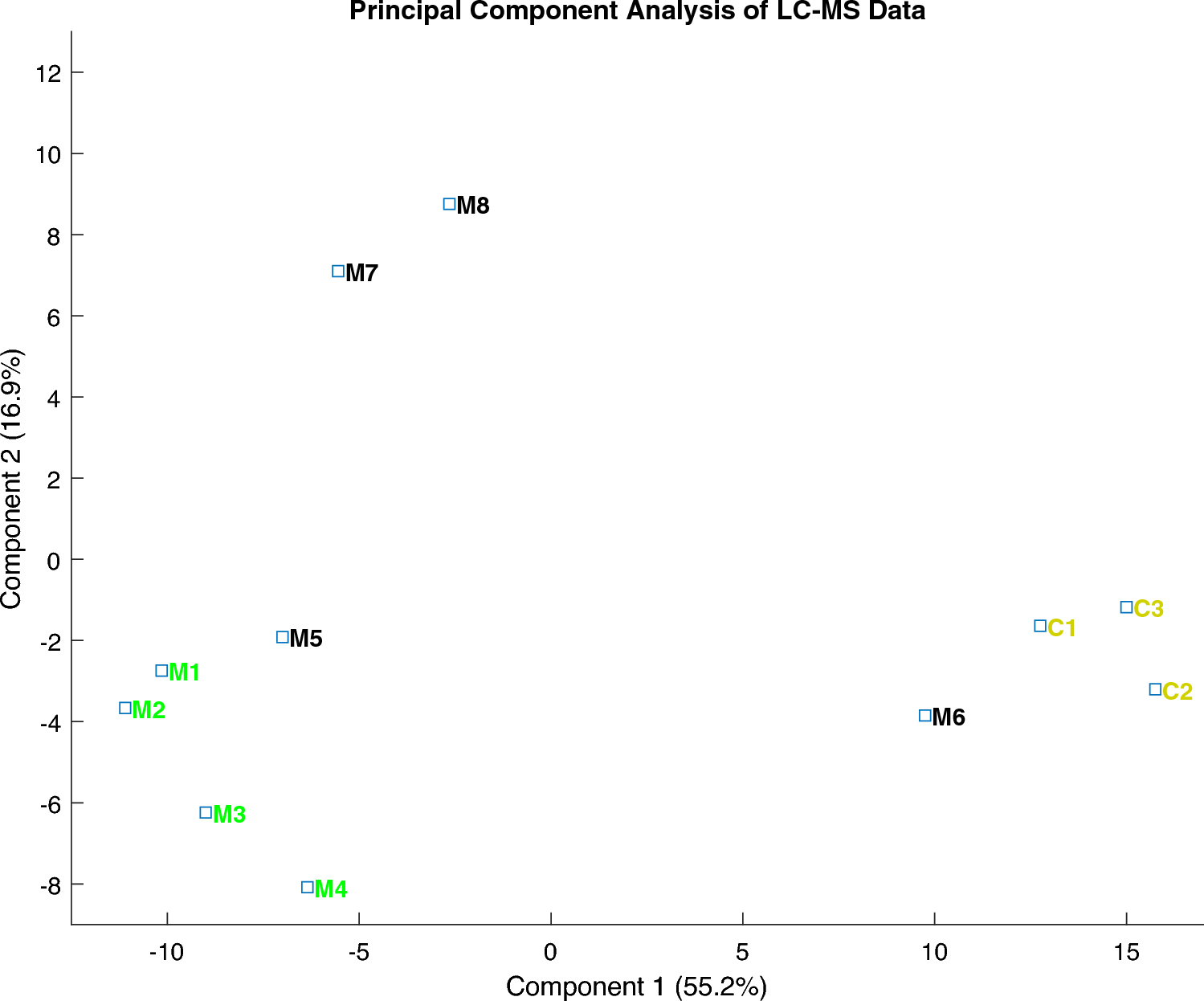
A comparative analysis was conducted between normal enamel and MIH enamel. By employing a microdissection methodology combined with a LC–MS technique, the study reveals the potential to distinguish distinctive proteomic profiles within MIH enamel that display varying clinical severity grades. The proteomic profiles of controls and individuals in the MIH groups I and II exhibit a well-separable appearance.
Throughout the study, a quantitative analysis was conducted to identify proteins with statistically significant differences and their associated biological processes. To underscore these findings, the proteomic results are discussed in relation to previous LC–MS studies and incorporated with data from other research focusing on healthy enamel or MIH enamel from a proteomic perspective. In the field of proteomic analysis of hard tissues, particularly erupted permanent human enamel, the predominant focus has been on two fundamental techniques: LC–MS (Castiblanco et al. 2015; Jagr et al. 2019; Mukthar et al. 2022, Rexhaj et al. 2023) and SDS-PAGE (Açil et al. 2005; Farah et al. 2010). LC–MS, as a cutting-edge technology, has revolutionized the field by enabling quantitative analysis and the detection of a more extensive array of proteins compared to SDS-PAGE. Furthermore, it is essential to acknowledge the limited number of studies in the literature that have explored healthy permanent enamel using LC–MS, each employing slightly different methodologies (Jagr et al. 2019; Castiblanco et al. 2015; Rexhaj et al. 2023). As far as available knowledge extends, only Mukthar et al. (2022) conducted a comparative analysis of healthy permanent enamel with MIH through the application of LC–MS. Thus, this study bridges a critical gap in the current body of knowledge in this area of research.
Well-established animal models provide valuable information about the proteins present in the extracellular matrix during amelogenesis, both in the secretory and maturation phases. In addition to human permanent enamel, several studies have been conducted on animal models, including both erupted and non-erupted teeth. Rats and mice, with continuously growing maxillary and mandibular incisors, have become widely used animal models. This characteristic allows for the study of all stages of amelogenesis on a single incisor at any given time throughout the animal's lifespan (De Lima Leite et al. 2018). Pig teeth have also been utilized in dental research due to their availability, large size, and similarity in size and morphology to human teeth (Robinson et al. 1987; Fincham et al. 1999; Green et al. 2019; Gil-Bona and Bidlack 2020). Gil-Bona and Bidlack (2020) highlight the advantages of the porcine model for studying spatial changes within the mineralizing tooth crown and throughout the thickness of the enamel layer. The similarity between human and porcine teeth is particularly useful for modeling early stage enamel formation.
Following the maturation, cell death of ameloblast by apoptosis is a normal process during amelogenesis. Recent studies indicate that cell death during embryonic development generates several signaling molecules, which affect the behavior of adjacent cells, stimulating morphogenesis, cell migration, and alteration of cell fate during odontogenesis (Lacruz et al. 2017; Abramyan et al. 2021). MIH enamel expresses a diverse and irregular hypo-mineralization appearance pattern, indicating an impact on ameloblasts at a critical phase during amelogenesis (Sidaly et al. 2016). One might speculate that such an insult could cause an increase in protein remnants related to biological processes of cell death to be retained in the mature enamel.
Enamel development comprises several functional stages: the secretory and maturation stages, with additional subdivisions, such as presecretory, early secretory, late secretory, transition, preabsorptive, early maturation, and late maturation stages. Amelogenesis involves various activities, including the formation and subsequent removal of a proteinaceous matrix, ion transport, pH regulation, and apoptosis, resulting in avascular enamel devoid of regenerative properties (Lacruz et al. 2017; Nanci, 2008). Dental development extends beyond the mineralization of crown and root tissues, continuing until teeth reach functional occlusion postnatally. Several theories on tooth eruption exist, yet the regulatory mechanisms remain largely unknown (Wise et al. 2002). Tooth eruption occurs in three distinct stages: pre-eruptive tooth movement, eruptive tooth movement, and post-eruptive tooth movement. The second stage, eruptive tooth movement, involves the onset of root formation until the crown emerges into the oral cavity. This stage includes phases, such as intra- and supraosseous eruption, stages characterized by epithelial cell death within an inflammatory environment and bone resorption. One might speculate if proteins originating from both epithelial and inflammatory cells may persist within abnormally developed enamel, causing protein entrapment or incorporation during tooth eruption. This could potentially contribute to an elevated protein abundance in hypomineralized enamel (Kjaer, 2014; Richman 2019).
From a proteomic perspective, MIH differs from other enamel disturbances e.g., Amelogenesis Imperfecta (AI), which has shown increased residual abundance of amelogenin. The findings of this study, consistent with other research, indicate an elevated protein content in MIH enamel, but not an increased abundance of amelogenin. Serum proteins, including albumin, have been suggested as an inhibitor of apatite crystal growth (Farah et al. 2010; Mangum et al. 2010). MIH enamel is characterized by a high content of serum albumin, hemoglobin, alpha-1-anti-trypsin, anti-thrombin III, and serpin B3 (Mangum et al. 2010). Recent studies by Williams et al. (2020) and Perez et al. (2020) have proposed that proteins, such as albumin and potentially other blood-associated proteins, may act as predominant mineralization inhibitors in hypo-mineralized enamel. In the present study, serum albumin, alpha-1-anti-trypsin, Antithrombin III, and Serpin B3 were detected with no statistically significant difference observed between MIH and normal enamel. However, hemoglobin subunit gamma-2 showed an overabundance in MIH compared to normal enamel. Further investigation is warranted to explore the relationship between serum albumin, and the presence and activity of different protease inhibitors, such as anti-thrombin-III and alpha-1-anti-trypsin, in MIH enamel.
Intriguingly, the analyzed MIH groups exhibit similar trends in the presentation of biological processes, both in over- and underabundant proteins (Figs. 2–6). In both analyzed MIH groups, an underabundance of proteins primarily associated with cell organization and biogenesis was identified. Additionally, proteins linked to cell death are predominantly overabundant in both MIH groups. Mukthar et al. (2022) reported proteins, found in MIH enamel to be mainly involved in biological processes with immune and inflammatory responses. In accordance with Mukthar et al. (2022), the present study found annexin A2, keratin type II cytoskeletal 1b, keratin type II cytoskeletal 78, plakophilin-1, and small proline-rich protein to be significantly overabundant in the MIH-affected enamel. In contrast to Mukthar et al. (2022), dentin sialophosphoprotein (DSPP), a protein transiently expressed during early enamel formation, was not identified in normal, or MIH enamel.
Several keratins were detected, some at high levels, e.g., keratins 16, 27 and 71 were found to be overabundant in MIH enamel. The occurrence of keratins in enamel has been questioned. Some keratins might play an important role in the formation of tooth enamel (Duverger et al. 2014; 2016; 2019). Given that keratins are cytoskeletal proteins, further studies need to reveal if they are deposited in enamel through exocytosis or with ameloblast fragments retained in enamel (Duverger et al. 2014, 2016 and 2019; Yang et al. 2019; Deshmukh et al. 2022).
Desmoplakin, desmoglein, and desmocollin were found to be overabundant in the MIH I enamel. Desmogleins and desmocollins, both transmembrane desmosomal cadherins, maintain extracellular cell–cell adhesion. Mutations in desmoplakin may lead to enamel abnormalities (Mahoney et al. 2010; Bartlett and Smith 2013). The role of proteinases in degrading junctional complexes, as well as the remodeling of junctional complexes during normal enamel developmental processes, has only been minimally investigated. Hypothesizing defective desmosome adhesion and degradation may affect amelogenesis (Bartlett and Smith 2013). The existence of enamel non-specific proteins in both normal and MIH enamel has been reported elsewhere (Açil et al. 2005; Farah et al. 2010; Jagr et al. 2019; Mukthar et al. 2022). Whether some of these proteins could constitute a possible source of contamination due to dissection and preparation of enamel samples for protein analysis, or constitute a part of the enamel proteome, is yet to be fully explained.
This study encounters similar limitations related to the microdissection of erupted permanent enamel, as discussed in previous studies (Jagr et al., 2019; Green et al. 2019; Gil-Bona and Bidlack 2020; Rexhaj et al. 2023). In the study conducted by Rexhaj et al. (2023), the challenges posed by the highly mineralized nature of dental enamel during microdissection and protein extraction, resulting in reduced protein content and limited accessibility due to low protein abundance, are highlighted. Furthermore, the proximity of dental tissues to enamel could potentially introduce contamination during sample dissection and preparation for protein analysis. While great care was taken during sample preparation to avoid contamination, it is important to note that some degree of contamination may still affect any proteomic dataset obtained from micro-sampled tissues.
To address potential contamination issues, all enamel surfaces underwent 35% phosphoric acid etching before microdissection, thereby eliminating surface debris. The increased porosity of MIH enamel may permit the adsorption of organic compounds from dietary sources, saliva, or pellicle onto the outermost enamel surface layer. The use of phosphoric acid, as a cleaning step, primarily affects a shallow layer of the outermost enamel surface, typically spanning a few micrometers. Importantly, the sample collection was obtained at a depth of 1–2 mm in the enamel, considered not to be affected using phosphoric acid as a detergent step (Torres-Rodríguez et al. 2012).
With the discovery of an increasing number of enamel matrix components across different studies, several questions appear: which proteins are endogenous, exogenous, or essential? and how do they contribute to enamel development (Gil-Bona and Bidlack 2020)? Another challenge and limitation is the lack of a standardized protocol for sample storage prior to microdissection and protein analysis, cleaning of teeth samples, microdissection, and use of the same methodological protocols for proteomic analyses. This complicates the comparison of data across different studies. By employing a combination of microdissection methodology, validated by Rexhaj et al. (2023), and LC–MS technique, this study showcases the capability to distinguish distinct proteomic profiles in MIH-affected teeth across various clinical severity grades. In this aspect, a strength of this study lies in the meticulous sampling and preparative protocols implemented, crucial for minimizing external contaminants. An inherent limitation in the study relates to the number of measurements conducted with the LC–MS, 11-plex equipment, which was constrained to 11 samples. With this sample size, the interpretation of the results must be approached with care, underscoring the need for further data collection to enhance reliability.


留言 (0)