract | Crossref Full Text | Google Scholar
14. Raphan T, Cohen B, Matsuo V. A velocity-storage mechanism responsible for optokinetic nystagmus (OKN), optokinetic after-nystagmus (OKAN) and vestibular nystagmus. In:Baker R, Berthoz A, , editors. Control of Gaze by Brain Stem Neurons. Developments in Neuroscience. Amsterdam: Elsevier (1977). p. 37–47.
15. Robinson DA. Vestibular and optokinetic symbiosis: an example of explaining by modelling. In:Baker R, Berthoz A, , editors. Control of Gaze by Brain Stem Neurons. Developments in Neuroscience. Amsterdam: Elsevier (1977). p. 49–58.
17. Bertolini G, Ramat S, Laurens J, Bockisch CJ, Marti S, Straumann D, et al. Velocity storage contribution to vestibular self-motion perception in healthy human subjects. J Neurophysiol. (2011) 105:209–23. doi: 10.1152/jn.00154.2010
PubMed Abstract | Crossref Full Text | Google Scholar
21. Cohen B, Uemura T, Takemori S. Effects of labyrinthectomy on optokinetic nystagmus (OKN) and optokinetic after-nystagmus (OKAN). Int J Equilib Res. (1973) 3:88–93.
PubMed Abstract | Google Scholar
23. Waespe W, Henn V. Behaviour of secondary vestibular units during optokinetic nystagmus and after-nystagmus in alert monkeys. Pflugers Arch. (1976) 362:R50.
25. Waespe W, Henn V. Neuronal activity in the vestibular nuclei of the alert monkey during vestibular and optokinetic stimulation. Exp Brain Res. (1977) 27:523–38. doi: 10.1007/BF00239041
PubMed Abstract | Crossref Full Text | Google Scholar
27. Waespe W, Henn V. The velocity response of vestibular nucleus neurons during vestibular, visual, and combined angular acceleration. Exp Brain Res. (1979) 37:337–47. doi: 10.1007/BF00237718
PubMed Abstract | Crossref Full Text | Google Scholar
28. Fuchs AF, Kimm J. Unit activity in vestibular nucleus of the alert monkey during horizontal angular acceleration and eye movement. J Neurophysiol. (1975) 38:1140–61. doi: 10.1152/jn.1975.38.5.1140
PubMed Abstract | Crossref Full Text | Google Scholar
30. Fernandez C, Goldberg JM. Physiology of peripheral neurons innervating semicircular canals of the squirrel monkey. II. Response to sinusoidal stimulation and dynamics of peripheral vestibular system J Neurophysiol. (1971) 34:661–75. doi: 10.1152/jn.1971.34.4.66.1
Crossref Full Text | Google Scholar
31. Goldberg JM, Fernandez C. Physiology of peripheral neurons innervating semicircular canals of the squirrel monkey. I. Resting discharge and response to constant angular accelerations. J Neurophysiol. (1971) 34:635–60. doi: 10.1152/jn.1971.34.4.635
PubMed Abstract | Crossref Full Text | Google Scholar
32. Dai M, Klein A, Cohen B, Raphan T. Model-based study of the human cupular time constant. J Vestib Res. (1999) 9:293–301. doi: 10.3233/VES-1999-9407
Crossref Full Text | Google Scholar
33. Guedry FE. Orientation of the rotation-axis relative to gravity: its influence on nystagmus and the sensation of rotation. Acta Otolaryngol. (1965) 60:30–48. doi: 10.3109/00016486509126986
PubMed Abstract | Crossref Full Text | Google Scholar
34. Benson AJ, Bodin MA. Interaction of linear and angular accelerations on vestibular receptors in man. Aerosp Med. (1966) 37:144–54.
PubMed Abstract | Google Scholar
35. Cohen B, Suzuki JI, Raphan T. Role of the otolith organs in generation of horizontal nystagmus: effects of selective labyrinthine lesions. Brain Res. (1983) 276:159–64. doi: 10.1016/0006-8993(83)90558-9
PubMed Abstract | Crossref Full Text | Google Scholar
38. Clément G, Lathan CE. Effects of static tilt about the roll axis on horizontal and vertical optokinetic nystagmus and optokinetic after-nystagmus in humans. Exp Brain Res. (1991) 84:335–41. doi: 10.1007/BF00231454
PubMed Abstract | Crossref Full Text | Google Scholar
39. Dai MJ, Raphan T, Cohen B. Spatial orientation of the vestibular system: dependence of optokinetic after-nystagmus on gravity. J Neurophysiol. (1991) 66:1422–39. doi: 10.1152/jn.1991.66.4.1422
PubMed Abstract | Crossref Full Text | Google Scholar
40. Raphan T, Cohen B. Organizational principles of velocity storage in three dimensions: the effect of gravity on cross-coupling of optokinetic after-nystagmus. Ann N Y Acad Sci. (1988) 545:74–92. doi: 10.1111/j.1749-6632.1988.tb19556.x
PubMed Abstract | Crossref Full Text | Google Scholar
41. Merfeld DM, Young LR, Tomko DL, Paige GD. Spatial orientation of VOR to combined vestibular stimuli in squirrel monkeys. Acta Otolaryngol Suppl. (1991) 481:287–92. doi: 10.3109/00016489109131403
PubMed Abstract | Crossref Full Text | Google Scholar
42. Angelaki DE, Hess BJ. Inertial representation of angular motion in the vestibular system of rhesus monkeys. I. Vestibuloocular reflex. J Neurophysiol. (1994) 71:1222–49. doi: 10.1152/jn.1994.71.3.1222
PubMed Abstract | Crossref Full Text | Google Scholar
44. Reisine H, Raphan T. Neural basis for eye velocity generation in the vestibular nuclei of alert monkeys during off-vertical axis rotation. Exp Brain Res. (1992) 92:209–26. doi: 10.1007/BF00227966
PubMed Abstract | Crossref Full Text | Google Scholar
47. Simpson JI, Graf W. Eye-muscle geometry and compensatory eye movements in lateral-eyed and frontal-eyed animals. Ann N Y Acad Sci. (1981) 374:20–30. doi: 10.1111/j.1749-6632.1981.tb30856.x
PubMed Abstract | Crossref Full Text | Google Scholar
48. Gdowski GT, McCrea RA. Integration of vestibular and head movement signals in the vestibular nuclei during whole-body rotation. J Neurophysiol. (1999) 82:436–49. doi: 10.1152/jn.1999.82.1.436
PubMed Abstract | Crossref Full Text | Google Scholar
49. McCrea RA, Gdowski GT, Boyle R, Belton T. Firing behavior of vestibular neurons during active and passive head movements: vestibulo-spinal and other non-eye-movement related neurons. J Neurophysiol. (1999) 82:416–28. doi: 10.1152/jn.1999.82.1.416
PubMed Abstract | Crossref Full Text | Google Scholar
52. Jeong H-S, Oh JY, Kim JS, Kim J, Lee AY, Oh S-Y. Periodic alternating nystagmus in isolated nodular infarction. Neurology. (2007) 68:956–7. doi: 10.1212/01.wnl.0000257111.24769.d2
PubMed Abstract | Crossref Full Text | Google Scholar
55. Wearne S, Raphan T, Cohen B. Control of spatial orientation of the angular vestibuloocular reflex by the nodulus and uvula. J Neurophysiol. (1998) 79:2690–715. doi: 10.1152/jn.1998.79.5.2690
PubMed Abstract | Crossref Full Text | Google Scholar
56. Angelaki DE, Hess BJ. Lesion of the nodulus and ventral uvula abolish steady-state off-vertical axis otolith response. J Neurophysiol. (1995) 73:1716–20. doi: 10.1152/jn.1995.73.4.1716
PubMed Abstract | Crossref Full Text | Google Scholar
57. Angelaki DE, Hess BJ. Inertial representation of angular motion in the vestibular system of rhesus monkeys. II. Otolith-controlled transformation that depends on an intact cerebellar nodulus. J Neurophysiol. (1995) 73:1729–51. doi: 10.1152/jn.1995.73.5.1729
PubMed Abstract | Crossref Full Text | Google Scholar
58. Cohen B, John P, Yakushin SB, Buettner-Ennever J, Raphan T. The nodulus and uvula: source of cerebellar control of spatial orientation of the angular vestibulo-ocular reflex. Ann N Y Acad Sci. (2002) 978:28–45. doi: 10.1111/j.1749-6632.2002.tb07553.x
PubMed Abstract | Crossref Full Text | Google Scholar
59. Sheliga BM, Yakushin SB, Silvers A, Raphan T, Cohen B. Control of spatial orientation of the angular vestibulo-ocular reflex by the nodulus and uvula of the vestibulocerebellum. Ann N Y Acad Sci. (1999) 871:94–122. doi: 10.1111/j.1749-6632.1999.tb09178.x
PubMed Abstract | Crossref Full Text | Google Scholar
60. Meng H, Blázquez PM, Dickman JD, Angelaki DE. Diversity of vestibular nuclei neurons targeted by cerebellar nodulus inhibition. J Physiol. (2014) 592:171–88. doi: 10.1113/jphysiol.2013.259614
PubMed Abstract | Crossref Full Text | Google Scholar
62. Heinen SJ, Oh DK, Keller EL. Characteristics of nystagmus evoked by electrical stimulation of the uvular/nodular lobules of the cerebellum in monkey. J Vestib Res. (1992) 2:235–45. doi: 10.3233/VES-1992-2306
PubMed Abstract | Crossref Full Text | Google Scholar
63. Clément G, Tilikete C, Courjon J-H. Retention of habituation of vestibulo-ocular reflex and sensation of rotation in humans. Exp Brain Res. (2008) 190:307–15. doi: 10.1007/s00221-008-1471-0
PubMed Abstract | Crossref Full Text | Google Scholar
64. Cohen H, Cohen B, Raphan T, Waespe W. Habituation and adaptation of the vestibuloocular reflex: a model of differential control by the vestibulocerebellum. Exp Brain Res. (1992) 90:526–38. doi: 10.1007/BF00230935
PubMed Abstract | Crossref Full Text | Google Scholar
65. Jäger J, Henn V. Habituation of the vestibulo-ocular reflex (VOR) in the monkey during sinusoidal rotation in the dark. Exp Brain Res. (1981) 41:108–14. doi: 10.1007/BF00236599
PubMed Abstract | Crossref Full Text | Google Scholar
66. Dai M, Raphan T, Cohen B. Adaptation of the angular vestibulo-ocular reflex to head movements in rotating frames of reference. Exp Brain Res. (2009) 195:553–67. doi: 10.1007/s00221-009-1825-2
PubMed Abstract | Crossref Full Text | Google Scholar
67. Maruta J. Lasting alteration of spatial orientation induced by passive motion in rabbits and its possible relevance to mal de débarquement syndrome. Front Neurol. (2023) 14:1110298. doi: 10.3389/fneur.2023.1110298
PubMed Abstract | Crossref Full Text | Google Scholar
68. Graybiel A. Angular velocities, angular accelerations, and Coriolis accelerations. In:Gazenko OG, Calvin M, , editors. Foundations of Space Biology and Medicine: Joint USA/USSR Publication. Washington, DC: National Aeronautics and Space Administration/Academy of Sciences of the USSR (1975). p. 247–304.
71. Eron JN, Cohen B, Raphan T, Yakushin SB. Adaptation of orientation vectors of otolith-related central vestibular neurons to gravity. J Neurophysiol. (2008) 100:1686–90. doi: 10.1152/jn.90289.2008
PubMed Abstract | Crossref Full Text | Google Scholar
72. Ito M. The Cerebellum and Neural Control. New York, NY: Raven Press (1984).
74. Dai M, Cohen B, Smouha E, Cho C. Readaptation of the vestibulo-ocular reflex relieves the mal de debarquement syndrome. Front Neurol. (2014) 5:124. doi: 10.3389/fneur.2014.00124
PubMed Abstract | Crossref Full Text | Google Scholar
75. Maruta J, Cho C, Raphan T, Yakushin SB. Symptom reduction in mal de débarquement syndrome with attenuation of the velocity storage contribution in the central vestibular pathways. Front Rehabil Sci. (2024) 5:1331135. doi: 10.3389/fresc.2024.1331135
PubMed Abstract | Crossref Full Text | Google Scholar
76. Yakushin SB, Raphan T, Cho C. Treatment of gravitational pulling sensation in patients with mal de debarquement syndrome (MdDS): a model-based approach. Front Integr Neurosci. (2022) 16:801817. doi: 10.3389/fnint.2022.801817
PubMed Abstract | Crossref Full Text | Google Scholar
79. Watanuki K, Takahashi M, Ikeda T. Perception of surrounding space controls posture, gaze, and sensation during Coriolis stimulation. Aviat Space Environ Med. (2000) 71:381–7.
留言 (0)