
記住我



Underlying this seemingly restored world, recovering from the spread of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), there are new plans of how to rapidly defend societies against future viral pandemics. Our main defense against pathogens is represented by the immune system that utilizes highly specialized cellular and molecular pathways to fight off infections. The interferons (IFNs) are one of the major effectors produced by the immune system to counteract foreign pathogens within the bloodstream and at mucosal surfaces. IFNs are divided into three separate families (type I, II, and III IFNs), with type II IFNs (IFN-γ) being mostly associated with defense against bacteria, and type I and type III IFNs recognized as the most potent host-derived antivirals. In humans, type I IFNs consist of several subtypes (1–13ɑ/β/ε/κ/ω), while type III IFNs comprise IFNλ1–4 (IFNλ2–3 in mice). Although structurally distinct, type I and type III IFNs share a large portion of the signaling cascade downstream of their respective receptors, IFNAR1/IFNAR2 and IL10R2/IFNLR1. The signaling events initiated by type I and III IFNs regulate the transcriptional activation of a large pool of interferon-stimulated genes (ISGs) with immunomodulatory and antiviral functions (as reviewed in Ref. [1]).
Upon respiratory virus encounter, IFNs are produced to restrict viral entry, replication, and spread. Nevertheless, SARS-CoV-2, as with all other viruses that successfully infect humans, has developed an array of strategies to promote its propagation. These strategies can target several layers of the cellular machinery that have evolved to fight an infection, for example, inhibition of host RNA translation and splicing, degradation of cellular mRNA,inhibition of nuclear transportation of mRNA, and blockade of cellular secretion of proteins 2, 3, 4, 5, 6. These processes lead to a tug of war between the host and the invading virus. In this context, the constant attempt of the immune system to counteract the evasive strategies of a virus may lead to the excessive production of immune mediators. Extensive research has shown that these immune mediators can contribute to dysfunction of both the lung and other tissues or organs in the body. This is also true for the most severe form of the disease initiated by SARS-CoV-2, which is characterized by high levels of proinflammatory chemokines and cytokines, with IFNs being no exception and playing dual and often opposing roles in coronavirus disease 2019 (COVID-19) (as reviewed in Ref. [7]). In this review, we focus on the diverging roles of IFNs across ages during the acute phase of SARS-CoV-2 infection, as well as on the role of IFNs during long COVID-19. We will also emphasize what we have learned from targeted IFN therapies during the COVID-19 pandemic.
From the first study shedding light on the inflammatory nature of severe COVID-19 [8], countless research papers have strived to understand the roles played by IFNs in the inflammatory process elicited upon SARS-CoV-2 encounter ([9] and as reviewed in Refs. 10, 11). Altogether, these papers show how the inflammatory profile of patients in blood and distinct anatomical compartments correlates to the severity of the disease. Patients with severe COVID-19 are characterized by increased production of proinflammatory cytokines and reduced type I/III IFN activity in the blood, at least upon early SARS-CoV-2 infection 8••, 12••. In keeping with a protective role of IFN in the blood, systemic type I IFN and ISGs are induced in patients with mild or moderate disease severity [13]. It is, though, important to consider both the timing and location of these measurements. In a longitudinal analysis, increased type I, II, and III IFNs, and/or ISGs, have been found in the blood of patients with severe disease [12]. This is most likely a consequence of persisting viral recognition since it correlates to the viral levels in patients with severe disease [14].A recent study also showed how a high IFN response early during SARS-CoV-2 infection may hamper development of antibody responses against the virus, increasing pathology and severity of disease in patients with high levels of inflammation and viremia. Similary to humans infected with SARS-CoV-2, mouse models of COVID-19 showed highly disparate results. Herein, one model has shown that type I IFN signaling is required to restrain viral replication early during the infection, while two different mouse models of SARS-CoV-2 infection showed that this family of IFNs plays detrimental roles 15, 16, 17, 18. Additionally, several studies have shown that airway epithelial cells (AECs) respond to SARS-CoV-2 and its variants of concern (VOCs) in a highly dynamic and transient fashion. The early response of AECs is dominated by inflammatory and immunomodulatory changes, while the later response is characterized by the high production of type I/III IFNs 19, 20. Even though systemic and mucosal responses of patients with a sustained infection are dominated by IFN-mediated alterations, the relative production and kinetics of the different IFNs are highly variable [21]. The landscape of IFNs, ISGs, and transcriptional changes also varies along the respiratory tract and according to disease severity 22•, 23•. Exactly how the IFN response contributes to the inflammatory process is, thus, a complicated matter. This complexity is driven by the (largely unknown) nature of the cells producing and/or responding to IFNs, where the response takes place, the presence of inherent genetic deficiencies, or autoantibodies attenuating immunological functions 24••, 25•, 26, 27•, 28, 29, 30•, 31, 32. Another important element is the capacity of proteins produced by SARS-CoV-2 to impede and/or dampen the antiviral mechanisms of the host (as reviewed in Ref. [33]). In this regard, it is important to note that even if SARS-CoV-2 can inhibit the induction of IFNs, inhibition of IFNs by viral effectors is not efficient when the viral burden increases 22•, 23•, implying that there is a threshold for the suppression of the IFN response.
Overall, these studies suggest that the timing and anatomical distribution of IFN production are fundamental to determining the host response upon the encounter with SARS-CoV-2. In the rest of this review, we will summarize the multiple roles played by IFNs in COVID-19 and highlight the lessons learned from clinical trials based on the use of recombinant IFNs as therapeutic agents against COVID-19.
The importance of IFNs in protecting against SARS-CoV-2 is strongly supported by the genetics of patients with severe disease . Genetic defects in IRF-9, IRF-7, TLR3, and TLR7, which impair both type I and III IFN production and downstream signaling, are over-represented in patients with severe COVID-19 25•, 31, 34••, 35, 36. Of note, X-linked chromosomal defects in TLR7 may also at least partially explain the bias in COVID-19 severity depending on sex (as reviewed in Ref. [37]), although other immunological, as well as behavioral factors may also contribute (as reviewed in Ref. [38]).
The importance of blocking the induction and/or signaling of multiple IFN families with regard to increased susceptibility to SARS-CoV-2, as well as other viruses, has also been recently confirmed for patients with STAT2 deficiency [30]. Nevertheless, it is important to note that genetic defects in IFNAR2, which only affect type I IFN signaling, have also been described 39, 40. Although these genetic defects are only detected in a small percentage of patients with life-threatening COVID-19, additional studies have shown that 10–20% of patients with severe disease have autoantibodies to IFN-ɑ, -λ, -ω, and to a lesser extent -β subtypes 24••, 26, 28, 29, 34••, 41, 42. Overall, underlying human genetic defects to the type I and III IFN systems, or the presence of IFN autoantibodies that mimic such immune deficiencies, may cause an insufficient defense against SARS-CoV-2 infection in some patients, thus facilitating the development of severe disease (as recently reviewed in Refs. 43, 44).
Age is one of the main determinants of COVID-19 severity and mortality 45, 46. In terms of age as a factor that determines COVID-19 severity, it is well established that immunological functions decrease as individuals get older. Similarly, mortality during the first wave of the COVID-19 pandemic was increased in the older adult population, while children infected with SARS-CoV-2 generally exhibited reduced severity during the infection. Notably, the correlation between the viral load and IFN expression is significant only in individuals <70 years of age, while it is lost in patients >70, demonstrating how the host natural antiviral mechanisms are dysregulated in older patients [23]. In keeping with alterations in the immune response of elderly, the presence of autoantibodies to type I IFNs also increases with age (>70 years), being prevalent already before the infection with SARS-CoV-2, and contributing to disease severity 26, 47. However, the overall increased incidence of severe SARS-CoV-2 infection in the older population can also be due to reduced immunological functions that go beyond IFNs and to the increased incidence of comorbidities (as reviewed in Ref. [38]).
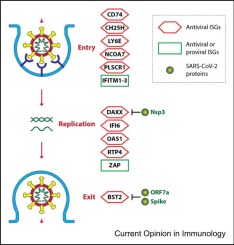
To understand the mechanisms underlying the protective or detrimental roles driven by IFNs, it should be taken into consideration that distinct IFN-λ subtypes have the capacity to induce a unique set of ISGs. The relative production of type I/III IFNs from cells of immune or mucosal origin should also be more thoroughly analyzed. The type of IFNs that are produced, the timing of their production, and their location along the respiratory tract also play important roles in determining the activity of IFNs (summarized in Figure 1a). Overall, exactly how type I/III IFNs mediate their effects during the SARS-CoV-2 infection in particular, and during lung virus infections in general, remains to be better elucidated.
Pediatric patients are generally less susceptible to severe forms of acute COVID-19. Nevertheless, a recent study showed that high levels of anti-IFN autoantibodies are rare but can occur in children and adolescents who have inborn errors of immunity, leading to a spectrum of responses to SARS-CoV-2. A patient with NF-κB2 haploinsufficiency showed high levels of anti–IFN-ɑ2 antibodies [48], while higher levels of autoantibodies against type I IFNs are detected in individuals with autosomal recessive NIK/gene name, no acronym or autosomal-dominant NF-κB2 deficiency [49]. These studies further support the tight link between the presence of high IFN autoantibodies and underlying inborn errors of immunity, as it also happens in autoimmune polyendocrine syndrome type 1 [43].
In children, milder manifestations of SARS-CoV-2 could be a consequence of higher type III IFN responses compared to adults, with both IFN-λ1 and IFNλ2/3 showing an inverse correlation with age [50], although another study showed that type III IFN levels are comparable irrespective of severity status [51]. Other than IFN levels, the strong IFN response of nasal epithelial cells of children can be caused by these cells being primed for cytokine production, possibly mediated by an increased crosstalk between immune and mucosal cells 52, 53. Additionally, together with high levels of type I and III IFNs, infants exhibit a more rapid induction of nasal mucosal immunity, while antibody titers against SARS-CoV-2 subtypes decay at a much slower rate than in adults [54]. Interestingly, both asymptomatic and nonsevere children have higher levels of activated natural killer (NK) cells compared to adults, although NK cells exhibit lower expression levels of killer-cell immunoglobulin-like receptors [50]. Of note, the activity of NK cells is tightly regulated by type I IFNs that are necessary to induce the cis- or trans-presentation of IL-15 55, 56. Since several studies have shown that type III IFNs can reduce SARS-CoV-2 replication while also inducing an efficient ISG response 57, 58••, the concept of accelerated clearance and reduced severity in pediatric patients is of interest and needs further research (Figure 1b). Finally, a recent publication demonstrated how nasal epithelial cells from children are intrinsically more capable to produce type I and type III IFNs compared to epithelial cells from the nose of aged indivduals that instead showed more pro-fibrotic and reparative programs [59].
Multiple lines of evidence suggest that exogenous IFNs can be used therapeutically against SARS-CoV-2. In animal models, both recombinant type I and III IFNs, or induction of IFNs by the use of synthetic ligands, have been successfully utilized to prevent SARS-CoV-2 infection when administered before or immediately after virus encounter 60, 61, 62, 63. Of note, the intranasal administration of type III IFN specifically showed protection against multiple SARS-CoV-2 VOCs [61]. Intranasal inoculation of IFN-λ in a Syrian hamster model has also been shown to reduce SARS-CoV-2 replication in the lung, production of inflammatory cytokines, and lung pathology [57]. Additionally, in air–liquid interphase models of the nasal mucosa, it has been shown that paracrine induction of type I and type III IFNs upon SARS-CoV-2 infection counteracts the spread of the virus. When these nasal cells were pretreated with IFN-β or IFN-λ1, the viral replication was reduced [64]. At the single-cell level, distinct IFN-⍺ subtypes have been shown to induce unique patterns of ISGs in models of cells infected with SARS-CoV-2. Amongst the 12 IFNs tested, IFN-ɑ5 showed the best antiviral capacity both in vitro and in vivo in mice [65]. Overall, these data show that distinct IFNs diversely control SARS-CoV-2.
The importance of distinct IFNs or IFN-dependent signaling in controlling SARS-CoV-2 in epithelial cells during natural infection has also been confirmed at the single-cell level in patients with COVID-19 [22]. Only patients who developed a mild disease were characterized by high levels of ISGs in cells of the nasopharyngeal mucosa [22]. Another study revealed a detailed map of the production of distinct members from the three IFN families along the respiratory tract, based on disease severity. High levels of type III IFNs (in particular of IFN-λ1 and IFN-λ3, but not IFN-λ2) and restricted members of the type I IFN family were upregulated in the upper airways of patients with mild disease. In contrast, patients with severe disease presented with high levels of IFN-λ2 and multiple type I IFN members in both the upper and lower airways [23]. Interestingly, in lung autopsies from patients deceased following SARS-CoV-2 infection, type III IFN levels were completely abolished, while IFN-β levels were high [66]. Additionally, the treatment with IFN-γ, but no other IFNs, and tumor necrosis factor-⍺ leads to an inflammatory-induced cell death called PANoptosis that can be involved in increasing the inflammatory response that characterizes patients with severe COVID-19 [67]. Together, this data further stress that not all IFNs are equal and that distinct IFNs at specific anatomical sites may be able to drive opposite functions.
An explanation for the different activity of distinct IFNs may lie in their capacity to induce unique sets of ISGs, as demonstrated for type III IFNs [23]. As for IFNs themselves, not all the ISGs equally control SARS-CoV-2. For example, viral clearance is enhanced in individuals of European ancestry that express a specific isoform of the IFN-inducible gene 2'-5' oligoadenylate synthetase 1 (OAS1) [68]. Furthermore, a clustered regularly interspaced short palindromic repeats screen showed that OAS1, specifically compared to OAS2/3, is a major antiviral factor against SARS-CoV-2 [69]. OAS1 exists in a prenylated isoform, associated with the endomembrane system, which is capable of efficiently detecting the presence of SARS-CoV-2 RNA and is associated with protection from severe disease [70]. In patients who have genetic impairment of OAS1, clearance of the virus can be restored by treating with pegylated type III IFN (peg-IFN-λ1) [71]. In contrast to these data, a small study (limited to 5 patients with COVID-19) showed that the OAS family members are enriched, together with type I IFN levels, in the nasopharyngeal swabs of patients with severe COVID-19 disease [72]. Overall, these data reveal the importance of OAS family members during COVID-19 and suggest a fine-tuning of their activity in response to the infection with SARS-CoV-2.
Although determining the causality between the landscape of IFNs and the severity of COVID-19 in patients may require more analyses, mouse studies have demonstrated that prolonged IFN signaling in the lower airways is associated with proapoptotic signals that disrupt the barrier activity of the lung 73••, 74. Although it may be counterintuitive that IFNs play opposing roles during lung viral infections, it is well known that a major driver of severe COVID-19 is the excessive production of immune mediators, often referred to as cytokine storm (although this term is still controversial in COVID-19 [75]). It is, thus, not surprising that IFNs can also play, among others immune mediators, a dichotomic role in the response to SARS-CoV-2. For example, the STAT2 deficiency described above, which impairs IFN responses, leads to excessive inflammation during the early stages of the infection [30]. In contrast, excessive inflammation can be a result of hyperactivation of macrophages upon sensing of type I IFNs produced by plasmacytoid (p)DCs following TLR7 activation 76, 77. Once again, rather than the presence or absence of IFNs, it is their tight regulation in space and time (see below) that determines the outcome of COVID-19.
The timing of IFN production appears to be crucial during SARS-CoV-2 infection. An early IFN activity is essential for appropriate innate antiviral immune suppression, while critical COVID-19 patients have reduced blood levels of type I IFN and associated ISGs [78]. Activation of type I IFN signaling in hospitalized patients induces a transcriptomic response associated with inflammation, not seen in healthy or nonhospitalized individuals [78]. Additionally, in patients hospitalized with severe disease, an ineffective type I IFN production and/or secretion can be partially attributed to decreased frequency of circulating pDC and monocytes and to the promotion of an inflammatory response in leukocytes [78]. However, during the infection with SARS-CoV-2, the activation of pDC and the production of IFNs are intact, suggesting that the ineffective production of type I IFN during severe disease is most likely a consequence of reduced cell frequencies 78••, 79. Additionally, a recent study showed that treatment with a modified IFN-⍺2 (IFNmod), which restrained endogenous signaling of type I IFN, reduced the viral load in both primary human AECs and in rhesus macaques (RM) [80]. Of note, administration of IFNmod to uninfected RM induced the production of antiviral ISGs.
Overall, these studies indicate that IFN signaling during SARS-CoV-2 infection can sustain a proinflammatory milieu that induces aberrant cell functions and promotes a severe disease.
Since the implementation of successful vaccination strategies against SARS-CoV-2, the severity, adversity, and the imminent threat of COVID-19 to our societies significantly decreased. However, unforeseen complications have risen, such as long COVID-19, also known as postacute sequelae of COVID-19 (PASC). In this condition, variable debilitating symptoms of fatigue, memory/concentration decline, autonomic dysfunction, or flu-like manifestations do not resolve in individuals for months or even years after SARS-CoV-2 seroconversion. Different biological factors are suggested to drive long COVID-19, for example, persisting virus/viral RNA in different anatomical tissue compartments, reactivation of neurotrophic viruses, organ damage, alterations in inflammatory profile of immune subsets, autoimmunity, dysbiosis of the microbiota, and endothelial cell dysfunction ([81] and as reviewed in Refs. 82, 83, 84). Development of long COVID-19 is more common in women, highlighting the role of underlying immunological functions that differ between the sexes [85], although the developmental process and consequences of long COVID-19 remain to be elucidated. At the immunological level, the cellular and cytokine profile in the peripheral blood is changed in individuals with long COVID-19. Decreased frequencies of cDC1s and memory CD4+ T cells are characteristic of these patients, while nonconventional monocytes and double-negative B cells are increased, together with antibody levels against SARS-CoV-2, Epstein–Barr virus, and varicella zoster virus (VZV) antigens [81]. Additionally, after seroconversion, some inflammatory cytokines, together with levels of IFN-λ1 and IFN-β, are elevated in individuals experiencing long COVID-19 86, 87. Of note, type II IFN has also been shown to be increased in the peripheral blood of patients with long COVID-19, as a consequence of CD14+ myeloid cell–dependent activation of CD8+ T cells [88]. The chronic inflammatory profile of patients with long COVID-19 can be a consequence of prolonged activation of pDCs that sustain the production of IFNs. As in acute COVID-19, autoantibodies directed against IFNs may also play an important role. In patients with long COVID-19, early autoantibodies to IFN-⍺2 inversely correlate to SARS-CoV-2 antibodies and are associated with prolonged symptoms [89]. Although the development of long COVID-19 does not seem to be correlated to the severity of acute disease, several biological or immunological parameters can be measured during the acute phase of the infection to predict individuals at risk of developing long COVID-19 86, 89, 90, 91, 92, 93, 94. Identifying specific traits and factors as biomarkers will aid in the identification of individuals who are likely to develop long COVID-19 and in the implementation of early intervention strategies to minimize the development of long COVID-19 (Figure 1c). Finally, long COVID-19 can possibly be subdivided into an inflammatory or noninflammatory subgroup. These subgroups are distinguished by high neutrophil activity, altered B-cell responses, together with increased type II IFN and NF-κB signaling 87, 95. Taken together, these papers suggest that an early, strong induction of type I/III IFNs during acute infection can play a role in the development of long COVID-19, but only if inflammation is sustained and becomes chronic. However, further studies are needed to elucidate the timing, location, and how unique subtypes of IFNs might play a role in the development and maintenance of long COVID-19.
As reported above, initial studies suggested that SARS-CoV-2 has an increased capacity to shut down IFN induction compared with other respiratory viruses [8]. Nevertheless, numerous studies have highlighted the capacity of the host to efficiently produce IFNs upon SARS-CoV-2 encounter. Specific cell types appear to have distinct IFN signatures [96], and in vitro studies have shown how both type I and III IFNs can inhibit SARS-CoV-2 replication 97, 98. However, translating the in vitro findings to interventional strategies against COVID-19 based on the administration of exogenous IFN has proved complicated, as described below. This may be due to the fact that the anti–SARS-CoV-2 activity of IFNs, especially in vivo, may require unique subsets of IFNs acting at different times and anatomical sites, rather than the generic presence of IFNs as extensively discussed above.
Since IFNs are already used to treat a variety of diseases (as summarized in Ref. [99]) and their safety profile in humans is well known, prophylactic and therapeutic clinical trials against SARS-CoV-2 infection were rapidly set up (see Figure 2a). In terms of type I IFNs, both subcutaneous and inhaled IFN-ɑ administration have shown to reduce viral shedding, duration, and symptoms of COVID-19 disease alone or together with antivirals, as discussed below (and reviewed in Ref. [100]). One study showed that early administration of inhaled IFN-β in hospitalized patients contributed to a more rapid recovery and higher odds of improvement [101]. However, in a follow-up phase III clinical trial, nebulized IFN-β did not show any beneficial effect in terms of recovery [102]. Prophylactic intranasal administration of IFN-ɑ2a/IFN-ɑ2b, together with personal protective equipment, has also been shown to reduce the risk of infection in a cohort of healthcare workers in China [103]. Nevertheless, a recent meta-analysis including 11 trials of IFN-ɑ treatment showed no benefit in survival or length of hospitalization in patients with moderate to severe COVID-19 [104]. Similarly, there has been a lack of efficacy of subcutaneous IFN-β treatment in reducing the duration, clinical course, or transmission of COVID-19 105, 106, 107. Treatment with IFN-β in combination with antivirals/immunosuppressive drugs (i.e. lopinavir, ritonavir, atazanavir, ribavirin, and hydroxychloroquine) have shown varied results in hospitalized patients. Some studies have shown that IFN-β in combination with a variety of these drugs can alleviate symptoms, reduce viral shedding, increase 14-day discharge, or reduce the 28 days of mortality 108, 109, 110. In contrast, other studies have found no beneficial effects 111, 112. Surprisingly, one study even showed how IFN-β in combination with remdesivir worsened clinical outcomes in patients who required high-flow oxygen levels at baseline [113]. Therefore, interpretations of the beneficial role of IFN-β are complicated and require further investigations.
Clinical trials that are still ongoing, or that are completed but whose results are still pending, are as follows: i) assessing the use of nasal IFN-γ for preventing acute respiratory infections (NCT05054114); ii) testing the therapeutic potential of subcutaneous injection of peg-IFN-β1a (NCT04647669); or iii) evaluating whether nasal administration of IFN-⍺ can prevent infection in patients with cancer with known/unknown exposure to COVID-19 (NCT04534725) (see Figure 2b for a detailed list of clinical trials). Additionally, there are multiple ongoing or completed trials whose results are still pending, which are focused on evaluating the combined treatment with antiretrovirals and different administration routes of IFN-⍺ or IFN-β. Overall, the studies available so far suggest that prophylactic or early treatment with type I IFN upon SARS-CoV-2 infection is beneficial. In contrast, the administration of type I or type II IFNs later after SARS-CoV-2 encounter has not yet shown a significant effect, or even showed an increased risk of death, as previously reported [114].
Besides type I or type II IFNs, the possibility to utilize type III IFNs against SARS-CoV-2 has raised a lot of enthusiasm since the beginning of the pandemic [115]. Unfortunately, some clinical trials aimed at evaluating the prophylactic or therapeutic potential of pegylated type III IFN have been terminated as they were unable to meet the enrollment goals (NCT04343976 and NCT04344600), while other trials showed no or limited protection 116, 117. Subcutaneous administration of peg-IFN-λ1 increased the rate of viral decline in patients with COVID-19, while respiratory symptoms also improved at a faster rate in patients with high viral load [117]. However, peg-IFN-λ1 administration within 72 hours of diagnosis did not improve symptoms or reduce viral shedding compared to placebo in a cohort of uncomplicated COVID-19 disease [116]. Notably, peg-IFN-λ1 administration induces an antiviral ISG response in immune cells expressing IFNLR1 while not affecting the levels of SARS-CoV-2 specific antibodies or virus-specific T cells [58]. A more recent trial, the TOGETHER study, that predominantly involved vaccinated adults with SARS-CoV-2 infection in Brazil and Canada, found that the incidence of an emergency department visit or hospitalization was significantly lower among those who received a single dose of peg-IFN-λ1 compared to individuals who received placebo [118]. Still, there are two clinical trials awaiting completion/publication of results assessing the effect of subcutaneous infection of peg-IFN-λ1 as a therapeutic option against the infection with SARS-CoV-2 (NCT04967430 and NCT04534673). Overall, these data highlight the complexity of timing, dosage, and distribution of IFNs when evaluating efficacy in clinical trials.
留言 (0)