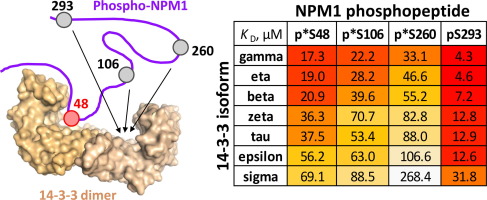
Improving the stability of a protein without impacting its desired function is a fundamental objective for protein engineering and design. Functional proteins often have strong constraints on their sequence. While approximately 5% of randomly sampled mutations can improve protein stability, far fewer do so without disrupting function1, 2. As the benefits of each stabilizing mutation is small, whereas even a single destabilizing mutation can be highly detrimental, the protein designer must ‘shoot the moon’ by getting nearly all chosen mutations correctly. Earlier efforts incorporated evolutionary information in the form of consensus mutations or substitutions sampled often in the evolutionary history of the protein family3, 4, 5. More recently, protein design approaches have made incredible advances in predictions of changes in protein stability6, 7, 8. Many algorithms, most notably PROSS9, 10, sample only from a set of substitutions commonly seen throughout evolution. Such approaches have been successful for stabilizing many classes of proteins, including enzymes, transporters, and binding proteins11, 12, 13. Still, identifying positions not to mutate and sets of substitutions that do not disrupt function is often performed empirically or with heuristics that may not transfer across to other systems. Such heuristics, like evolutionary conservation, may fail when applying design to engineered proteins with functional constraints that have not undergone natural evolution.
We have recently engineered over twenty new genetically encoded biosensors using the chemically induced dimerization PYR1-HAB1 partner proteins[14]. In plants, PYR1 is a soluble, 27 kDa receptor which recognizes the hormone abscisic acid (ABA)[15]. Upon noncovalent binding of ABA, PYR1 undergoes a subtle yet important conformational change that results in potent recognition of the PP2C phosphatase HAB1[16]. Extensive genetic analyses have revealed many mutations on PYR1 which result in constitutive binding to HAB1[17]. These mutations occur at the HAB1 binding interface, but also at distal positions. Thus, PYR1 is quite sensitive to mutations which disrupt function, hampering protein engineering and design efforts. It is unknown whether the conformation-induced mechanism is identical between the native and the engineered biosensors. One current limitation in developing new sensors and enhancing existing ones is stability. It would be laborious to stabilize all sensors individually and thus highly advantageous to find mutations that would stabilize the platform across more already-developed sensors.
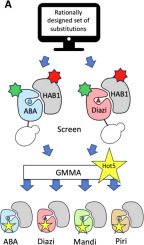
In this work, we identified amino acid substitutions that stabilized several already-developed PYR1 sensors of different specificity. We used a combined experimental and computational global multi-mutant analysis (GMMA), which has previously been shown to identify multiple substitutions that together progressively enhance a protein’s stability also under functional constraints 18, 19. The ability to identify substitutions that work together is a result of analyzing variants with multiple amino acid substitutions where the beneficial effect is observed in various slightly different backgrounds. We hypothesized that this reduced sensitivity towards the sequence background would also enable GMMA to identify substitutions that will enhance PYR1 across sensors of different specificity, i.e. with slightly different functional constraints.
Experimental screening of libraries of multiple-substituted variants provides a rich source of information where the effect of each amino acid substitution may be represented in hundreds of measurements and stability is probed by the ability to rescue the relevant function from inactivation due to other destabilizing substitutions. The task then remains to disentangle individual substitution effects from the web of protein variants interconnected by sharing the same amino acid substitutions. GMMA achieves this by adopting a sequence-function relationship that involves a linear function, F, that is translated to the assay readout, A, via a non-linear function, here a simple sigmoid:Fv=Fref+∑s∈vΔFs;Av∝11+expβFvHere, a protein variant, v, consists of several amino acid substitutions, s, and β is a constant. Different flavors of such models have been shown to describe high-throughput screens well 2, 18, 20, 21, 22, 23 and it has been observed that the linear scale correlates well with thermodynamic stability although also containing aspects of function 19, 24, 25. The linear dimension, F, may be extended with higher-order terms, e.g. to model spatial interactions between specific positions, but here we are mainly interested in additive effects and assume that higher order effects are effectively averaged out if a substitution is observed in more than 20 mutational backgrounds. Further, several studies have demonstrated that a majority of variants have fundamentally additive effects on protein function and stability 24, 25, 26.
Applying GMMA to data generated by yeast display of PYR1 combinatorial mutagenesis libraries, we investigated the same sets of substitutions in two different sensor backgrounds and found a common set of stabilizing substitutions between the two sensors. We show that when these substitutions are incorporated into diverse sensors, they can also stabilize these proteins while not impacting in vivo and in vitro function. This showcases that both the stabilizing effect and functional constraints identified by analyzing multi-mutants may indeed be transferable between slightly different sequence backgrounds.





留言 (0)