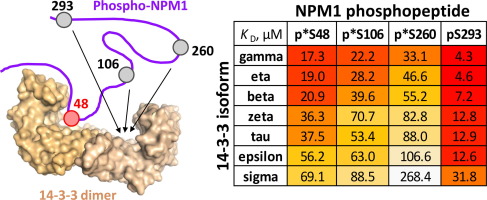
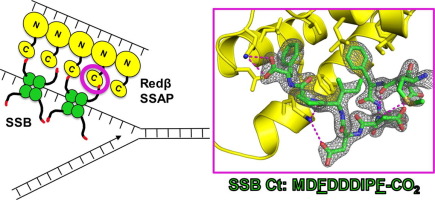
Structural basis for the interaction of Redβ single strand annealing protein with Escherichia coli single-stranded DNA binding protein
Redβ is a 261-amino acid (Mr 29.6 kDa) protein from bacteriophage λ that binds to single-stranded DNA (ssDNA) to promote the annealing of complementary strands[1], [2]. It functions together with an exonuclease (λ-exo) as part of a two-component DNA recombination system with potential roles in replication, genome packaging, repair of dsDNA breaks, and promotion of genetic diversity[3], [4]. A schematic overview of the system is shown in Figure 1. First, λ-exo binds to double-stranded DNA (dsDNA) ends and resects the 5’-strand to form a long 3’-overhang[5], [6], [7]. Redβ then assembles on the 3’-overhang to promote its annealing with a complementary DNA strand. The two proteins form a complex that is thought to mediate loading of Redβ directly onto the 3’-overhang as it is generated by λ-exo[8], [9]. Binding of Redβ to λ-exo is mediated by its C-terminal domain (CTD, residues 194-261), whereas its N-terminal domain (NTD, residues 1-177) is responsible for DNA binding and oligomerization[10], [11].
Homologs of λ-exo and Redβ are encoded in the genomes of a wide variety of bacteriophage with dsDNA genomes12. While their precise roles for phage propagation have been elusive, the proteins have been exploited in powerful methods for bacterial genome engineering known as recombineering and multiplexed automated genome engineering (MAGE)[13], [14], [15]. In one type of recombineering called single-strand oligonucleotide repair (ssOR), a synthetic oligonucleotide that is complementary to a target site but encodes an alteration is electroporated into E. coli cells expressing Redβ. Redβ then binds the oligonucleotide and anneals it to a target site exposed as ssDNA at the lagging strand of a replication fork[16], [17]. Redβ forms a protein-protein interaction with E. coli SSB protein18, which coats the ssDNA at the lagging strand to protect it from degradation and coordinate access of numerous proteins involved in replication[19], [20], [21] (Figure 1C). Mutational analysis indicates that this interaction involves the acidic C-terminal tail of SSB (C-tail) and a site on Redβ CTD that overlaps with its λ-exo binding site18. Mutations that disrupt the Redβ-SSB interaction completely abolish oligonucleotide recombination in vivo18. This suggests that for a Redβ–ssDNA complex to gain access to the complementary target site on the lagging strand, it must interact with the SSB C-tail to either displace SSB and expose the ssDNA at the target site or co-localize to the lagging strand and compete with SSB for the target site.
Here, we have determined the x-ray crystal structure of the Redβ CTD in complex with a peptide corresponding to the last nine residues of SSB (SSB-Ct, residues 169-177, MDFDDDIPF). All nine residues of the SSB peptide are ordered in the complex and bind to the expected hydrophobic groove on the surface of the CTD, largely through the insertion of two phenylalanine side chains of SSB (Phe-171 and Phe-177), as well as electrostatic interactions with the SSB C-terminal carboxylate. Mutation of residues forming key interactions in the structure to alanine significantly disrupts the complex formed by the full-length proteins, suggesting that the interactions seen in the crystal structure are functionally relevant. A dimeric form of the CTD–SSB-Ct complex was observed in all four crystal forms, but native mass spectrometry (nMS) did not detect the presence of the dimeric complex in solution. nMS also revealed that the complex between the CTD and the SSB-Ct peptide is weak, suggesting that further interactions or oligomeric assemblies may be relevant in vivo. Structural knowledge of the complex helps to explain prior data on the ability of Redβ and orthologous proteins to operate in bacterial hosts with different C-terminal sequences of their SSB proteins22.





留言 (0)