
記住我







According to the World Health Organization, the most leading cause of deaths worldwide in 2019 was ischemic heart disease (16%), also called coronary heart disease. In Japan, heart disease was the second leading cause (14.8%) of death after cancers (24.6%) in 2022, and the prevalence of heart disease is currently increasing. Coronary heart disease, including angina pectoris and myocardial infarction, is caused by the atherosclerosis of the coronary arteries. The development of atherosclerosis involves many pathogenic processes, including lipoprotein subendothelial retention, modification and aggregation, macrophage chemotaxis, and foam cell formation [1]. Atherosclerosis is induced by the subendothelial retention of atherogenic lipoproteins containing apolipoprotein (apo) B, such as low density lipoproteins (LDL), lipoprotein remnants, and lipoprotein (a) (Lp(a)) [1, 2]. In the subendothelium, apoB-containing lipoprotein particles are modified by phospholipid (PL) hydrolysis, oxidation, proteolysis, glycosylation, aggregation, or complexation with proteoglycans [1]. Macrophage uptake of modified lipoproteins, but not native LDL, induces the formation of foam cells that accumulate cholesteryl esters (CEs). In contrast, high density lipoprotein (HDL) particles have atheroprotective functions and mediate the efflux of excess cholesterol from foam cells in atherosclerotic lesions [3].
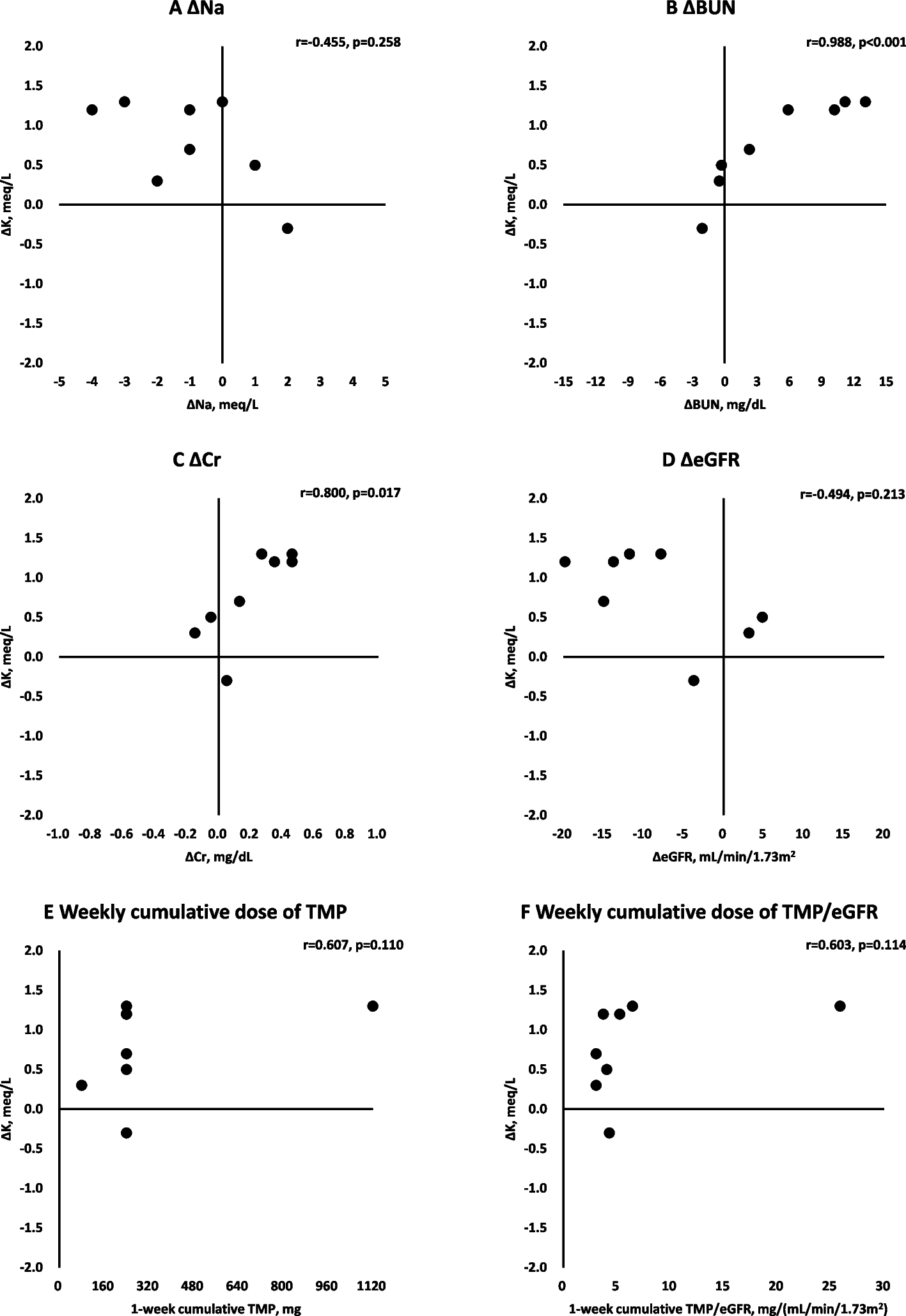
Lipoproteins and coronary heart diseaseIn the plasma lipoprotein structures, a hydrophobic core composed of triglycerides (TGs), also called triacylglycerols, and CEs is surrounded by a surface monolayer composed of PLs, unesterified (free) cholesterol (FC), and apolipoproteins (Fig. 1) [1]. Plasma lipoproteins are classified into HDL (diameter 5–12 nm, density 1.063–1.210 g/mL), LDL (diameter 18–25 nm, density 1.006–1.063 g/mL), very low density lipoprotein (VLDL) (diameter 30–80 nm, density 0.94–1.006 g/mL), and chylomicron (CM) (diameter 75–1,200 nm, density < 0.94 g/mL) (Fig. 2) [3]. Each LDL particle or VLDL particle contains one molecule of apoB100, and each CM particle contains a single molecule of apoB48. In contrast, HDL particles primarily contain apoA-I but not apoB. CM particles also contain ApoA-I. Lp(a) (diameter ~ 25 nm, density 1.05–1.1 g/mL) is an LDL-like particle containing apo(a) covalently bound to apoB-100 [4].
Fig. 1
Model of plasma lipoprotein structure. A hydrophobic core consisting of TGs and CEs is surrounded by a surface monolayer composed of PLs, FC, and apolipoproteins. CE, cholesteryl ester; FC, free cholesterol; PL, phospholipid; TG, triglyceride
Fig. 2
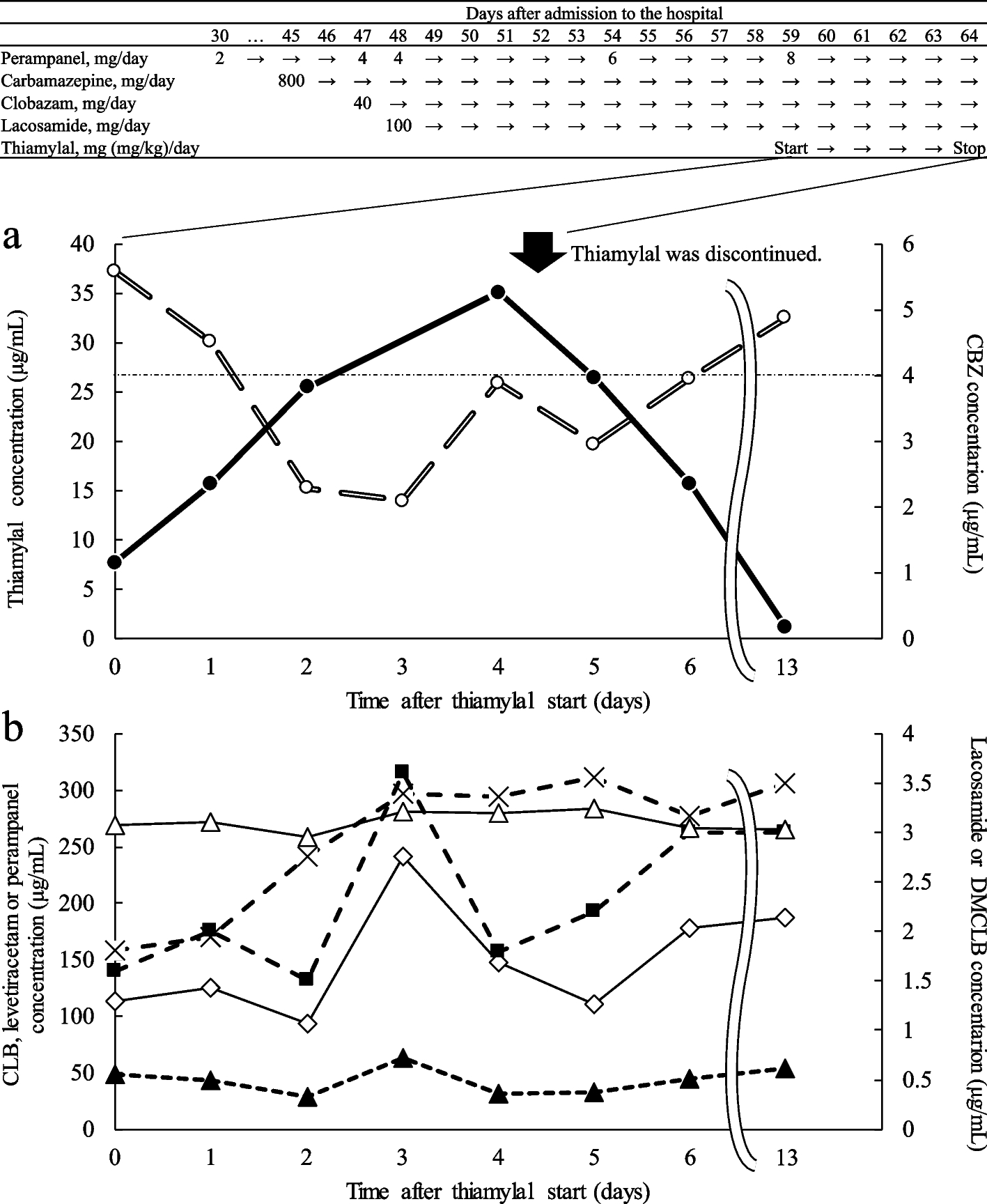
Metabolism of plasma lipoproteins. CM (diameter 75–1,200 nm) and VLDL (diameter 30–80 nm) are formed in the intestine and liver, respectively, in which TG and PL are carried to apoB by MTP. In the plasma, lipolysis by LPL mediates the conversion of CMs to CM remnants and that of VLDLs to VLDL remnants and subsequently to LDLs (diameter 18–25 nm). CM and VLDL particles are rich in TGs, and LDL particles are rich in CEs. CM remnants, VLDL remnants, and LDLs are removed from circulation by the liver. In the formation of HDL in peripheral tissues, the cellular efflux of PLs and FC is mediated by apoA-I and ABCA1. Through esterification of FC by LCAT, pre-β-HDL (diameter 5–7 nm) is converted to HDL3 (diameter 7–9 nm) and HDL2 (diameter 9–12 nm). CE, cholesteryl ester; CM, chylomicron; FC, free cholesterol; HDL, high density lipoprotein; LCAT, lecithin:cholesterol acyltransferase; LDL, low density lipoprotein; LPL, lipoprotein lipase; MTP, microsomal triglyceride transfer protein; PL, phospholipid; TG, triglyceride; VLDL, very low density lipoprotein
CM and VLDL particles are assembled in enterocytes and hepatocytes, respectively, where TG and PL molecules are transferred to apoB by microsomal TG transfer protein (MTP) (Fig. 2) [1]. The PL molecules in the CM and VLDL particles are derived from intracellular membranes, particularly the endoplasmic reticulum membrane. In the circulation, TG hydrolysis, also called lipolysis, by lipoprotein lipase (LPL) is activated by apoC-II on lipoprotein particles and mediates the conversion of CMs to CM remnants and that of VLDLs to VLDL remnants and subsequently to LDLs. The CM and VLDL particles are TG rich, whereas LDL is rich in CEs. The PL molecules in lipoprotein particles are hydrolyzed by hepatic TG lipase and LPL [5]. ApoC-II is responsible for the activation of LPL [6]. LDL particles and lipoprotein remnants are removed from circulation by the liver. ApoE on lipoprotein remnants enhances the binding to heparan sulfate proteoglycans on the hepatocyte surface and their uptake into the hepatocytes through LDL receptor or LDL receptor-related protein [1, 7, 8]. LDL receptor interacts with apoB-100 and apoE, but not apoB-48. Proteins on LDL particles are almost exclusively apoB-100, which promotes their internalization into the hepatocytes via LDL receptor [1]. Proprotein convertase subtilisin-kexin type 9 (PCSK9) binds to LDL receptor and prevents the recycling of internalized LDL receptor to the cell surface, leading to the intracellular degradation of LDL receptor [9]. ApoC-III inhibits lipolysis mediated by LPL and hepatic uptake of lipoproteins mediated by apoE or apoB-100 [1, 6,7,8]. The binding of apolipoproteins and enzymes to the lipoprotein surface is regulated by the PL composition of the surface monolayer, which determines lipoprotein metabolism.
During the formation of HDL particles containing apoA-I, the efflux of PLs and FC from the cell surface plasma membrane is mediated by the interaction of apoA-I with the ATP-binding cassette transporter ABCA1, which is the initial step of the reverse cholesterol transport pathway (Fig. 2) [10, 11]. The nascent form of HDL, pre-β-HDL (diameter 5–7 nm, density > 1.210 g/mL), includes discoidal particles containing two apoA-I molecules [3, 12]. By the action of lecithin:cholesterol acyltransferase (LCAT), pre-β-HDL particles are converted to spherical α-HDL particles, HDL3 (diameter 7–9 nm, density 1.125–1.210 g/mL) and HDL2 (diameter 9–12 nm, density 1.063–1.125 g/mL), and most of plasma HDL are spherical α-HDL particles [12]. On HDL particles, LCAT catalyzes the formation of CEs by the transesterification of a fatty acid from the sn-2 position of PC to the 3-hydroxyl group of FC [12]. The transformation of smaller HDL3 particles to larger HDL2 particles is promoted by the esterification of FC by LCAT. ApoA-I serves as the main activator of LCAT on the HDL surface [3, 12]. Cholesteryl ester transfer protein (CETP) facilitates the transfer of CEs from HDL to apoB-containing lipoproteins and that of TGs from apoB-containing lipoproteins to HDL [12]. Phospholipid transfer protein mediates the transfer of PLs from apoB-containing lipoproteins to HDL and that between HDL particles [3]. CEs and FC in HDL particles are transferred to hepatocytes through scavenger receptor B1-dependent selective uptake but not through whole particle uptake [3, 12].
The incidence of coronary heart disease is positively correlated with LDL-cholesterol levels, but negatively correlated with HDL-cholesterol levels [13]. The levels of LDL-cholesterol and HDL-cholesterol represents the sum of the concentrations of FC and CEs in LDL and HDL, respectively. Familial hypercholesterolemia, caused by a mutation in LDL receptor, is characterized by the elevated levels of LDL-cholesterol [1]. Tangier disease is characterized by a complete deficiency of ABCA1 function and extremely low levels of HDL [3, 10]. 3-Hydroxy-3-methylglutaryl-coenzyme A reductase inhibitors, such as pravastatin, simvastatin, fluvastatin, atorvastatin, pitavastatin, and rosuvastatin, lower LDL-cholesterol levels and reduce the risk of coronary heart disease [14,15,16,17,18,19,20]. Evolocumab, a monoclonal antibody against PCSK9, lowers the levels of LDL-cholesterol and Lp(a), and prevents cardiovascular events [21, 22]. In addition, the small interfering RNA inclisiran inhibits the hepatic synthesis of PCSK9 and reduces the level of LDL-cholesterol [23]. In patients with homozygous familial hypercholesterolemia, the MTP inhibitor lomitapide lowers LDL-cholesterol, VLDL-cholesterol, and plasma TG levels [24, 25]. The CETP inhibitor anacetrapib markedly increases HDL-cholesterol levels, but only slightly reduces coronary events [26]. The high level of plasma TG is a weak risk factor for coronary heart disease compared to the high level of LDL-cholesterol or the low level of HDL-cholesterol [27]. In fact, a potent selective peroxisome proliferator-activated receptor α modulator, pemafibrate, is ineffective in reducing the incidence of cardiovascular events despite the marked reduction in TG levels [28, 29]. In contrast, the relationships of PLs in plasma lipoproteins with coronary heart disease are not well understood.
Phospholipid molecular structuresPL, consisting of two hydrophobic acyl chains and a hydrophilic head group, is an amphiphilic molecule comprising surface monolayers of plasma lipoprotein particles. In mammals, there are two groups of PLs: glycerophospholipids (GPLs) containing a glycerol backbone and sphingophospholipids (SPLs) containing a sphingosine backbone [30]. Various long-chain saturated, monounsaturated, and polyunsaturated fatty acids are esterified into PL molecules (Figs. 3 and 4). The chain length of fatty acids linked to PLs varies from 14 to 24 carbon atoms. In addition to chain length, fatty acids are classified according to the number, position, and stereochemistry (cis or trans) of their double bonds. Most of the double bonds in fatty acids are cis. For example, linoleic acid, also called 9,12-octadecadienoic acid, is denoted as 18:2(n-6), which means 18-carbon chain with 2 double bonds and the first double bond in the 6th position from the methyl end.
Fig. 3
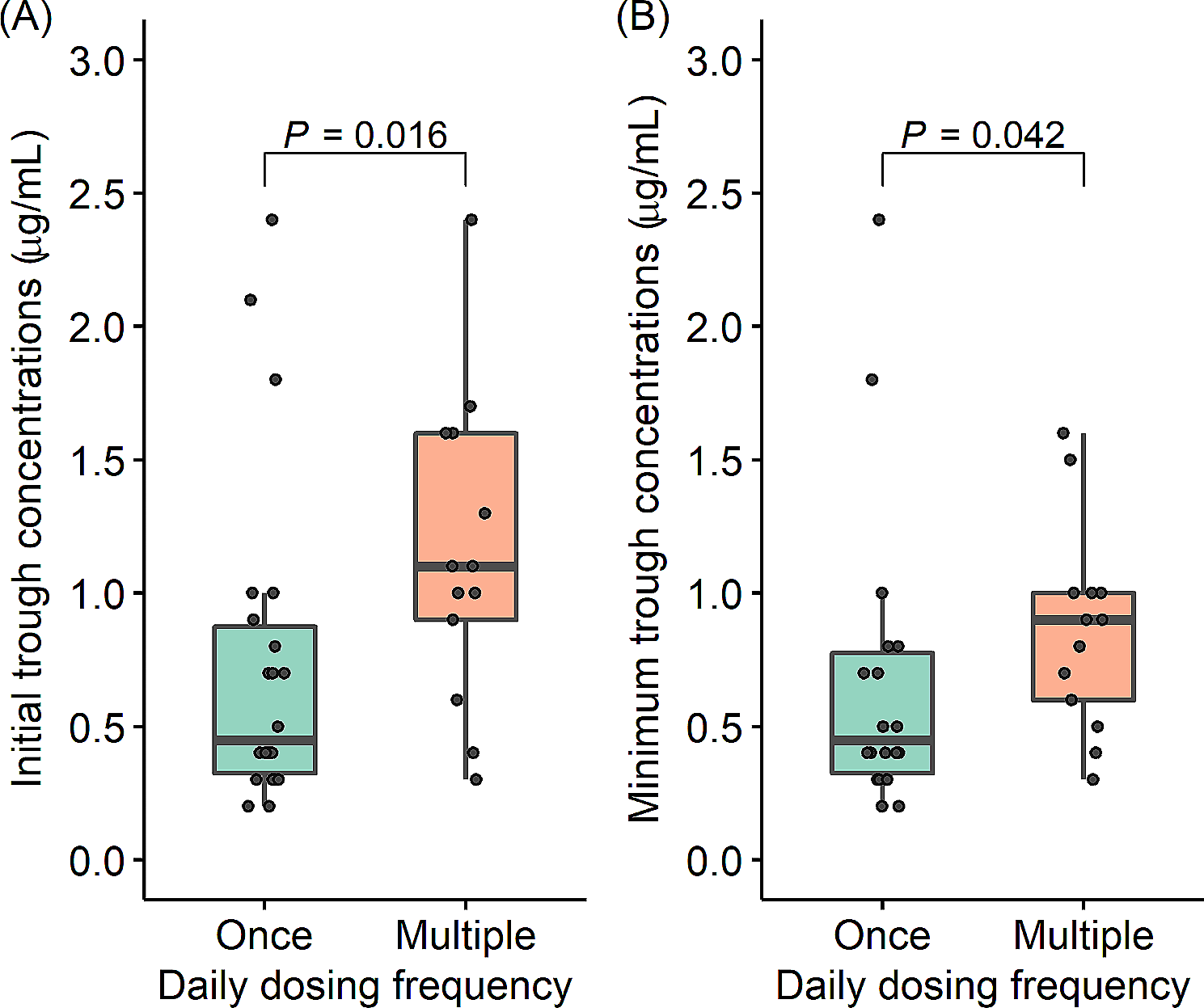
Chemical structures of saturated fatty acids. For example, 14:0 denotes 14-carbon chain with no double bond
Fig. 4
Chemical structures of unsaturated fatty acids. For example, 20:5(n-3) denotes 20-carbon chain with 5 double bonds and the first double bond in the 3rd position from the methyl end. DHA, docosahexaenoic acid; EPA, eicosapentaenoic acid; ETA, eicosatetraenoic acid
In the GPL structures, long-chain fatty acids are esterified to the sn-1 and sn-2 positions of the glycerol backbone. Based on the structures of the polar head groups linked via a phosphodiester bond to the sn-3 position of the glycerol backbone, GPLs are divided into different classes, such as phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylserine (PS), phosphatidic acid (PA), phosphatidylinositol (PI), and phosphatidylglycerol (PG) (Fig. 5). In the structure of PA, only a small phosphate group is attached to the sn-3 position of the glycerol backbone of diglyceride (DG), also called diacylglycerol. The polar head groups of PC, PE, PS, PI, and PG are phosphocholine, phosphoethanolamine, phosphoserine, phosphoinositol, and phosphoglycerol, respectively. PI is further phosphorylated at the 3-, 4-, and 5-positions of the inositol ring to form PI phosphates, namely PI(3)P, PI(4)P, PI(5)P, PI(3,4)P2, PI(3,5)P2, PI(4,5)P2, and PI(3,4,5)P3. In the structure of cardiolipin (CL), two PA molecules are linked to one glycerol molecule. PS, PA, PI, PG, and CL are negatively charged GPLs. Lysoglycerophospholipids, including lysophosphatidylcholine (LPC), lysophosphatidylethanolamine (LPE), lysophosphatidylserine (LPS), lysophosphatidic acid (LPA), lysophosphatidylinositol (LPI), and lysophosphatidylglycerol (LPG), possess only one acyl chain. Plasmanylcholine and plasmanylethanolamine are ether-linked GPLs characterized by an ether bond at the sn-1 position of the glycerol backbone (Fig. 6). Plasmenylcholine and plasmenylethanolamine, also called plasmalogens, contain a vinyl-ether bond at the sn-1 position. Lysoplasmanylcholine and lysoplasmenylcholine have one alkyl chain and one alkenyl chain, respectively.
Fig. 5
Chemical structures of glycerophospholipids and DG. Glycerophospholipids consist of a glycerol backbone, a hydrophilic head group, and two hydrophobic acyl chains. Based on the head group structures, glycerophospholipids are divided into classes, including PC, PE, PS, PA, PI, PG, and CL. Lysoglycerophospholipids, such as LPC, possess only one acyl chain. PI is further phosphorylated to form PI phosphates, including PI(4)P, PI(4,5)P2, and PI(3,4,5)P3. CL, cardiolipin; DG, diglyceride; LPC, lysophosphatidylcholine; PA, phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PG, phosphatidylglycerol; PI, phosphatidylinositol; PS, phosphatidylserine
Fig. 6
Chemical structures of ether-linked glycerophospholipids. Plasmanylcholine and plasmanylethanolamine contain an ether bond at the sn-1 position of the glycerol backbone. Plasmenylcholine and plasmenylethanolamine, also called plasmalogens, contain a vinyl-ether bond at the sn-1 position. Lysoplasmanylcholine and lysoplasmenylcholine possess one alkyl chain and one alkenyl chain, respectively
In the molecular structures of SPLs, such as sphingomyelin (SM), ceramide phosphoethanolamine, and ceramide-1-phosphate, one fatty acid is attached via an amide bond to the sphingosine backbone (Fig. 7). In the structure of SM, the phosphocholine headgroup is bound to ceramide (Cer). Sphingosylphosphocholine and sphingosine-1-phosphate are lysosphingophospholipids. Sphingosine containing 18 carbon atoms (d18:1) is the most common sphingoid base.
Fig. 7
Chemical structures of sphingophospholipids, Cer, and sphingosine. Cer, SM, ceramide phosphoethanolamine, and ceramide-1-phosphate contain one fatty acid attached via an amide bond to the sphingosine backbone. Sphingosylphosphocholine and sphingosine-1-phosphate are lysosphingophospholipids. d18:1 denotes sphingosine containing 18 carbon atoms. Cer, ceramide; SM, sphingomyelin
Thus, a very wide variety of PL molecular species are present in the human body. The biosynthetic pathways of PLs have been summarized in our previous review [30].
Phospholipids and coronary heart diseaseAmong PL classes in plasma lipoprotein particles, PC is the most abundant, and SM is the second most abundant [1, 3]. Other PL classes are quantitatively minor in the lipoprotein particles. The SM/PC ratio in LDL is higher than that in VLDL [1, 31, 32]. Jiang et al. have shown that the levels of plasma SM and the ratios of SM/(PC + SM) are positively associated with coronary heart disease, suggesting that SM is a risk factor [33].
Fernandez et al. have reported that SM (38:2) is associated with an increased risk of cardiovascular disease, whereas LPC (16:0 and 20:4) is associated with a decreased risk of cardiovascular disease [34]. The plasma level of PE (36:5) has been reported to be associated with cardiovascular diseases [35]. There are negative associations of plasma SM (28:1) and LPC (18:1 and 18:2) with incident coronary heart disease events [36]. Sigruener et al. have reported that coronary heart disease mortality is positively associated with the plasma concentrations of PC (30:1, 32:0, 34:1, 36:1, and 38:0), ether-linked PC (PC-O) (32:0, 32:1, 34:0, 34:1, and 38:5), PE (30:1, 32:1, 34:1, 34:2, 34:3, 36:1, 36:2, 36:3, 36:4, 38:2, and 40:1), ether-linked PE (PE-O) (32:0 and 36:2), SM (d18:1/16:0, d18:1/16:1, d18:1/24:1, and d18:1/24:2), and Cer (d18:1/16:0, d18:1/18:0, and d18:1/24:1), but inversely associated with those of PC (32:2, 36:4, 36:5, 38:3, 38:4, 38:5, 38:6, 38:7, 40:6, and 40:7), LPC (16:0 and 18:0), PE (36:6), PE-O (38:7), SM (d18:1/23:0, d18:1/23:1, and d18:1/24:0), and Cer (d18:1/23:0 and d18:1/24:0), suggesting that LPC species and highly polyunsaturated PC species have protective effects [37]. In patients with coronary heart disease, the concentration of Cer (d18:1/16:0) is associated with the occurrence of major adverse cardiac events [38, 39]. Havulinna et al. have reported that serum concentrations of Cer (d18:1/16:0, d18:1/18:0, and d18:1/24:1) are associated with the risk of incident major adverse coronary events in apparently healthy individuals [40]. In addition, Laaksonen et al. have shown that the ratios of Cer (d18:1/16:0)/Cer (d18:1/24:0), Cer (d18:1/18:0)/Cer (d18:1/24:0), and Cer (d18:1/24:1)/Cer (d18:1/24:0) in the plasma are risk predictors of cardiovascular death in patients with stable coronary heart disease and acute coronary syndromes [41]. Alshehry et al. have reported that, in patients with type 2 diabetes mellitus, the plasma levels of Cer (d18:1/24:1), SM (34:1), plasmanylcholine (32:0, 32:1, 34:1, 36:1, and 36:2), plasmenylcholine (32:1 and 34:1), LPC (20:1), and lysoplasmanylcholine (18:0, 18:1, 22:0, 22:1, 24:0, 24:1, and 24:2) were associated with future cardiovascular events, whereas the plasma levels of polyunsaturated acyl chain-containing PC (34:5, 35:4, and 40:6) and plasmenylcholine (36:5 and 38:6) were inversely associated with events [42].
For primary prevention, pravastatin reduces the risk of cardiovascular events. Changes in plasma PI (36:2) and PC (38:4) have been shown to be negatively and positively associated with pravastatin treatment, respectively, and positively and negatively associated with future cardiovascular events, respectively, independent of changes in LDL-cholesterol [43]. In contrast, the plasma levels of PI species are consistently lower in patients with coronary heart disease than in healthy individuals [44]. Idiopathic heart failure is largely attributable to coronary heart disease, pressure overload, and type 2 diabetes mellitus. The plasma concentrations of Cer (d18:1/16:0) and PC (32:0) have been shown to be associated with heart failure risk [45]. Jensen et al. have reported that the plasma concentrations of very-long chain saturated fatty acid (20:0, 22:0, or 24:0)-containing Cer and SM are associated with a reduced risk of atrial fibrillation, whereas those of palmitic acid (16:0)-containing Cer and SM are associated with an increased risk of atrial fibrillation [46]. Long-term high blood pressure is a risk factor for coronary heart disease, and the plasma levels of PC (32:1 and 40:5) and PE (38:3, 38:4, 38:6, 40:4, 40:5, and 40:6) are collectively associated with hypertension [47].
Using lipoprotein model particles, we have demonstrated that SM in the surface monolayer of the particles increases the acyl chain order and reduces the head group hydration, which may be due to the saturated acyl chains and the intra- and intermolecular hydrogen bonding between the amino and hydroxyl groups of SM molecules [48]. We have shown that SM in the particle surface monolayer decreases the binding of apoE to the particles and the apoE-mediated uptake of the particles by HepG2 human hepatoblastoma cells [7]. Additionally, the incorporation of SM into the particle surface potentiates the inhibitory effects of apoC-II and apoC-III on apoE-mediated cellular uptake of the particles [7]. Arimoto et al. have shown that SM in the particle surface monolayer reduces LPL-mediated lipolysis and delays the plasma clearance of the particles in rats [49]. These findings suggest that SM suppresses the removal of TG-rich lipoproteins from the circulation.
In atherosclerotic lesions, many types of cells secrete sphingomyelinase (SMase), which hydrolyzes SM to form Cer and induces the aggregation of LDL particles [50]. Aggregated LDL particles in human atherosclerotic lesions contain higher amounts of Cer than LDL particles in the plasma [
留言 (0)