Most included studies demonstrated that exercise interventions can induce significant changes in DNA methylation. Trials evaluating specific CpG sites observed increased methylation related to RANKL, FKBP5, AURKA, BPIFA, BRCA1, p66Shc, and ASC genes. Regarding global methylation, studies showed significant downregulation resulting from exercise. Similarly, epigenome-wide analyses revealed exercise alters the DNA methylation profile. The vast majority of trials are in line with previous cross-sectional studies reporting significant effects of exercise on DNA methylation [7, 8, 10].
Several factors may explain the differences between findings. First, demographics such as age, sex, and ethnicity can drive heterogeneity [7, 8, 38]. Second, intervention factors including duration and intensity seem key, as 6–12 months may suffice to alter age-related genes [7], while optimal intensity remains unclear [10]. The type of exercise performed during an intervention may also be a key factor influencing epigenetic modifications. Significant changes in DNA methylation included both aerobic exercise and strength training [30, 31]. In contrast, in a study by Duggan et al. [13], participants only performed aerobic exercise training without strength training. Although the intervention lasted 12 months with a frequency of six sessions per week at moderate to vigorous intensity, no significant changes in DNA methylation were observed, suggesting that including strength training may be crucial. Similarly, in the study by Boyne et al. [25], no significant changes in DNA methylation were observed despite a duration of 12 months, frequency of three to five times per week, and moderate to vigorous intensity. Taken together, these results indicate the importance of strength training in driving changes in DNA methylation.
In addition, most studies that reported significant differences in DNA methylation also documented improvements in physical fitness. For example, in a study by Chelly et al. [30], participants in the training group showed increased DNA methylation of the RANKL gene along with enhanced maximal oxygen consumption (VO2max). Willmer et al. [31] observed significant DNA methylation changes that were accompanied by an improvement in cardiorespiratory fitness, measured as peak oxygen consumption (VO2peak). In the study by Boyne et al. [25], after correlating changes in DNA methylation with improvements in physical fitness (measured as VO2max), a significant dose–response relationship emerged between physical fitness and RASSF1 methylation in a group of inactive postmenopausal women. Hence, despite the constraints of this study suggesting that the findings should be viewed as preliminary, it seems that improved physical fitness could be a critical factor in the influence of exercise on DNA methylation.
The primary outcome of this study might account for the observed differences in results, suggesting that gene-specific effects of exercise could contribute to inconsistencies. While gene-specific studies enable hypothesis testing, observed differences may be attributable to differential responsiveness across various genes [8, 13]. For instance, Duggan et al. [13] found no significant differences in DNA methylation between groups for specific genes such as p14ARF, EVL, and ESR1. This result does not negate the possibility that exercise interventions may lead to significant DNA methylation changes in other genes, but rather indicates no observed changes in the genes under examination when compared to the control group. Most gene-focused studies have reported an increase in DNA methylation following exercise interventions [15, 26, 30,31,32]. However, studies on global DNA methylation have frequently reported a significant decrease [33,34,35]. Although this appears contradictory, it may not be, as changes in global DNA methylation reflect changes that likely encompass numerous genes, some undergoing hypermethylation and others hypomethylation. Consequently, global assessments may miss the subtleties of gene-specific effects [10]. Epigenome-wide association studies (EWAS) have identified significant variations in specific genes between exercise and control groups, but these findings include the risk of false positives due to the exploratory nature of such studies, potentially mistaking unrelated changes for effects of the intervention [34].
DNA methylation assessments also differed in tissue type. As patterns change across tissues [7, 10, 11], contrasting findings could reflect this specificity. For instance, Duggan et al. [13] found no DNA methylation changes in colon tissue, while Denham et al. [15] observed reduced sperm DNA methylation after exercise training. In addition, both studies analyzed different genes, which may indicate that exercise has effects on specific tissues and genes.
Moreover, methods used to quantify methylation differ. Pyrosequencing enables simple, rapid, and reliable analysis [39], but microarrays have higher genomic coverage [34, 35]. Within platforms, designs target varying genomic regions with different methylation statuses [8].
Reduced DNA methylation found in some studies aligns with increased DNA methylation in others when considering regulatory effects on transcription [26, 34]. Promoter methylation is associated with silencing and loss therefore enhances expression [4, 8, 10]. However, gene body methylation can increase expression, explaining the hypomethylation observed by Ngwa et al. [36] within VPS527.
4.1 Limitations
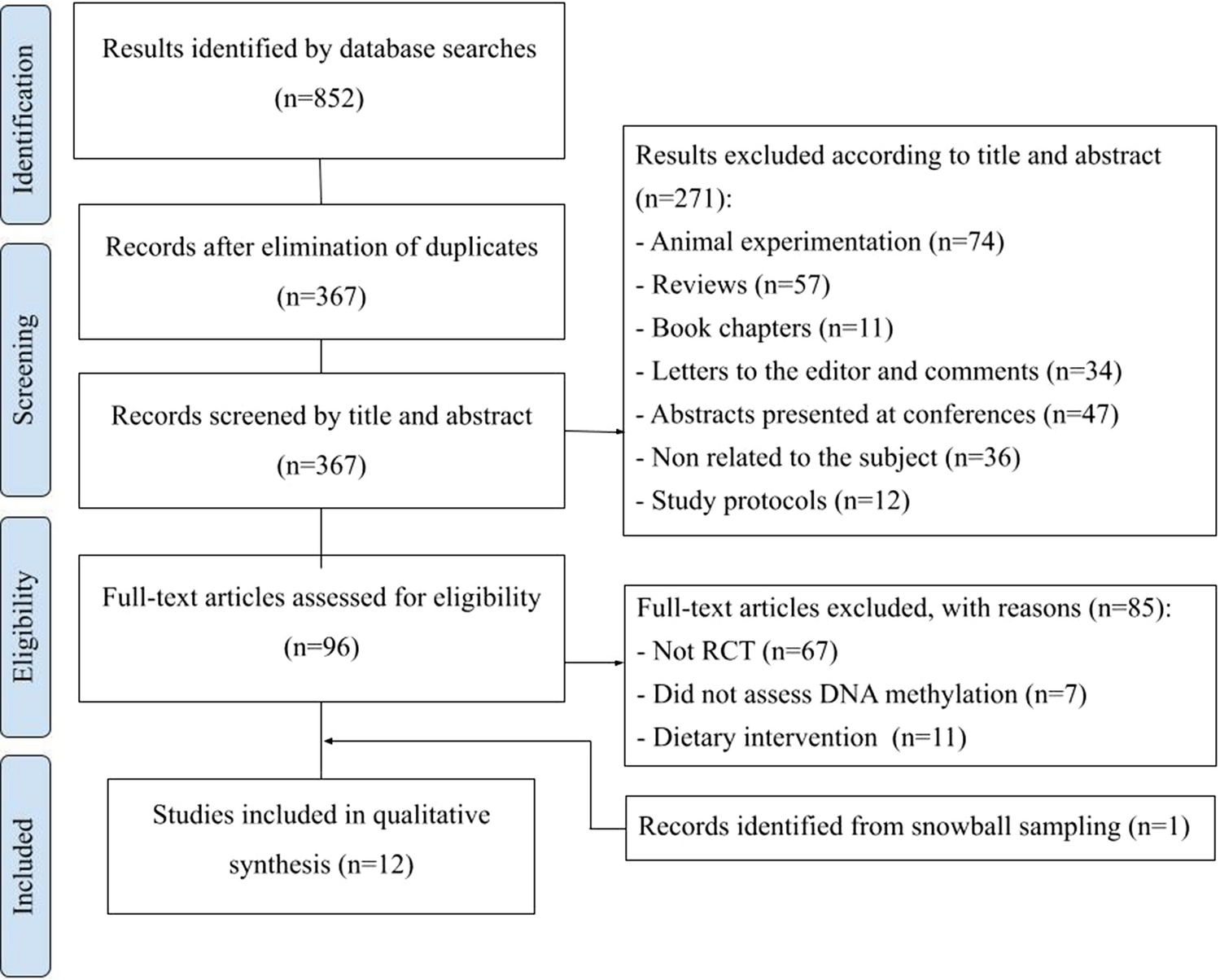
This systematic review has limitations including small sample sizes [13, 35] and predominant use of heterogeneous blood samples without cell type control [8, 15, 25]. Differing methods and tissues also complicate comparisons. While our methodological approach was in strict accordance with PRISMA 2020 guidelines and employed the PEDro scale to enhance the review's reliability and robustness, we must acknowledge certain limitations inherent to these methodologies. A significant limitation arises from the heterogeneity among the included studies, with variability in participant characteristics, intervention protocols, measurement methods, and outcome measures possibly affecting the heterogeneity and, consequently, the review's overarching conclusions. Additionally, despite meticulous data selection and extraction processes aimed at mitigating bias, the potential for inherent biases in study selection and interpretation persists. These limitations need to be considered when interpreting the review's findings, and subsequent research should strive to tackle these challenges, thereby reinforcing the evidence base in this domain. However, the review uniquely summarizes exercise intervention effects on DNA methylation using only randomized controlled trials [15], further controlling bias through inclusion of sedentary, non-smoking samples.
Though epigenetic exercise research is nascent, findings could enable use of DNA methylation to track intervention impacts on disease risk. This could motivate adherence by demonstrating behavioral change effects. However, practical applications remain speculative.
Future investigations should consider several factors. First, studies should evaluate DNA methylation based on demographics such as age and ethnicity, which can influence patterns [7]. Second, specific tissue- and cell-specificity impacts should be examined across bodily tissues, controlling for cell type distributions in blood [8]. This could reveal differentially susceptible genomic regions. While global assessments have utility, gene-specific analyses are essential for elucidating disease pathways [10]. Identifying exercise-responsive genes represents a key step for assessing epigenetic links between exercise and pathology. Additionally, comparing activity types, intensities, and durations in clinical trials is needed to elucidate dose–response relationships. Considering fitness impacts may also be valuable, as evidence suggests fitness strongly predicts DNA methylation changes [15, 38]. Analyzing potential interactions with diet and mediation by body composition represents other avenues worth exploring. Ultimately, besides randomized controlled trials, it would also be worthwhile conducting longitudinal studies, which may offer a more comprehensive understanding of the temporal dynamics of exercise-induced DNA methylation changes.
In summary, our analysis highlights the intricate relationship between exercise, DNA methylation, and prevention of chronic diseases. By identifying the key genes and genomic profiles responsive to exercise interventions, we pave the way for personalized approaches to health promotion and disease management. The insights gained from this study not only contribute to advancing our understanding of the molecular mechanisms underlying the benefits of exercise, but also hold promise for developing targeted interventions that harness the full therapeutic potential of physical activity. Continued exploration of dose–response relationships and comprehensive analysis of exercise parameters will be crucial to optimizing health outcomes and shaping public health strategies for the future. Furthermore, the identification of differentially methylated genes and pathways responsive to exercise interventions could enable the use of epigenetic biomarkers to monitor the molecular effects of exercise programs. Characterizing such profiles represents an essential step towards elucidating the mechanism of exercise.







留言 (0)