
記住我







More than one in ten new cases of cancer identified in women are related to breast cancer, making it the most common cancer diagnosed in such a group (Alkabban and Ferguson, 2023). In 2020, approximately 2.3 million women worldwide were diagnosed with breast cancer, resulting in 685,000 deaths related to the disease (Sung et al., 2021). The incidence rate tends to be higher among older women, with a median age of breast cancer diagnosis recorded as 63 years from 2014 to 2018, and increasing to 69 years from 2015 to 2019 (Cao et al., 2021) (Figure 1). Breast cancer is predominantly an incurable metastatic cancer that frequently spreads to distant organs such as the liver, brain, lungs, and bones. Early detection plays a crucial role in achieving a favorable prognosis and higher survival rates (Sun et al., 2017). Global data on cancer incidence rates reveal that breast cancer accounted for 49.3% of all diagnosed cases in women in 2020, surpassing any other cancer type (Lee et al., 2019). Given the existence of multiple biological subtypes, each characterized by unique molecular profiles and clinicopathological characteristics, breast cancer is often regarded as a diverse group of illnesses (>100). Gene expression profiling has allowed the distinction between receptor-positive subtypes (Luminal A, B, Normal like, and HER-2 positive) and receptor-negative subtypes (such as TNBC or Basal-like), in addition to histological subtypes (Kashyap et al., 2022). Breast cancer screening significantly improves survival rates by enhancing the likelihood of early detection and prompt treatment of malignant tumors. The recommended screening modalities for breast cancer include breast self-examination, clinical breast examination, and mammography for women aged forty (Sebastian-delaCruz et al., 2021) and above. While developing nations emphasize the importance of breast self-examination as a pivotal strategy for early breast cancer identification, inadequate funding hinders the implementation of mammography screening (Lee et al., 2019). Presently, the three primary methods used for treating breast cancer are radiation therapy, chemotherapy, and surgery.
In the past, breast cancer was commonly perceived as a malignancy with minimal immunological responses. The immunological landscape surrounding breast cancer comprises a diverse array of cells and cytokines, some with anti-tumor properties while others possess pro-tumor or immunosuppressive characteristics. Persistent inflammation driven by these cells and cytokines can hasten the progression of breast cancer. Among immune system components, such as NK cells and CTLs, there exists a focus on combating cancer by targeting breast cancer cells. However, various elements including tregs, macrophages, MDSCs, and T-helper cells contribute to breast cancer advancement through mechanisms such as promoting metastasis, secreting proinflammatory cytokines, and attenuating the function of cytotoxic T-cells. B-cells may exhibit a protumorigenic role by reducing antitumor immunity, whereas they can also release antibodies that neutralize tumors, thus assuming an antitumorigenic role (Amens et al., 2021a). Inflammation can severely damage breast tissue and facilitate breast cancer development by increasing proinflammatory cytokines like TNF-α and interleukins (Amens et al., 2021a) Tumor cells have shown resilience against chemotherapy through various mechanisms. These mechanisms can be classified into six categories: elevated expression of specific ATP-binding cassette transport proteins, evasion of cell death, enhancement of DNA repair pathways, relevant mutations in cellular targets, increased resilience to adverse environments, and biotransformation of anticancer drugs (Miraghel et al., 2022). When it comes to treatment considerations for breast cancer, several factors are taken into account, including the expression of HER2, progesterone receptor (PR), and estrogen receptor (ER), along with other patient- and tumor-specific characteristics. The protein HER2 is encoded by the erythroblastic oncogene B (ERBB2) gene. Oncogene amplification, leading to the overexpression of HER2, fosters the proliferation of cancer cells and contributes to the emergence of various subtypes of breast cancer (van den Ende et al., 2022).
Reflecting on historical limits, the care of breast cancer has significantly evolved throughout time, leading to a more sophisticated approach. When it comes to early diagnosis, previous screening methods were less advanced and less generally available, resulting in delayed discovery of breast cancer in many cases. However, modern imaging techniques such as digital mammography, ultrasound, MRI, and 3D mammography enable earlier and more accurate identification of breast cancer, hence improving survival rates and treatment outcomes. Aside from that, survival rates for breast cancer were poorer due to fewer treatment options, late-stage diagnosis, and less effective medicines. However, nowadays, the breast cancer survival rates have considerably improved thanks to advances in early identification, tailored treatments, and supportive care, resulting in better long-term results and increasing chances of cure, particularly when the cancer is diagnosed early (Li et al., 2021a). For examples, in recent years, extensive research has focused on various non-invasive biomarkers, including circulating cell-free or exosomal non-coding RNAs, proteins (such as tumor-associated autoantibodies, carcinoembryonic antigen, carbohydrate antigen, tissue polypeptide-specific antigen, etc.), polygenic risk scores involving single nucleotide polymorphisms, and cell-free DNA. Among these biomarkers, circulating cell-free miRNAs have garnered attention. MiRNAs are small non-coding RNAs measuring around 17 to 25 nucleotides, and they play a crucial role in regulating gene expression at a post-transcriptional level. Acting as master regulators, miRNAs can repress translation or degrade messenger RNAs (mRNAs), thus controlling gene expression. They are essential components of biological regulatory processes involved in development, host-pathogen interactions, cell differentiation, proliferation, apoptosis, metabolism, and cancer across various organisms (Maldonado et al., 2022). Mature miRNAs can be exported from cells and are primarily located within the cytoplasm. Some miRNAs persistently exist in bodily fluids like serum, plasma, saliva, or urine due to their interaction with exosomes or RNA-binding proteins, protecting them from degradation. Changes in the levels of these diagnostic circulating miRNAs have been observed even before the detection of tumors using conventional diagnostic tools. Breast cancer research and knowledge have advanced substantially. Breast cancer is now recognized as a diverse disease with distinct subtypes based on molecular features. Aside from that, advancements in treatment procedures increase the identification of specific mutations, gene amplifications, and expression patterns, resulting in personalized therapy regimens tailored to individual patients (O’Day and Lal, 2010). The function of miRNAs in treatment approach has a significant impact on patient survival. The function can affect a variety of biological processes, including cell proliferation, differentiation, apoptosis, metabolism, and immunological response. It regulates several molecular pathways that contribute to the growth of breast cancer. For example, depending on their target genes, miRNAs can serve as oncogenes (oncomiRs) or tumor suppressors. OncomiRs enhance cancer cell proliferation, survival, and invasion by targeting tumor suppressor genes or apoptosis regulators, whereas miRNAs regulate angiogenesis by targeting genes involved in VEGF signaling and angiogenic pathways. They also affect metastasis by altering genes related to cell adhesion, migration, and invasion, encouraging the spread of cancer cells to distant organs. MiRNAs regulate hormone receptor signaling in breast cancer. MiRNAs can target estrogen receptor alpha (ERα) or PR, impacting hormone responsiveness and treatment results in hormone receptor-positive breast tumors. Finally, changes in the expression of various miRNAs are linked to different breast cancer subtypes, clinical stages, and patient outcomes. MiRNAs can be used as diagnostic biomarkers for early detection, as well as prognostic indications for disease progression, metastasis, and treatment response (Loh et al., 2019). Thus, circulating cell-free miRNAs may have greater utility than other listed biomarkers in detecting early-stage breast cancer (Sehovic et al., 2022). The objective of this study is to provide a concise overview of the importance and role of miRNAs in breast cancer. It will discuss their involvement in the antitumor immune response as tumor suppressors and their participation in metastasis as oncogenes. Furthermore, miRNAs hold promise as therapeutic targets for gene therapy due to their ability to influence specific pathways that either promote or inhibit carcinogenesis. This perspective will emphasize the latest strategies developed for therapeutics utilizing miRNA.
Breast cancerBreast cancer, with an estimated 2.3 million new cases worldwide, currently stands as one of the most prevalent malignancies across the globe and ranks as the fifth leading cause of cancer-related deaths. Among women worldwide, breast cancer takes the foremost position in terms of cancer-related mortality (Łukasiewicz et al., 2021a). Breast cancer is a complex disease influenced by various factors. While it affects populations globally, significant regional differences in incidence, mortality, and survival rates exist. These variations can be attributed to factors such as age, estrogen levels, lifestyle choices, and genetics (Momenimovahed and Salehiniya, 2019). Age is a crucial risk factor for breast cancer, and its incidence strongly correlates with advancing age. Being a woman and growing older are the two primary variables affecting the risk, with the majority of cases occurring in women aged 50 or older (Centers for Disease Control and Prevention, 2023). However, extensive research has focused specifically on young women, who often experience more aggressive forms of the disease, receive intensive therapies, and face concerns of lower survival rates (Plichta et al., 2020). The role of estrogen, categorized into endogenous and exogenous estrogens, is significant in breast cancer risk. Endogenous estrogen, particularly in estrogen receptor-positive subtypes that account for 70% of cases after menopause, plays a major role in breast cancer development. Breast adipose tissue in cancer patients exhibits significantly elevated expressions of CYP19 and aromatase, leading to increased estrogen production stimulated by tumor-derived substances such as prostaglandins released by fibroblasts, infiltrating lymphocytes, or epithelial cells (Parida and Sharma, 2019). Additionally, sedentary lifestyle, regular alcohol consumption, and abdominal obesity increase the risk of breast cancer, regardless of menopausal status. In premenopausal women, abdominal obesity is associated with a 3.3-fold increase in risk. Smoking, both current and prior, raises the risk of postmenopausal breast cancer (McCarthy et al., 2021). Finally, genetic influences contribute to breast cancer. Germline mutations in known susceptibility genes account for 5%–10% of cases and are categorized as “high-penetrance,” “moderate-penetrance,” and “low-penetrance” based on their impact and recurrence. Mutations in genes such as BRCA1 and BRCA2 account for 10%–15% of heritable breast cancer cases, while other high-risk genes, including TP53, CDH1, phosphate and tensin homolog (PTEN), and STK11, also lead to breast cancer development. The NF1 gene, associated with neuroectodermal disorders and exhibiting wide variation in gene expression due to autosomal dominant inheritance, is another gene implicated in invasive breast cancer (Naeem et al., 2019).
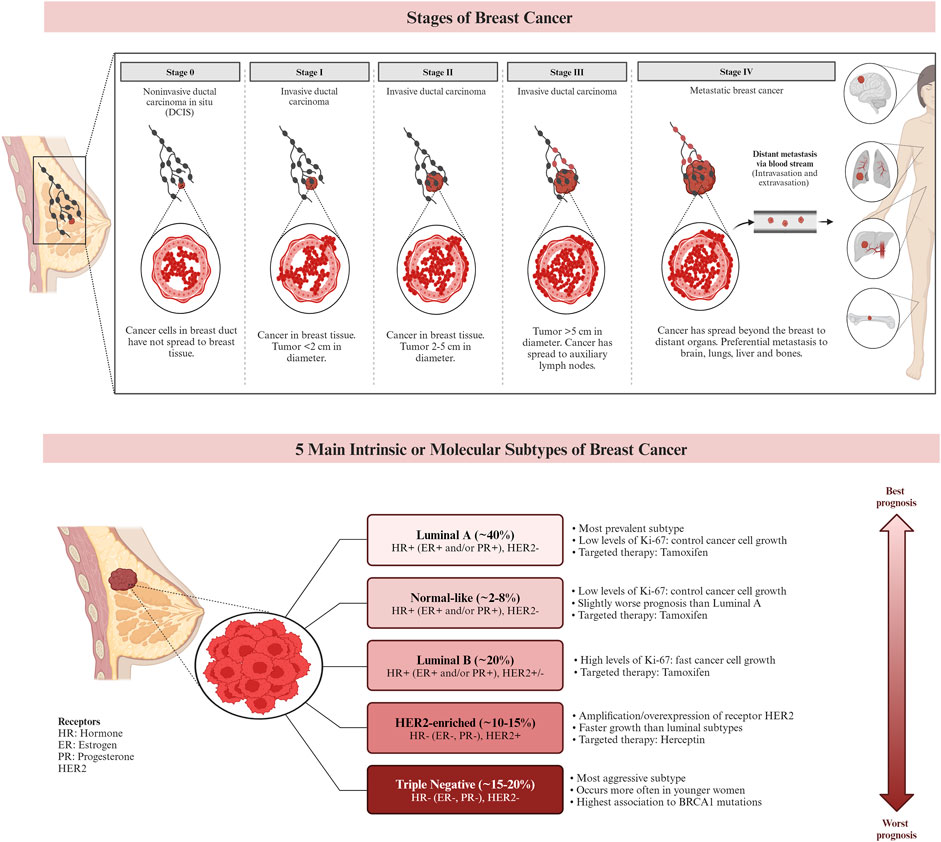
Genetic alterations and DNA damage, which can be influenced by estrogen exposure, are known to contribute to the development of breast cancer. Inherited genes such as BRCA1 and BRCA2 have been associated with an increased risk of cancer. As a result, individuals with a family history of ovarian or breast cancer have a higher likelihood of developing breast cancer themselves. In a healthy individual, the immune system typically targets cells with abnormal DNA or development. However, in cases of breast cancer, this surveillance system may fail, allowing the tumor to grow and spread (Alkabban and Ferguson, 2023). Currently, breast cancer is classified using histopathologic characteristics, molecular characterization, and immunohistochemistry (IHC). Invasive ductal carcinoma and invasive lobular carcinoma are the most common histologic subtypes, accounting for 80%–85%, and 10%–15% of all cases of invasive breast cancer, respectively. Other less common histologic cancer subtypes make up less than 1% of invasive breast cancers. The characterization of breast cancer through IHC plays a crucial role in determining treatment strategies and predicting prognosis (Bhattacharyya et al., 2020). IHC is frequently employed to classify breast cancer subtypes based on the expression of specific markers, including the ER, PR, and HER2. Approximately 73% of breast cancers are ER/PR + HER2− tumors, which represent the most common subtype. It is worth noting that these tumors can also be PR positive. Research suggests that the etiologies of different breast cancer subtypes may vary (McCarthy et al., 2021). Figure 2 demonstrates the stages and subtypes of breast cancer.
Figure 2. The stages and the subtypes of breast cancer. Breast cancer subtypes can be categorized based on their expression of hormone receptors (estrogen receptor (ER) and progesterone receptor (PR)), the proliferation marker Ki-67, and the receptor tyrosine kinase HER2. Targeted therapies, such as Herceptin (aimed at the HER2 protein) and Tamoxifen (aimed at the ER), can be used in the treatment of certain breast cancer subtypes. Prognosis varies based on the subtype of breast cancer. (Larsen et al., 2013; Chen et al., 2018; De Cicco et al., 2019; Hashmi et al., 2019).
Subtypes of breast cancerTriple negative breast cancer (TNBC)Breast cancer subtype known as triple negative breast cancer (TNBC) is a malignant and complex one that presents with challenges for therapeutic targeting due to its absence of expression of the ER, PR, and HER2 receptors. From a biological perspective, TNBC tumors typically exhibit increased aggressiveness and size, along with a higher oncological grade and lymph node metastases. Although metaplastic cancers, which may exhibit squamous or spindle cell differentiation, medullary-like cancers with a prominent lymphocytic infiltrate, and uncommon special type cancers like adenoid cystic carcinoma are the most common histologies observed in TNBC (Cao and Niu, 2020).
Luminal breast cancerER-positive tumors known as luminal breast cancers account for over 70% of all instances of breast cancer. There are two primary biological processes that differentiate Luminal-like cancers into Luminal A and B subtypes: proliferation-related pathways and luminal-regulated pathways. The presence of ER and/or PR and the lack of HER2 are characteristics of luminal A cancers. The ER transcription factors in this subtype activate genes whose expression is typical of the luminal epithelium lining the mammary ducts. Additionally, it exhibits decreased expression of genes involved in cell division. Luminal B tumors, on the other hand, have a worse prognosis and are higher grade. In addition to being ER positive, they could also be PR negative, HER2, or both. Furthermore, genes linked to proliferation are highly expressed in it. This subtype expresses less of the PR and FOXA1 genes and proteins, which are typical of the luminal epithelium, but not the ER (Łukasiewicz et al., 2021b).
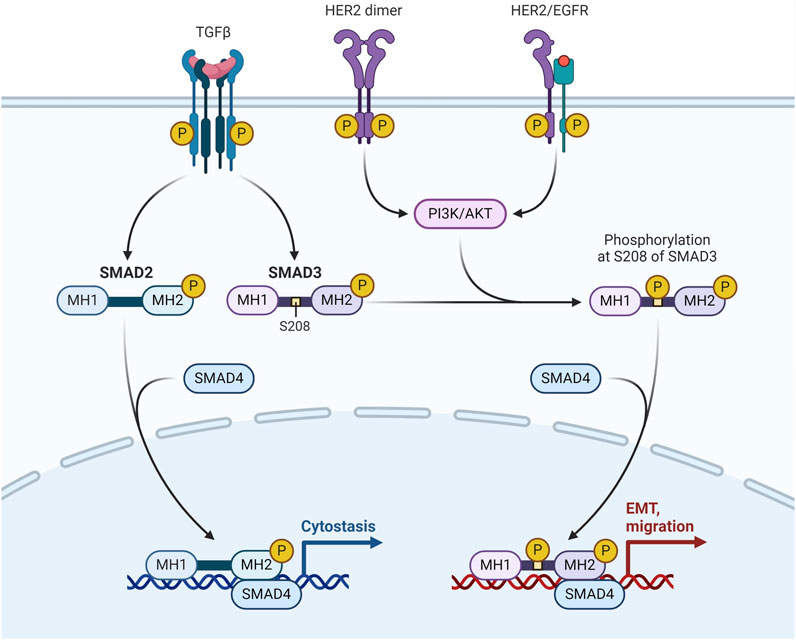
HER2-enriched breast cancerThe transmembrane tyrosine kinase receptor encoded by the HER2, often referred to as HER2/neu and Erb-B2, binds to its extracellular signal to start a cascade that regulates cell proliferation, differentiation, and survival. Adequate tumor development and unfavourable clinical prognosis are caused by overexpression of the HER2 protein and/or HER2 gene amplification in 12%–20% of all breast cancer cases. They have modest levels of ER expression and overexpression of the Erb-B2 oncogene. It's interesting to note that the HER2-enriched subtype does not exhibit upregulation of proliferation genes such Ki-67 and proliferating cell nuclear antigen (Conant and Soo, 2021) (Figure 3).
Figure 3. HER-2/EGFR signaling pathway in breast cancer. HER-2/EGFR signaling switches TGFβ function from inducing antiproliferation to promoting breast cancer development via the AKT-mediated phosphorylation of Smad3 at S208 in the linker (Huang F. et al., 2018).
Claudin-low breast cancerThe most notable gene expression traits that characterize the claudin-low breast cancer subtype include stem cell-like/less differentiated gene expression patterns, high expression of epithelial-mesenchymal transition genes, and low expression of cell-cell adhesion genes. Claudin-low tumors have significant immune and stromal cell infiltration in addition to these gene expression characteristics, but they differ greatly in many other respects. According to reports, most claudin-low breast cancers are triple negative—they do not express the ER, PR, or HER2. These tumors are linked to a poor prognosis (Bergholtz et al., 2020). The poor expression of essential cell-cell adhesion molecules, such as occludin, E-cadherin, and claudins 3, 4, and 7, is a defining characteristic of the claudin-low subtype. Because of their extreme enrichment in stem cell and mesenchymal characteristics, these tumors are regarded as the most primordial forms of breast cancer (Pommier et al., 2020).
Breast cancer patients at stages I, II, III, and IV have 5-year survival rates of 100%, 93%, 72%, and 26%, respectively. While the death rate from breast cancer has decreased as a result of early identification and detection, more advancements in prevention, detection, and treatment are desperately needed to improve the prognosis and survival of breast cancer patients. MiRNAs have garnered significant attention in recent times due to their regulatory role in the development, advancement, and spread of breast cancer. Moreover, there exists a strong correlation between the expression level of specific miRNAs and the morphological characteristics, immunohistochemical profiles, histopathological parameters, clinical outcomes, prognosis, and response to treatment of breast cancer It is noteworthy that approximately half of the human miRNA-encoding genes are located in regions associated with cancer or fragile chromosomal sites. Various studies have demonstrated altered expression of miRNAs in breast cancer since the initial discovery of miRNA dysregulation in breast cancer in 2005 (Loh et al., 2019).
MiRNAs and the mechanismMiRNAs have become a focal point in both fundamental and applied biomedical research due to their impact on gene expression, widespread presence in bodily tissues and fluids, and potential as disease biomarkers (Citron et al., 2017; Yoon et al., 2020). The primary mechanism by which miRNAs exert their influence is through the targeting of messenger RNA (mRNA) (Pu et al., 2019). Alterations in miRNA expression can significantly impact target regulation, thereby influencing cellular homeostasis. Consequently, the relative levels of miRNA, and subsequently mRNA, play a pivotal role in processes such as carcinogenesis and other diseases (Hill and Tran, 2021).
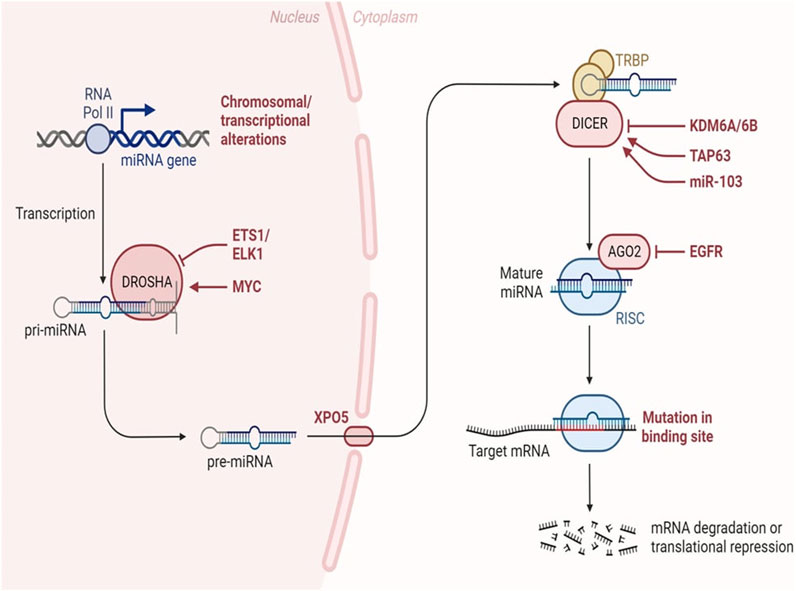
Mechanism involved in MiRNAs and its Pathological functionsBiogenesisMiRNA biogenesis denotes the intricate process by which these diminutive RNA molecules are synthesized within a cell. This intricate pathway involves several sequential stages, commencing with the transcription of miRNA genes and culminating in the production of mature miRNAs capable of finely modulating gene expression. The maturation of miRNAs is facilitated by the activity of RNAse III enzymes, namely, Drosha and Dicer in animals, and DCL in plants. Following cropping and exportation to the cytoplasm, miRNAs undergo further processing, being diced into smaller fragments, which are subsequently incorporated into Argonaute proteins. Notably, Drosha, in conjunction with Di George syndrome critical region gene 8 (DGCR8), assumes the role of a molecular ruler, precisely measuring the cleavage site during this intricate miRNA biogenesis process (Hajarnis et al., 2015). The biogenesis of miRNAs initiates with the transcription of dedicated information units by RNA polymerase II, progressing through successive maturation steps to produce the final functional regulators (Treiber et al., 2019). Two distinct groups of enzymes participate in miRNA biogenesis: processors, comprising RNA endonucleases cleaving miRNA precursors in the nucleus (Drosha) and the cytoplasm (Dicer), and effectors, encompassing the Argonaute family (Ago protein) and proteins orchestrating post-transcriptional regulatory effects on mRNA targets (RISC complex) (Daugaard and Hansen, 2017). The overall pathway includes the transport system responsible for exporting miRNA precursors from the nucleus to the cytoplasm (Exportin-5) and auxiliary proteins modulating specific steps, such as TRBP and DGCR8. Nuclear processors are exclusive to miRNA biosynthesis, while the cytoplasmic branch of the pathway may intersect with other regulatory mechanisms like RNA-mediated interference (RNAi). Human miRNA biogenesis involves a two-step process with nuclear and cytoplasmic cleavage events. In the nucleus, miRNAs are transcribed as long transcripts termed pri-miRNA, either by their independent promoters or by sharing promoters with their host gene (Creugny et al., 2018). This transcription is predominantly executed by RNA polymerase II (Schier and Taatjes, 2020), although certain virus-encoded miRNAs are transcribed by RNA polymerase III (Nanbo et al., 2021). Recognition of a stem-loop structure within the pri-miRNA occurs through the action of the RNase III Drosha and its cofactor DGCR8 (Nguyen et al., 2020). This intricate process ensures the formation of mature miRNAs essential for post-transcriptional gene regulation.
The miRNA biogenesis pathway is depicted schematically, encompassing the key enzymes and cellular compartments involved. Eukaryotic genomes harbour transcriptional units responsible for generating miRNAs, which can be found in various genomic regions, including intergenic spaces, coding genes, and non-coding genes. Following transcription, primary miRNAs adopt a characteristic hairpin-loop secondary structure, recognized and excised by the microprocessor complex comprising DGCR8 and Drosha. Subsequently, the resulting precursor miRNAs undergo exportation to the cytoplasm, where the Dicer nuclease processes them into double-stranded RNA. The mature miRNA chain is then selected by Ago2 and incorporated into the RNA-induced silencing complex, initiating its regulatory function. In this intricate process, transcriptional units within the genome give rise to miRNAs, which exhibit a diverse genomic distribution. The microprocessor complex, involving DGCR8 and Drosha, plays a crucial role in recognizing and cleaving the hairpin-loop structure of primary miRNAs. Following this nuclear processing, precursor miRNAs are transported to the cytoplasm, where Dicer further refines them into double-stranded RNA. The mature miRNA is subsequently chosen by Ago2 and integrated into the RNA-induced silencing complex, where it becomes a pivotal player in post-transcriptional gene regulation (Figure 4).
Figure 4. MiRNA biogenesis in cancer. Changes occurring throughout the process of miRNA synthesis can impact the accessibility of the target mRNA. Adapted from Lin et al. (2015) (Lin and Gregory, 2015).
Regulation of gene expressionMiRNAs orchestrate post-transcriptional gene regulation by binding to target messenger RNAs (mRNAs), inducing either translational repression or mRNA degradation (Sebastian-delaCruz et al., 2021; Rani and Sengar, 2022). This regulatory mechanism is pivotal for precise modulation of gene expression, contributing to the maintenance of cellular homeostasis (Fernández-Hernando and Suárez, 2018).
MiRNAs regulate gene expression post-transcriptionally in a sequence-specific manner, with significant biological impact (Acuña et al., 2020). Research has delved into understanding the mechanisms of miRNA-mediated post-transcriptional regulation, using in vitro, in vivo, and cell-free extracts, along with bioinformatics tools predicting miRNA regulation of nearly 30% of protein-coding genes in mammalian cells (Boivin et al., 2018). MiRNAs influence translation by inhibiting initiation, preventing ribosomal association, and blocking elongation (Oliveto et al., 2017; Ahmed et al., 2018; Naeli et al., 2023). They also regulate mRNA stability by recruiting decay machinery components, leading to deadenylation or decapping followed by exonuclease activity (Frédérick and Simard, 2022; Passmore and Coller, 2022). Additionally, miRNAs are secreted as exosomes or into extracellular fluids, suggesting potential roles in intercellular communication and as disease biomarkers, including infectious diseases (Acuña et al., 2020). Figure 4 shows the how the alterations throughout miRNA biogenesis can affect availability of target mRNA.
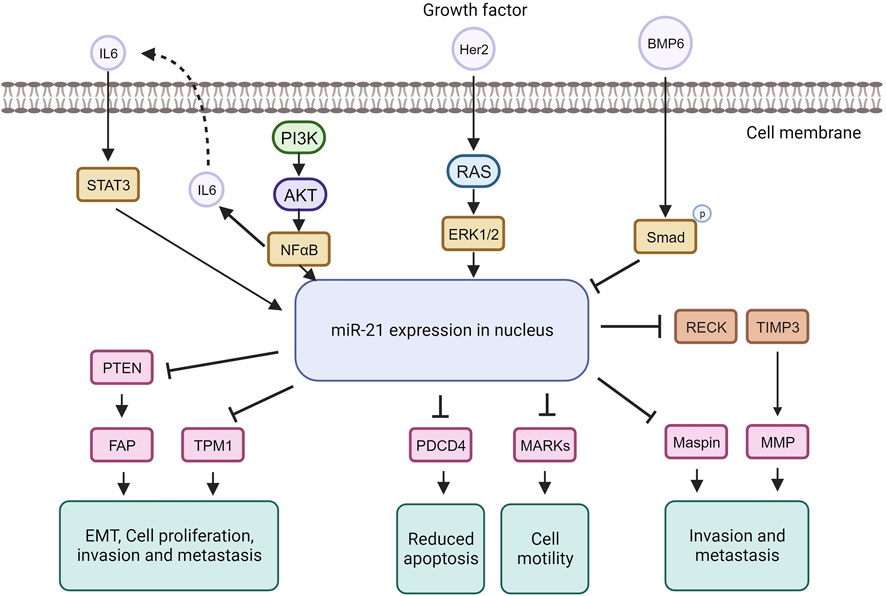
Disease associations in cancer (OncomiRs)MiRNA dysregulation is commonly linked to cancer, where some function as oncogenes (oncomiRs), promoting tumor formation, while others act as tumor suppressors (O’Bryan et al., 2017). In breast cancer, miR-21, an extensively studied oncomiR, is often elevated, leading to increased cell proliferation and invasion. MiR-21 exerts its oncogenic functions by targeting genes like TPM1, PDCD4, PTEN, and Smad7, enhancing tumor cell growth and inhibiting apoptosis (Zhang Y. et al., 2020; Najjary et al., 2020; Shen et al., 2020) (Figure 5).
Figure 5. MiRNA-21 targets and their function in tumorigenic pathways.
The tumor suppressor TPM1 is suppressed by miR-21, promoting cancer cell growth (Shen et al., 2020). TPM1, part of the actin-binding protein family, is crucial for regulating contraction by mediating calcium response in muscle. It acts as a tumor suppressor in neoplastic transformation, inhibiting cell growth and transformation (Sun et al., 2020). Research suggests TPM1 stabilizes the cytoskeleton, hindering actin filament movement, cell proliferation, and migration (Hardeman et al., 2020). Increased miR-21 under high glucose conditions downregulates TPM1, promoting cell proliferation and migration. Treatment with Tan counteracts this effect by upregulating TPM1, inhibiting proliferation and migration. Inhibiting TPM1 function partially reverses Tan’s anti-proliferative and anti-migrative effects (Jia et al., 2019).
MiR-21 represses the tumor suppressor PDCD4, impacting the mTOR pathway involved in cell growth and proliferation (Cristofoletti et al., 2019; Moustafa-Kamal et al., 2020). MiR-21 downregulates PDCD4 expression by directly targeting its 3′UTR region. PDCD4, a pro-inflammatory tumor suppressor protein, is consistently reduced in human cancers. Under normal conditions, PDCD4 is primarily in the nucleus but can move to the cytoplasm in response to environmental changes. PDCD4 regulates tumor cell apoptosis by binding to eukaryotic initiation factors 4A and 4E (Takaki and Eto, 2018; Wang and Yang, 2018; Matsuhashi et al., 2019). In breast cancer, miR-21 downregulates both PTEN and Smad7. Through PTEN inhibition, miR-21 induces fibroblast activation protein, promoting tumor cell growth. In contrast, Smad7 inhibition by miR-21 enhances fibroblast-to-myofibroblast transition and alpha-smooth muscle actin expression, boosting proliferation and invasion (Najjary et al., 2020). MiR-21 plays a role in the epithelial-mesenchymal transition of breast cancer cells by affecting the p-AKT and p-ERK pathways through PTEN inhibition (Najjary et al., 2020). The upregulation of Her2/neu receptors activates the MAPK signaling pathway, leading to increased miR-21 expression, promoting invasion and metastasis (Sareyeldin et al., 2019). Studies indicate that elevated miR-21 is associated with advanced breast cancer stages, metastasis, and poor prognosis (Anwar et al., 2019). MiR-21 influences invasion and metastasis through its targets PTEN, TPM1, PDCD4, and the tumor suppressor maspin (Huang et al., 2020), making it a potential target for cancer therapy. However, anti-cancer drugs like doxorubicin can paradoxically upregulate miR-21 through NF-kappa B activation, promoting breast cancer progression (O’Bryan et al., 2017). Nevertheless, targeting miR-21 in cancer therapy has shown increased sensitivity to anti-cancer agents in breast cancer (Hashemi et al., 2023).
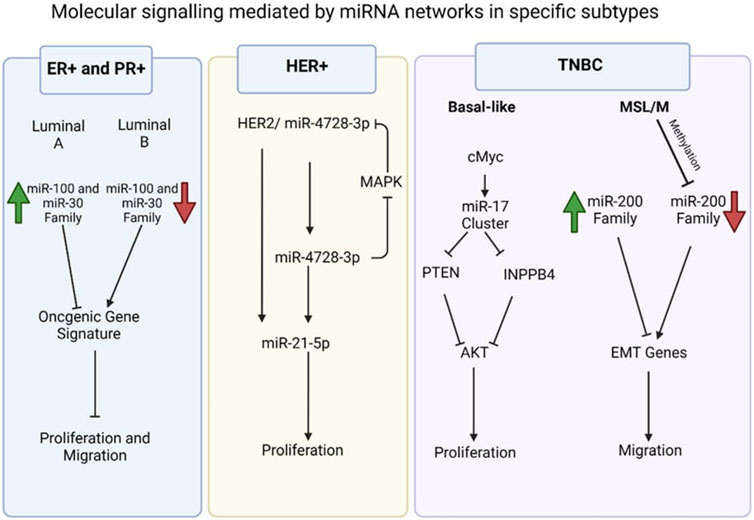
Dysregulated MiRNA profiles in breast cancerBreast cancer subtypes defined by gene and miRNA expressionExamples of significant miRNAs associated with specific breast cancer subtypes and their effects on cell phenotypes are shown in Figure 6; Table 1 demonstrates the list of miRNAs involvement in regulation of breast cancer. In estrogen receptor positive/progesterone receptor positive (ER+/PR+) breast cancers, significant miRNAs such as miR-100 and the miR-30 family play essential roles, particularly in distinguishing between luminal A and luminal B molecular subtypes. In human epidermal growth factor receptor two positive (HER2+) breast cancers, miR-4728-3p, located within the intronic region of HER2, is co-expressed with HER2 and involved in feedback regulation of HER2 and oncogenic miR-21-5p, thereby promoting several oncogenic processes in advanced tumors. In TNBCs, the cMYC oncogene-driven miR-17–92 cluster is overexpressed, especially in the basal-like one molecular subtype, promoting proliferation by targeting inhibitors of the proliferation mediator AKT such as PTEN and inositol polyphosphate-4-phosphatase type II B. Furthermore, in TNBC cell lines categorized as mesenchymal stem-like and mesenchymal molecular subtypes, epigenetic changes inhibit the miR-200 family, facilitating the expression of epithelial-to-mesenchymal transition and migration genes, ultimately promoting a migratory phenotype (Arun et al., 2022).
Figure 6. Molecular signalling mediated by miRNA network in specific subtypes (Arun et al., 2022).
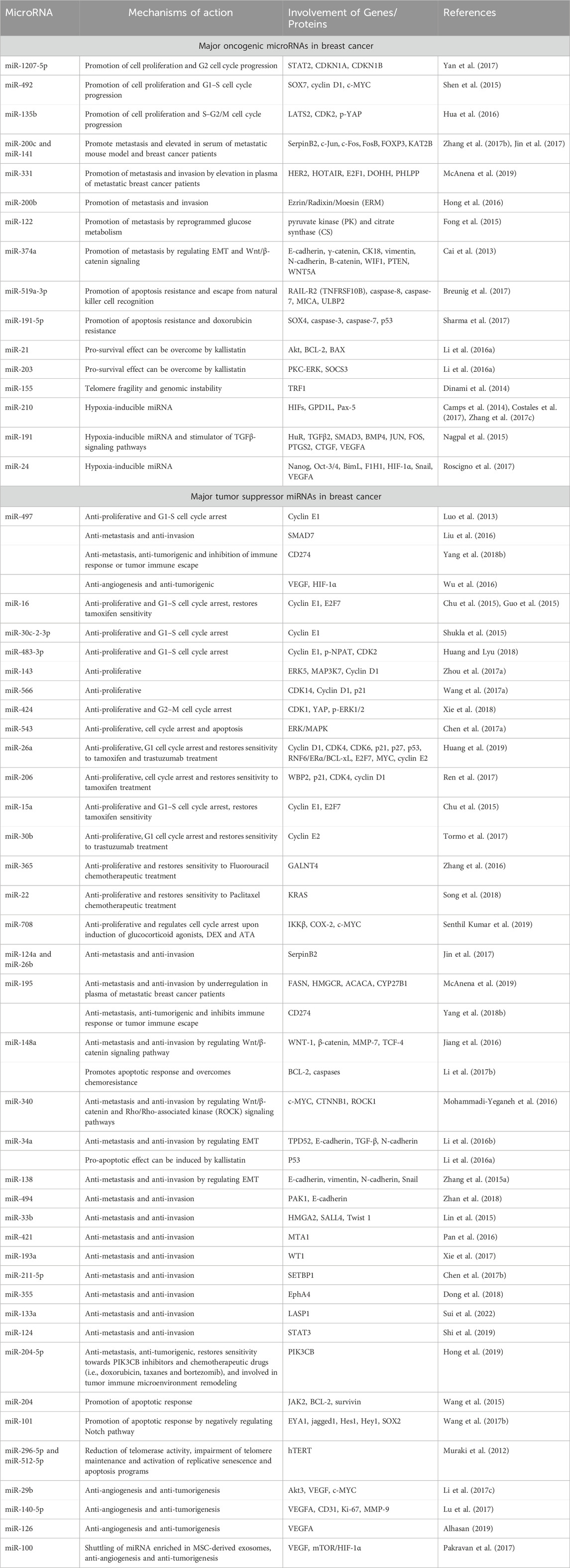
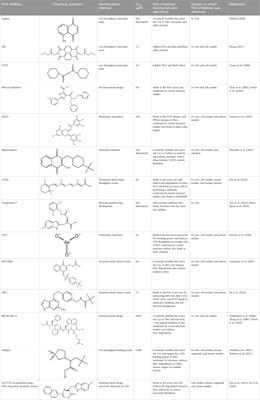
Table 1. Involvement of miRNAs in the regulation of breast cancer.
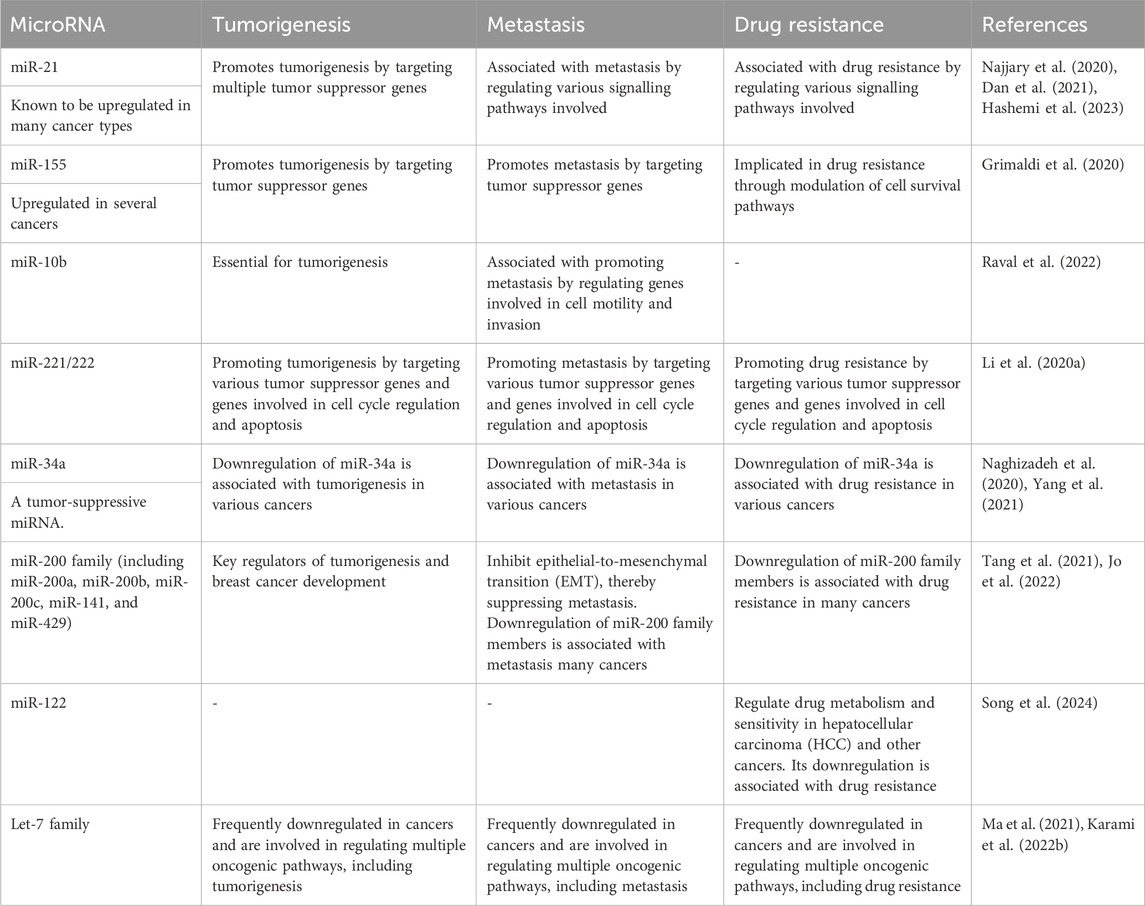
Specific MiRNAs linked to tumorigenesis, metastasis, and drug resistanceSeveral miRNAs have been identified as key players in tumorigenesis, metastasis, and drug resistance across various cancer types (Pan et al., 2021). These miRNAs play crucial roles in cancer progression and treatment response, making them potential targets for therapeutic interventions and biomarkers for prognosis and drug response prediction as shown in Table 2. However, their precise roles can vary depending on the specific cancer type and context, highlighting the complexity of miRNA-mediated regulation in cancer biology (Hill and Tran, 2021).
Table 2. MicroRNAs and their relationships with tumorigenesis, metastasis, and drug resistance.
MiRNA as diagnostic biomarkersThe care of patients in breast cancer remains a significant clinical problem. The scientific community is currently engaged in the ongoing exploration of supplementary biomarkers that can be employed in clinical settings to enhance the efficacy of oncologists in patient management (Jordan-Alejandre et al., 2023). Scientists have utilized genomics, bioinformatics, and molecular biology methodologies to identify miRNAs that offer significant diagnostic insights (Wu et al., 2014). As an illustration, Li et al. conducted a study wherein they employed the exiqon miRNA qPCR panel to screen plasma samples of breast cancer patients (as opposed to normal controls) and identified five distinct types of miRNAs (Let-7b-5p, miR-122-5p, miR-146b-5p, miR-210-3p, and miR-215-5p). Subsequently, they proceeded to validate their findings through quantitative reverse transcription polymerase chain reaction (qRT-PCR) and found that the ROC correlation coefficient for miRNA expression in both samples was 0.978. This finding provides evidence for the potential of the miRNA panel as a biomarker in the diagnosis of breast cancer (Li M. et al., 2019). In addition, Bakr et al. outlined the utilization of miR-373 as a diagnostic biomarker in individuals with breast cancer. Furthermore, they conducted an assessment of the target genes (VEGF and cyclin D1) of miR-373, revealing its role as an oncomiR. This finding suggests that miR-373 could serve as a crucial biomarker for the diagnosis and prognosis of breast cancer through its targeting of VEGF and cyclin D1 (Bakr et al., 2021).
In contrast, Zhang et al. conducted a study investigating the increase of miR-26b-5p, miR-106b-5p, miR-142-3p, miR-142-5p, miR-185-5p, and miR-362-5p in patients with breast cancer. The analysis demonstrated that the miRNA panel had the ability to distinguish between breast cancer patients and healthy controls, and it also exerted an influence on multiple cancer-related pathways (Zhang K. et al., 2021).
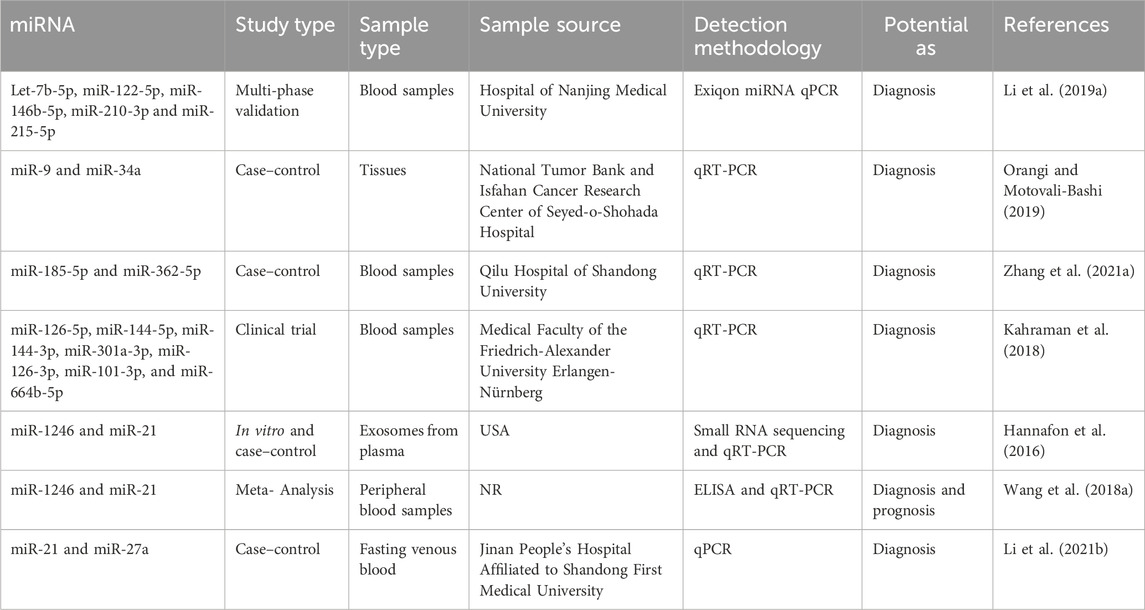
In addition, Lv et al. established a correlation between the decrease in miR-145 levels and the diagnosis of breast cancer, regardless of the specific kind of cells involved. A meta-analysis was conducted on a total of eight papers that employed quantitative methodologies utilizing qRT-PCR. They demonstrated that the expression of miR-145 was 2.57 times higher in breast cancer tissues compared to normal breast tissues. The research findings indicated a significant inverse correlation between the expression of miR-145 and the histological grade of tumors, so implying the potential utility of miR-145 as a biomarker (Lv et al., 2020). The diagnostic relevance of seven miRNA molecules (miR-126-5p, miR-144-5p, miR-144-3p, miR-301a-3p, miR-126-3p, miR-101-3p, and miR-664b-5p) was investigated and emphasized by Kahraman et al. The researchers employed RT-qPCR to examine 63 blood samples obtained from women, consisting of 21 patients diagnosed with TNBC and 21 healthy patients. The study revealed that miR-126-5p exhibited a substantial expression level, making it a prominent miRNA that suppresses tumor growth in breast cancer. Furthermore, it was found to be the most influential miRNA for diagnosing breast cancer in the panel under analysis (Kahraman et al., 2018). Table 3 presents a diverse range of miRNA compounds that have the potential to serve as biomarkers.
Table 3. MiRNA biomarkers as diagnosis and prognosis.
MiRNA and cancer immune evasionMiRNA regulated innate immune response through immune cellsImmune cells in the body provide innate immune response whenever tumor cells arise which should kill and eliminate the tumor cells. However, these tumor cells are able to escape from the immune response and progress to another area of the body, aggravating the cancer (Thomopoulou et al., 2021).
This is made possible by the bi-directional communication between tumor cells and immune cells. These interactions create tumor microenvironment (TME) where tumor cells use different mechanisms to manipulate the cells around it (Tan and Naylor, 2022). Tumor cells manipulate other cells to meet their metabolic needs. Since tumor cell metabolism differs from normal cell metabolism, it competes with normal cells for resources like glucose. In an article by Elia and Haigis (2021), it is stated that T cells and cancer cells compete for glucose and amino acids. Due to T cell dependency on metabolism, limited glucose supply alters its function and differentiation which helps in tumor progression. Similarly, other immune cells also compete with cancer cells to sustain their metabolism which affects the normal function of the immune cells thus allowing the tumor to progress and metastasize (Elia and Haigis, 2021).
The impaired functionality of immune cells can either be corrected or further impaired by miRNAs. MiRNAs play a crucial role in cancer immune evasion by regulating the pathways of immune cells involved in cancer like macrophages, NK cells, Tregg cells, and T helper cells (Cortez et al., 2019). Unfortunately, while some miRNAs can manipulate the impaired immune cells to stop tumor progression, others influence tumor cells activities leading to cancer progression.
MiRNAs can be used as biomarkers for various cancers and infectious diseases. In breast cancer patients, miRNA-125b (miR-125b) and miRNA-155 (mir-155) are used for diagnosis, monitoring prognosis, and assessing patient’s response to chemotherapy. In addition to that, they are also believed to have the ability to regulate innate immune response in breast cancer by modulating the M1/M2 profile.
The most discussed miRNA in relation to breast cancer is miR-155. In breast cancer, overexpression of miR-155 was initially thought to be oncogenic however, a study conducted by Wang et al. (2022), proves otherwise when it demonstrates that miR-155 deficient macrophages increases tumor progression while macrophages showing an overexpression of these noncoding RNAs improves tumor suppression (Wang et al., 2022a). One of the main functions of miR-155 is regulating innate immune responses by differentiating B lymphocytes and CD4+ T cells as well as activating Tregg cells. Furthermore, miR-155 has a direct relation with dendritic cells in which during the immune response in the early stage of breast cancer, both activation of dendritic cells and expression of miR-155 are increased. A study by Asadirad et al. (2019), stated that miR-155 is transported into dendritic cells via the exosomes derived by the cancer cells to increase the expression of immune cells like MHCII, CD86, CD40, CD83, IL12p70, IFN-γ, and IL10. In addition to that, the abundance of miR-155 seen in early breast cancer patients as compared to metastatic breast cancer patients indicates that miR-155 is responsible for the regulation of innate immune response in breast cancer (Asadirad et al., 2019).
Other miRNAs that are highly expressed in early breast cancer include miRNA-126 (miR-126), miRNA-19a (miR-19a), and miRNA-20 (miR-20). Due to that, these three miRNAs are also believed to have an influence in the regulation of innate immune response in breast cancer. MiR-126 prevents breast cancer metastasis by reshaping the TME. Reshaping of TME is made possible by targeting the stromal cell-derived factor 1 (SDF-1) which suppresses the production of monocytes. MiR-19a prevents breast cancer metastasis by converting tumor-associated macrophages (TAM) or M2 macrophages to M1 macrophages through the inhibition of various proto-oncogenes. MiR-20 is a T-helper cells modulator which affects breast cancer by targeting FOXO-1 and STAT3 pathway (Ilhan et al., 2023).
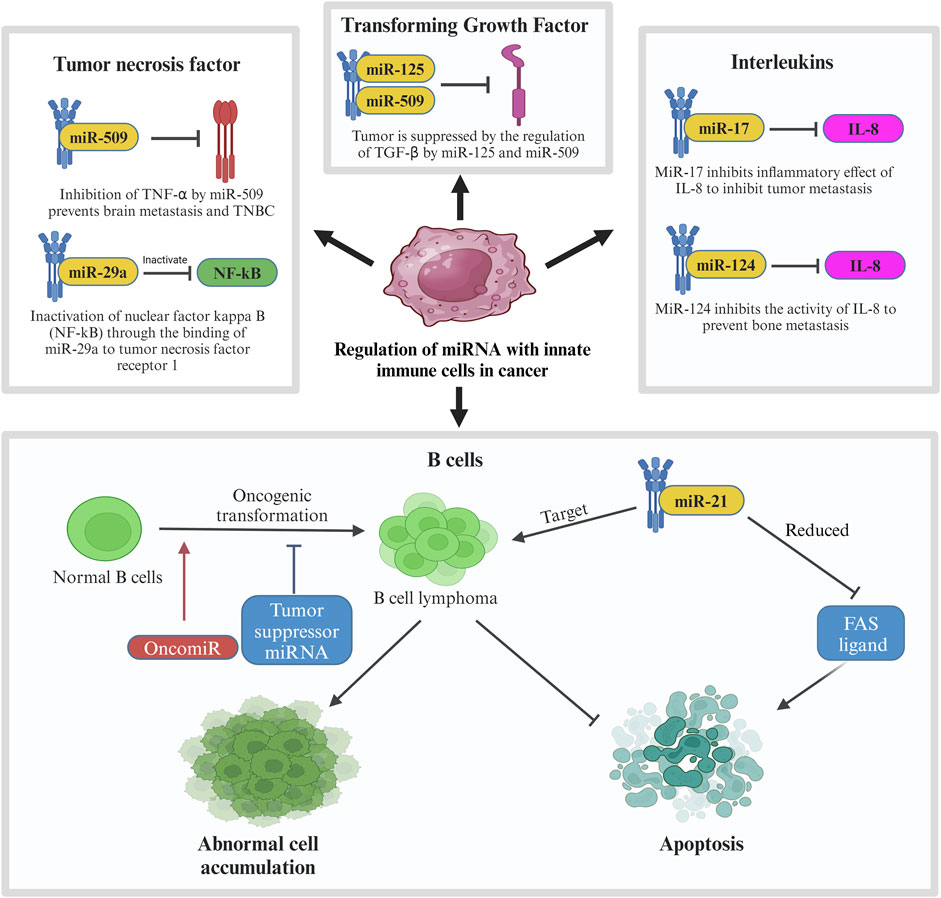
Like miR-20, many other miRNAs affect the pathway of cytokines produced by the innate immune system as seen in Figure 7. When the body detects breast cancer cells, innate immune cells like lymphocytes and macrophages secrete cytokines like tumor necrosis factor (TNF), transforming growth factor (TGF), and interleukins (IL). MiRNAs affect these cytokines to either promote or suppress tumor progression (Bahiraee et al., 2019). One of the cytokines, tumor necrosis factor-alpha (TNF-α), is involved in brain metastasis of primary breast cancer. MiRNA-509 (miR-509) prevents this action by suppressing the levels of TNF-α regulated in the tumor environment. Suppression of TNF-α by miR-509 also prevents TNBC cell proliferation and invasion. Another miRNA involved with TNF is miRNA-29a (miR-29a). This miRNA binds to 3′-UTR of tumor necrosis factor receptor 1 (TNFR1) gene to inactivate the nuclear factor kappa B (NFκB) pathway. Inactivation of this pathway reduces the cell proliferation and induces apoptosis to MCF-7 cell which is a cell from the breast cancer cell line (Zhao et al., 2017).
Figure 7. Regulation of miRNA with innate immune cells in cancer. In breast cancer, innate immune cells including tumor necrosis factor, transforming growth factor, interleukins and B cells are involved in regulation of miRNA in which their pathways are interrupted by tumor suppressor miRNAs to prevent further tumor metastasis. In contrast, oncogenic MiR-21 is involved in the mechanism by targeting B cell lymphoma, leading to abnormal cell accumulation and suppression of apoptosis.
Another type of cytokine, transforming growth factor beta (TGF-β) plays a very important role in cell differentiation, proliferation, and motility in breast cancer which also makes it crucial for TGF-β to be controlled by a regulatory network (Tzavlaki and Moustakas, 2020). In breast cancer, the regulation of TGF-β is suppressed which leads to its overactivation. Overactivation of TGF-β promotes invasion and metastasis of breast cancer cells. In relation to TGF-β in breast cancer, some miRNAs like miR-10b, miR-21, miR-106b, and miR-181 are regulated by TGF-β to promote metastasis while others like miR-145 and miR-206 regulate TGF-β to suppress tumor progression. Other than TGFs, innate immune cells also produce various interleukins. IL-1, IL-8, and IL-11 are known to aid in tumor progression in breast cancer. IN TME, these ILs promote invasion and metastasis of the cancer cells. Certain miRNAs like miR-17, miR-20, and miR-124 supress the effect of these ILs to inhibit tumor progression. MiR-17 and miR-20 bind to 3′-UTR of IL-8 to inhibit the in the inhibition of tumor metastasis to the neighbouring cells whereas the expression of miR-124 prevents bone metastasis caused by IL-11. It is observed that miR-124 has a direct downstream effect to IL-11 by reducing the luciferase activity of the 3′-UTR of IL-11. Overexpression of miR-124 inhibits the activity of IL-11 which prevents bone metastasis in breast cancer patients.
Another interleukin targeted by miRNAs in breast cancer is IL-6. Targeting this interleukin has a dual effect in breast cancer which may either be tumorigenic or anti-tumorigenic. The tumorigenic effect is caused by inhibiting apoptosis, triggering the survival of the tumor cells, and allowing metastasis. IL-6 stimulates miR-155 expression which is known to target suppressors of cytokine signalling pathway (SOCS), hence promoting the progression of breast cancer. Blocking IL-6 pathway can prevent this progression through the inhibition of tumor migration and invasion (Gyamfi et al., 2018). Another method of overcoming this situation is through the downregulation of miR-155 using a photosensitizer. The anti-tumorigenic effects are caused by inhibiting IL-6/STAT3 and SOCS pathway. One of the characteristics of miR-146 is suppression of inflammatory cytokines expression (Gao and Dong, 2017) which makes it a good tumor suppressor marker. In primary breast cancer, miR-146 expresses a negative feedback loop to IL-6 as a mean to inhibit the tumor progression initiated by IL-6/STAT3 pathway. Other miRNAs that can block IL-6 pathway are miR-203 and miR-7. MiR-203 directly inhibits the SOCS pathway in MCF7 cells which suppresses breast cancer progression while miR-7 directly binds to 3′-UTR of IL-6 to negatively regulate its expression which results in antitumorigenic and antimetastatic effects in breast cancer. Figure 7 shows the regulation of miRNA with innate immune cells in breast cancer.
Although the immune system has similar mechanism in every individual, in breast cancer patients, the expression of miRNAs may differ in different patients based on their race. This is because they are genetically influenced and causing women of different races to have different miRNAs expression. A study on Latina breast cancer patients shows that upregulation of miR-141-3-p is associated with cancer recurrence, while downregulation of miR-166 and miR-150-5p are associated with large tumors (Almohaywi et al., 2023). Another study conducted on Egyptian breast cancer patients shows an overexpression of miR-29b and under expression of miR-31 that are linked to the patient’s relapse (Abbas et al., 2022). Therefore, patient ancestry is an important factor when considering the breast cancer biomarkers.
MiRNA regulates specific immune responses in breast cancerThe specific immune system is also known as adaptive immune response which produces respective antibodies that can be used to fight against the particular microbes. The certain microbes that have previously entered the body will be immediately attacked by the immune system if they again infect the specific individual (NCHBI, 2020). The common examples of specific immune responses in breast cancers are the cytotoxic T lymphocytes, natural killer cells and B cells. The cytotoxic T lymphocytes and natural killer cells exert their effect in a direct route onto the breast cancer cells by killing the abnormal cells therefore to induce the antitumor immunity. They play the dominant character in fighting against breast cancer. As breast cancer is growing, the quantity of T-cells will be increasing simultaneously but often associated with reduction in its function due to the extremely immuno-suppressive surroundings in malignant conditions (Speiser and Verdeil, 2017). On the other hand, B cells will result in antitumor immunity through the secretion of immunoglobulins which are able to minimize the amount of early neoplasms (Amens et al., 2021b).
In this case, the tumor cells will have two-way interaction with their local environment which known as tumor microenvironment with the presence of tumor, normal breast, immune, endothelial cells, fibroblast that found in the existence of cancer and mesenchymal stromal (Syeda et al., 2022). By simply secreting the small extracellular vesicles, bidirectionally, tumor cells able to signal the other components. These generated small extracellular vesicles may modulate the activity of mesenchymal stromal, cancer-associated fibroblasts and immune. Meanwhile particularly during hypoxic condition, the performance of endothelial cells can be controlled by mesenchymal stromal and tumor (Rashid et al., 2023). The combination of immune, cancer-associated fibroblast and endothelial cells may enhance angiogenesis (Fu et al., 2020). The major role will be presented by the breast cells which able to largely control the antitumor immune response by producing the small extracellular vesicles and certain extracellular signaling molecules (Mammes et al., 2021). It therefore led to destruction in the immune cells as the products consist of numerous T cells that responsible to impede or activate the polarization, activation, expansion, movement and recruitment of the target cells (Loric et al., 2023).
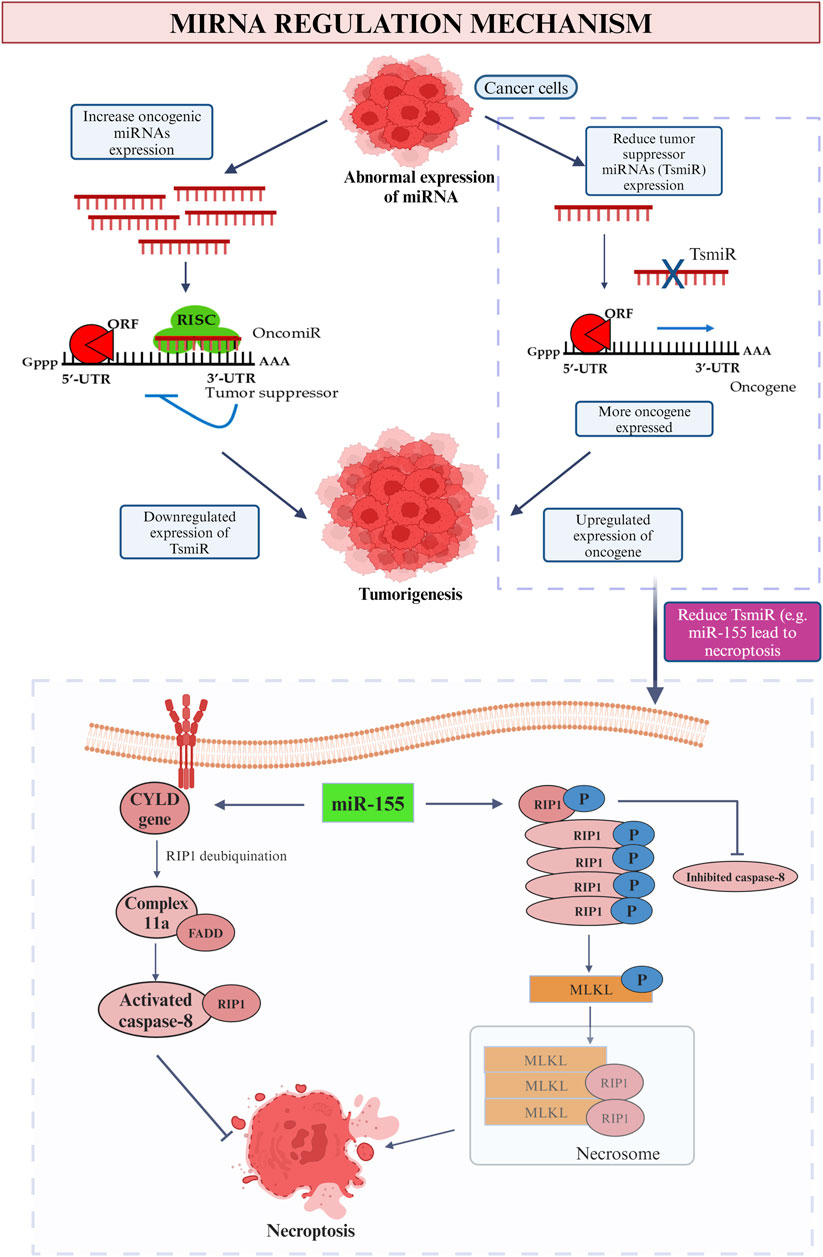
There are numerous miRNAs have been studied previously and it was found that they are capable in both enhancing and impeding tumor development, depends on the types and roles of each miRNA. Many of them are proven to be related to invasion, prognosis and the formation of new blood vessels (Hussen et al., 2021). The miRNA that is involved in breast cancer can be categorized into few classes such as the oncogenic miRNA and tumor suppressor miRNA (Figure 8). Subdivision can be continued from both classes into various families. These miRNAs are derived from both the tumor and stroma cells to perform their role in inhibiting the immune system (Wang et al., 2022b). Positive regulation of oncogenic miRNAs hinder the tumor suppressor gene thus allow the formation of tumor while in contra to it, negative modulation of miRNAs with tumor suppressor effect will improve the oncogenes translation (Zacharias et al., 2020). As an example of tumor suppressor miRNA in suppressing the cell proliferation, miR-340 is one of the best instance by which it obstructs the Wnt appearance by aiming LGR5 and FHL2 thus inhibit the cell proliferation (Otmani and Lewalle, 2021).
Figure 8. MiRNA regulation mechanism. This picture depicted the role of oncogenic and tumor suppressor miRNA in cancer and the influence on tumor suppressor miRNA (miR-155) on promotion and inhibition of necroptosis.
In the context of promoting malignancy, oncogene miRNA is responsible to inhibit the function of the specific tumor suppressor genes thus boosting the cancer progression and promote breast malignancy. Meanwhile, tumor suppressor miRNA restrains the oncogenes that related in breast tumorigenesis thus downregulate and leads to breast cancer (Saikia et al., 2020). In a nutshell, irregular miRNAs will be found in the appearance of cancer cell. Increase level of oncogenic mRNA results in lesser manifesting of tumor suppressor gene while due to the low level of tumor suppressor miRNAs, positive feedback possessed onto the appearance of oncogene. Both of the feedbacks lead to tumorigenesis which involves the process of cell proliferation, invasion, prevent apoptotic event, angiogenesis, replicative immortality and metastasis (Loh et al., 2019). On the contrary, with the purpose to inhibit the breast cancer development, certain miRNAs may be participated in the replacement therapies which hamper the appearance of oncogenes (Smolarz et al., 2022). To be more persuading, a research held in 2014 found that patients who diagnosed with early stage breast cancer will have increased level of miR-155, miR-19a, miR-181b and miR-24 as compared to normal healthy people (Hamam et al., 2017).
As an example of oncogenic miRNA, high level of miR-155 play an antitumor role in breast cancer through the directly suppressing of cytokine signaling (SOCS1) which aids in the modulation of IFN-y production in T cells is vital to assemble and activate the adequate amount of T effector cells (Chen et al., 2020). In breast cancer, it is the dominant regulator in metabolizing glucose hence able to promote the energy metabolism through few pathways. Moreover, the coordination between NF-κB-miR-155 axis and NF-κB-miR-146a axis may also upregulating the inflammatory reaction (Hoorzad et al., 2023). The above founding all indicate high level of miR-155 is potent as a role that inhibiting the activity of immune response. However, it is also crucial in the first line differentiation of myeloid progenitors, responsible for various cellular processes such like proliferation, stemness, apoptosis and angiogenesis, dysregulation of it may still result in hematologic problem and exerting its oncogenic natural in solid tumors such as breast cancer (Kalkusova et al., 2022).
On the other hand, tumor suppressor miRNAs such as miR-23a/27a/24-2, miR-146a and miR-223a also possessed similar responsibilities but in a different alternative. As an instance, miR-146a is the main component that responsible to downregulate the differentiation of Th1 cell (Testa et al., 2017). Along with the elevating level of IFN-y in miR-146a, destruction is able to be seen on both CD4+ and CD8+ T cells (Mastroianni et al., 2019). When the signal transducer and activator of transcription one is not able to be targeted, the Treg cells will no longer capable of impeding the Th1 response which triggered by IFN (Goropevšek et al., 2017). Assemble of innate immune cells, autoantibodies as well as triggered CD4+ and CD8+ T cells were found to result in autoimmune disorders and severe hyperinflammatory (Li B. et al., 2017). Moreover, with the stimulation of T-cells, it negatively responses in limiting the NF-kB activity through the suppression of NF-kB activators (Wang et al., 2019). This communication is proved to reduce the death in T-cells that enhanced by activation.
Furthermore, elevate expression of miR-494 in myeloid-derived suppressor cells (MDSCs) diminish the quantity of PTEN thus result in the accumulation of MDSCs and help in the invasion and metastasis of breast tumor. In normal situation, PTEN represents the major pathway to regulate the activation and reproduction of several immune cells including T- and B- cells (Taylor et al., 2019). Hence, it is reduced in this case via the lipid phosphatase and protein phosphatase activities to diminish its ability in producing divergent tumor suppressing genes (Chen et al., 2022). When PTEN is not enough to construct the gene that suppresses the tumor growth, then a high amount of MDSCs will be collected in the respective area. MDSCs have potential to impede the T cells activities thus create a favorable condition for tumor to develop and proliferate without any interruption. Meanwhile, it also improves the tumor with resistance to immunotherapy therefore its survival is even assured (Law et al., 2020).
Meanwhile, miR-34a is a well-known miRNA that can be simply detected in every healthy cell with the responsibility to repress various tumors which comprise the breast, colon, prostate cancer and more. According to Rui et al. (2018), they suggested that the elevated the number of miR-34a, the more the breast cancer is under controlled (Rui et al., 2018). A series of assessments have been performed in order to support their statement which include the use of TargetScan device to expose the effect of miR-34a towards Notch1. In a more particular way, Notch1 is an elementary vital option that contribute in the emergence of breast tumor as it consists of Delta-like-4 that lead to the formation of new useless blood vessels in the malignancy conditions when the Delta-like-4 is being downregulated (Kontomanolis et al., 2018). It is crucial to modulate the proliferation of breast tumor. Besides, MTT examination states that the survival rate of the breast cancer cell line, MCF-7 cell is apparently reduced by miR-34a. It prompts the abnormal cells that can be found in breast to perform apoptosis and some of the malignant cells are arrested in G1 phase to prevent the further atypical replication of mutated DNA (Ngabire et al., 2018). With this, it is strong evidence showing that increasing amount of miR-34a is highly beneficial in treating and inhibiting breast cancer. It may be used as a therapeutic agent to cure the breast cancer.
Moreover, let-7 family that can be counted as a member of miRNA which is found ahead of the other miRNAs play an important role to suppress numerous cancer development. The name lethal-7 is given as its evolution often result in death of the patient (Bernstein et al., 2021). It is made up of 22 nucleotides but does not possess the ability to encode a protein. Meantime, it is a big family which act as an originate for nine miRNAs which are the let-7a, let-7b, let-7c, let-7d, let-7e, let-7f, let-7g, let-7i and miR-98 (Mizuno et al., 2018). Out of various functions that can be performed by this big family, they also capable of restricting the growth of breast cancer via impeding the proliferation, metastases and invasion of the abnormal cells by targeting MAGE-A (Liu et al., 2019). MAGE-A is also known as Melanoma-associated antigen-A which is an antigen that is noticed with the presence of tumor. These antigens will be noted by the cytotoxic T cells thus lead to the activation of a strong specific immune response that fights against the breast tumor. Elevated appearance of MAGE-A genes always indicating that the patient may have a reduced survival rate from breast cancer (Poojary et al., 2020). Based on the declaration by (121), miRNA let-7a also able to decrease the presence of E2F2 protein in order to inhibit the aberrant reproduction of osteosarcoma cells, which is the proliferation of high-grade malignant mesenchymal cancer cell that derived from the abnormal bone and osteoid (Dekkers et al., 2019). The E2F transcription factor 2 (E2F2) that stimulated by the infected E2F or Myc proteins is vital for tumorigenesis by which high expression of E2F2 may assist in high grade tumor, metastasis of cancer cell or any other terrible prognosis (Świętek et al., 2023).
MiRNA regulated cancer mediated immune cell deathImmune cell death may occur in two approaches which include the death of cell, apoptosis and destruction of the body tissues, necrosis when there is only inadequate blood flows to the particular area (Khalid and Azimpouran, 2023). Apoptosis is commonly a pathway that rigorously performed by every cells each time when the cells reach to the limit of their lifespan but the condition become out of control in the presence of cancer cells (Shirjang et al., 2019). As explained by Taghavipour et al. (2020), miRNAs able to enhance and reduce the chance of apoptosis through two different alte
留言 (0)