
記住我
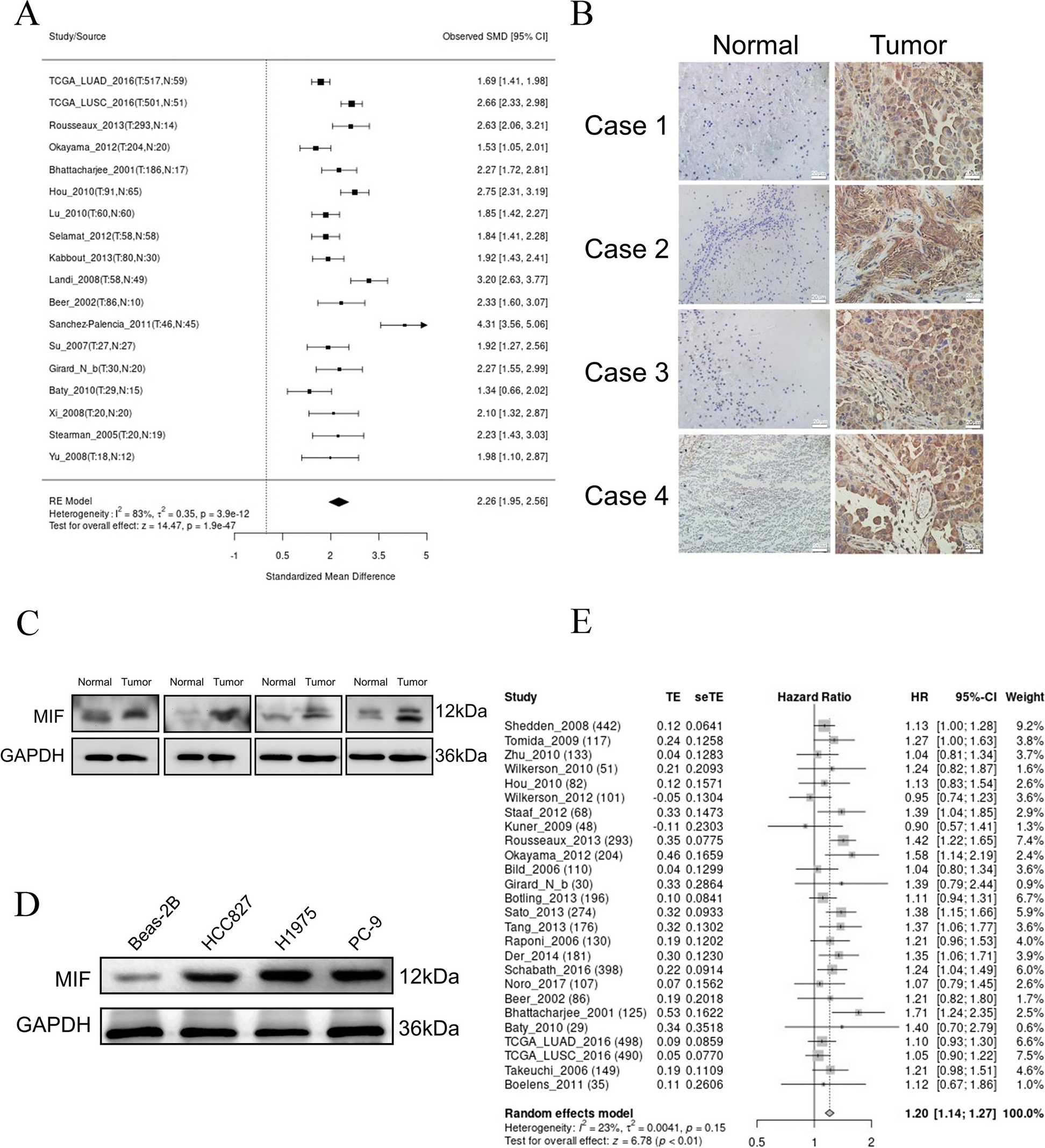





In the previous integration analysis of significantly mutated genes, we highlighted the effect of the post-transcriptional modification gene set on CCA development (Fig. 1. A). Notably, in our cohort, approximately 7.5% (5/67) of CCA patients harbored RBM10 mutations, an alternative splicing factor involved in RNA maturation. The same result was validated in a large-scale pan-cancer Chinese population by exome sequencing [17]. The mutational landscape of RBM10 in solid tumors revealed high mutation rates in both extrahepatic cholangiocarcinoma (7.4%, 26/351) and intrahepatic cholangiocarcinoma (6.3%, 32/508) in the Chinese populations(Fig. 1. B), whereas Western populations exhibited lower frequencies (0%, 0/36 in the TCGA population; 0.5%, 1/200 in the MSK-IMPACT population [18]) (Fig S1. A & B), hinting at a potential specificity of RBM10 mutations to Chinese CCA patients. Moreover, the RBM10-alter group showed a higher mutation count (p = 7.99e-7) and tumor mutation burden (p = 2.79e-7) than the unaltered group (Fig S1. D & E). We further analyzed the mutation patterns in the RBM10 alter group in Chinese CCA population cohort: classic tumor driver genes (KRAS, SMAD4, CTNNB1, TGFBR2, SF3B1, etc. P < 0.05) were more prone to co-occur in the RBM10 alter group (Fig. 1. C-D); the majority of RBM10 mutations were protein-truncating variants, including splice site, nonsense, and frameshift mutations (Fig. 1. E). Owing to the same high mutation rate of RBM10 in LUAD, RBM10 has been investigated and is suggested to play an important role in LUAD [19]. The RBM10 mutation pattern in LUAD mirrored those in CCA(Fig S1. C), resulting in RBM10 expression decreasing (Fig S1. F). RBM10 mutations also comprised a substantial proportion of missense mutations in CCA and LUAD, whose functional impacts were mostly ambiguous and needed experimental validation. Within our initial data, we detected three missense mutations, one nonsense mutation, and one splice site mutation. The three missense mutations affected two crucial domains (R305H and E355V in the second RNA recognition motif; C761Y in the C2H2-type Zinc finger domain) (Fig. 1. F). The effects of mutations in the second RNA recognition motif have been reported [11, 20], whereas the consequences of alterations in the C2H2-type zinc finger domain remain largely unexplored. Consequently, the in-silico pathogenicity prediction also indicated that RBM10C761Y occurring in the C2H2-type zinc finger domain by PROVEAN and SIFT respectively, deserved further investigation (Fig. 1. G).
Fig. 1
Mutation spectrum of RBM10 in CCA. A Functional cluster analysis was conducted in our previously identified potential driver genes in CCA. The gene symbols in red represented the CCA-related genes that were identified in our previous study. Gene symbols in black were previously reported in CCA. NMU, our own CCA cohort (67 cases); ICGC, International Cancer Genome Consortium (179 cases); NC Zou et al.’s study [38] (102 cases). B RBM10 alteration frequency in pan-cancer in Chinese population from Cibioportal. C Top 30 Concurrent gene alterations in CCA patients with RBM10 mutations in the Chinese population. D Significant alteration rate difference between RBM10 alter and unalter groups in CCA (P < 0.05). Blue column RBM10 unalter group; red column, RBM10 alter group. E RBM10 point mutation distribution in 58 of 859 CCA samples in the Chinese population. Lollipop plot showed mutations throughout the RBM10 gene area, using MutationMapper. Green rectangle, two RNA recognition motifs (RRM); red rectangle, two zinc fingers (left, C4-type zinc finger domain; Right, C2H2-type zinc finger domain); purple rectangle, Octamer repeat domain(OCRE). F RBM10 point mutation distribution in 5 of 67 CCA samples from our dataset. G Pathogenicity prediction of mutations in our cohort using PROVEAN and SIFT
RBM10 wild-type was downregulated in CCA and indicated worse survivalTo elucidate the role of RBM10 mutations in CCA development, the clinical associations of wild-type RBM10 expression were initially examined in CCA. Analysis of RBM10 mRNA expression in 64 CCA patients revealed elevated levels in adjacent normal tissues compared to CCA tissues (Fig. 2. A & B). We subsequently assessed the protein expression of RBM10 in CCA tissue microarrays by immunohistochemistry (IHC) and observed the same trend. The representative images of RBM10 expression in tumor and para-tumor tissues were displayed (Fig. 2. C). Follow-up data based on CCA tissue microarrays confirmed RBM10 downregulation in tumors correlating with reduced overall survival (P = 0.0487) and recurrence-free survival (P = 0.0207) (Fig. 2. D & E), suggesting RBM10’s potential as a tumor suppressor gene in CCA. Furthermore, the multivariate analysis showed that RBM10 wild-type expression was an independent predictive factor for postoperative overall survival (P = 0.039) (Fig S1. G).
Fig. 2
RBM10 was downregulated in CCA. A The different levels of RBM10 mRNA expression in paired CCA and adjacent tissues (n = 64). B The individual RBM10 mRNA expression difference distribution between tumor and para-tumor. C Representative IHC stains of RBM10 in CCA and adjacent normal tissues. D, E The prognostic value of RBM10 expression in CCA was evaluated by Kaplan-Meier analysis of overall survival (OS) and recurrence-free survival (RFS) according to the IHC staining intensity of RBM10 in TMA (n = 177). Blue line, high RBM10 expression in CCA; red line, low RBM10 expression in CCA
RBM10C761Y was identified as a loss-of-function mutation in CCATo mitigate the influence of background RBM10, we assessed the expression levels of RBM10 in four CCA cell lines (QBC939, RBE, HuCCT1, and HCCC9810) and one normal bile duct cell line (HiBEC) before establishing experimental cell models. We selected two cell lines, QBC939 and RBE, which exhibited the lower expression levels of RBM10 (Fig S2. A & B). Then, we establish two stable RBM10WT/ RBM10C761Y overexpressing CCA cell lines. The Sanger sequencing was performed to confirm the absence of RBM10C761Y mutations in wild-type CCA cell lines (RBE and QBC939) (Fig. 3. A) and the transduction efficiency of lentiviral vectors was evaluated at RNA and protein levels (Fig. 3. B & C). Additionally, the C761Y mutation was detected exclusively in the MUT group by designed mutation-specific primers (Fig S2. C & D). We observed fewer EdU-positive cells in the wild-type (WT) group than in the negative control (NC) group. However, the mutant (MUT) group significantly restored this ability (Fig. 3. D). The cell viability of CCA cells was assessed by Cell Counting Kit-8 (CCK-8) assay, which showed that the WT group exhibited lower cell viability than the NC group, while the MUT group showed no significant difference from the NC group (Fig. 3. E). Colony formation assay demonstrated the consistent alteration of proliferation ability (Fig. 3. F). In addition, overexpression of wild-type RBM10 inhibited CCA cell migration. By contrast, RBM10C761Y lost the tumor-suppressive functions, as shown by the transwell and wound healing assay (Fig. 3. G&H).
Fig. 3
RBM10 was a tumor suppressor and RBM10C761Y was a loss-of-function mutation in CCA. A Sanger sequencing plot of interested region including RBM10 C761Y in QBC939 and RBE. B, C Establishment of stable WT and MUT cells was evaluated at mRNA and protein levels. NC, negative control; WT, wild-type; MUT, mutant. D, E, F Proliferation ability of different groups was examined by EdU, CCK-8, and colony formation assay. G, H Migration ability of different groups was detected by wound-healing and transwell assay. I Photograph of excised tumors from mice in different groups (n = 5). J Representative images showed H&E and IHC staining for RBM10 and Ki67 in tumors removed from nude mice
To elucidate the in vivo functions of RBM10WT and RBM10C761Y in CCA development, these CCA cells were inoculated subcutaneously into nude mice. Xenografts derived from QBC939 cells with stable wild-type RBM10 overexpression were smaller than those from control QBC939 cells. In agreement with the in vitro findings, the suppressive effects of RBM10C761Y on tumor growth were significantly reduced in vivo compared with those of RBM10WT (Fig. 3. I). The expression of Ki67 was decreased in the WT group, while no significant change was observed in the MUT group compared with the NC group (Fig. 3. J). Combined, these data suggested that the loss-of-function mutation RBM10C761Y impaired the tumor suppressive role of wild-type RBM10 in CCA.
ASPM alternative splicing served as the target of RBM10RBM10 modulated the alternative splicing of thousands of target genes and may influence the progression of CCA by altering the splicing patterns of cancer-related genes. To gain a deeper insight into the mechanism by which RBM10WT and RBM10C761Y functioned in CCA development, we performed RNA sequencing in the NC, WT, and MUT groups. Splicing changes caused by RBM10C761Y in CCA cell lines were analyzed using the rMATS pipeline, which identified five types of alternative splicing events: retained intron, mutually exclusive exons, alternative 3’ splice site, alternative 5’ splice site, and skipped exon. The most obvious changes were exon skipping (ES) events, which were also the most prevalent events involving RBM10 (Fig. 4. A). Compared to the WT group, 1197 significant ES events (false discovery rate (FDR) < 0.01) were defined as RBM10C761Y-associated ES events (Fig. 4. B). Based on RBM10 cross-linking immunoprecipitation (CLIP) sequencing data [21, 22], inclusion level difference between the two groups and ES events count, we excluded 975, 80, and 54 ES events respectively. The remaining 88 ES events were defined as RBM10C761Y-modulated ES events. The 18th exon skipping of assembly factor for spindle microtubules (ASPM) was among the top significant ES events (Fig. 4. C & Table S1). To minimize the confounding effects of different cell lines on CLIP sequencing, we also identified SMN2 exon6 event as potential targets of RBM10C761Y regulation by integrating the rMATS analysis results and the changes of the predominant transcripts in the sequencing results, regardless of the CLIP results (Fig S3. A & Table S2). In both the WT and MUT groups, we employed RNA immunoprecipitation experiments to determine which gene specifically binds to RBM10. The experimental results revealed that ASPM interacts with both the wild-type RBM10 and the C761Y mutant, however, the binding capacity of the C761Y mutant to ASPM was diminished (Fig. 4. D & E). These results implied that RBM10 may be involved in the ASPM ES event and C761Y mutant impaired the interaction between ASPM and RBM10.
Fig. 4
RBM10 modulated alternative splicing of ASPM in CCA. A The number of different alternative splicing events associated with RBM10C761Y mutation in CCA. RI, retained intron; MXE, mutually exclusive exon; A3SS, alternative 3’ splice site; A5SS alternative 5’ splice site; SE, skipped exon. B Significant exon skipping (ES) changes between RBM10 WT and MUT groups (FDR < 0.01, |inclusion level differences| > 0.1). C A flowchart of exploring RBM10 mutation-related ES events. D, E RNA immunoprecipitation using anti-RBM10 or anti-SMN2 antibodies, or igG in different groups. RNA immunoprecipitation (RIP) products were analyzed by RT-PCR. The recovered RNA in each RIP was normalized to its input. F, G Isoforms expression of ASPM in NC, WT, and MUT groups were evaluated at mRNA and protein levels. Green rectangle in (G), CAMSAP CH domain; red rectangle in (G), isoleucine and glutamine domain. H RT-PCR results revealed that the relative expression of ASPM203/ASPM201 was negatively related to RBM10 expression in CCA samples(n = 77) by Pearson rank correlation analysis (r=-0.2989, P = 0.0074). I The effects of RBM10 WT and MUT on ASPM exon18 ES event were examined using minigene splicing reporter assays
The ASPM exon18 ES event resulted in two different transcripts (ASPM203 and ASPM201). The changes of ASPM203/ASPM201 were evaluated at RNA and protein levels in three groups of cells. The expression of ASPM203 in the WT group was lower than that in the NC group, while the MUT group did not show significant changes compared to the NC group (Fig. 4. F & G). Further analysis of RNA from CCA tissues revealed that the ratio of ASPM203 to ASPM201 was negatively correlated with the expression of RBM10 (Fig. 4. H P = 0.0074, r=-0.2989). We also performed ASPM minigene splicing reporter assays to explore the effects of the mutations on ASPM ES event. Compared with the NC group, the WT group showed significant changes in the splicing of ASPM minigene, while the MUT group did not affect the splicing (Fig. 4. I). Together, these observations demonstrate the ASPM exon18 ES events may serve as the target of RBM10.
RBM10C761Y reduced interaction with SRSF2 and influenced the ASPM ES eventTo further understand how RBM10C761Y mutation affected the ASPM ES event, the ASPM pull-down assay was conducted to explore the proteins involved in this ES event. However, RBM10 was not detected in the isolated pull-down products (Fig. 5. A). To identify more alternative splicing factors in the event, we further conducted silver staining and mass spectrometry (MS) analysis on the pull-down products (Fig. 5. B). There were 254 binding proteins in the positive probe group (Fig. 5. C). We focused on spliceosome-related proteins and found U2AF2 and SRSF2 as two potential candidates, which played important roles in alternative splicing [23, 24] (Fig. 5. D & E). RNA pull-down assay indicated that SRSF2 bound to ASPM minigene (Fig. 5. F). Co-immunoprecipitation experiments revealed that SRSF2 could form a complex with RBM10 wild type, while the mutant binding ability decreased in two cell lines (Fig. 5. G & H). Further RNA immunoprecipitation experiments showed that SRSF2 can bind to ASPM, and knocking down SRSF2 significantly reduced the binding ability of RBM10 to ASPM (Fig. 5. I). SRSF2 knockdown increased the ratio of ASPM203 to ASPM201 in the WT group at the RNA level (Fig. 5. J). The ASPM minigene splicing reporter assays also revealed that knocking down SRSF2 impaired the ability of the RBM10 wild type to regulate the splicing event of ASPM minigene (Fig. 5. K). The above results suggested that RBM10 interacted with SRSF2 and functioned in the ASPM exon18 ES event, while RBM10C761Y reduced the connection.
Fig. 5
RBM10 and SRSF2 cooperatively controlled the splicing of ASPM. A Immunoblot analyses of RBM10 in ASPM-pulldowns derived from the WT and MUT groups transfected with ASPM-minigene constructs. B Sliver staining of ASPM-pulldowns. MS analysis was performed in WT groups. C Venn diagram of ASPM-pulldowns between positive and negative probes. D Pathway enrichment analysis of positive probe unique ASPM-pulldowns. E Potential spliceosome factors involved in ASPM ES. F Immunoblot analyses of U2AF2 and SRSF2 in ASPM-pulldowns. G, H Co-immunoprecipitation was performed to confirm the interaction between SRSF2 and RBM10/RBM10C761Y. RBM10 mutation weakened the interaction between SRSF2 and RBM10. I RNA immunoprecipitation using anti-RBM10 or anti-SRSF2 antibodies, or igG with SRSF2 knockdown. J RT-PCR was performed to detect the expression of ASPM isoforms in NC, WT, MUT, and SRSF2-knockdown groups. K Effects of RBM10 and SRSF2 on ASPM exon18 ES event were examined using minigene splicing reporter assays
RBM10C761Y-modulated ASPM203 involved in CCA developmentTo verify the role of ASPM203 in RBM10C761Y-mediated CCA progression, we silenced ASPM203 in the NC and MUT groups using siRNA. Silencing ASPM203 in the NC group reduced the proliferation and migration of CCA cell lines (Fig S4. A & B). We also found that ASPM mutations in TCGA CCA populations were all amplification mutations (Fig S4. C), suggesting that ASPM may facilitate the development of CCA. The mean percent spliced-in index of ASPM203 in pan-cancer was above 0.8, indicating that ASPM203 might be involved in cancer progression (Fig S4. D). The use of RNAi targeting exon18 sequences did not affect the expression of ASPM201(Fig. 6. A & B). EdU staining demonstrated that the knockdown of ASPM203 inhibited the proliferation of the MUT group (Fig. 6. C). Colony formation and CCK-8 assay also showed that the MUT group regained tumor suppressor ability by downregulating ASPM203 (Fig. 6. D & E). The same effect was observed in wound healing and transwell assay (Fig. 6F & G). These results suggested that RBM10C761Y promoted CCA progression depending on ASPM203.
Fig. 6
RBM10C761Y-mediated oncogenic ASPM isoforms involved in CCA development. A, B ASPM isoform expressions were examined in the MUT group with specific ASPM203 knockdown at mRNA and protein levels. C Edu assay, D growth curve, E and colony formation assay were performed to assess the effects on the proliferation capacity of CCA cell lines after knocking down ASPM203 in the MUT group. F Wound healing assay and G transwell assay were performed to assess the effects on the migration capacity of CCA cell lines after knocking down ASPM203 in the MUT group. Si-203: Si-ASPM203
ASPM203 interacted with DVL2 to enhance Wnt/β-catenin signalingTo explore the regulatory mechanism of ASPM203 in CCA, we analyzed the transcriptome sequencing data of the WT and MUT groups. GO term analysis showed that transcription regulation was also upregulated in the MUT group (Fig S4. F). Pathway enrichment analysis revealed that the Wnt/β-catenin signaling pathway was significantly upregulated in the MUT group compared with the WT group (Fig S4. G). Previous studies have shown that ASPM203 interacted with dishevelled (DVL) protein to participate in cancerogenesis [25, 26], which suggested that ASPM203 bound to DVL protein, preventing degradation and thus enhancing the Wnt/β-catenin signaling pathway (Fig. 7. A). Notably, co-immunoprecipitation assay revealed that ASPM was associated with DVL2 but not with DVL3 in CCA cells (Fig. 7. D & S3. E). We analyzed the changes of Wnt-related genes in NC, WT, and MUT groups and found that CTNNB1 (β-catenin) and DVL2 did not change at the mRNA level, while CCND1 (cyclin D1) decreased at the transcription level in the WT, and MUT group had no difference with NC group (Fig. 7. B). At the protein level, DVL2, β-catenin, cyclin D1, and N-cadherin all decreased in the WT group, while the MUT group had no difference with the NC group (Fig. 7. C). When knocking down ASPM203 in the NC group, DVL2 and CTNNB1 did not change significantly at the transcription level, but decreased significantly at the protein level (Fig. 7. E & F). Further, knocking down ASPM203 in the MUT group resulted in similar changes to the WT group at protein level(Fig. 7. G). Immunofluorescence experiments also showed that nuclear β-catenin was reduced in the WT group compared to the NC group, while there was no significant change in the MUT group, but knockdown of ASPM203 in the MUT group reduced the nucleus translocation of β-catenin (Fig S5. A). The main form of DVL2 protein degradation is the ubiquitin-proteasome pathway [27]. Subsequently, we transfected HA-tagged ubiquitin into cells that were treated with MG132 to inhibit DVL2 degradation. When immunoprecipitated with DVL2 antibody, the level of ubiquitinated DVL2 was increased in the WT group, while there was no difference between the MUT group and the NC group. Conversely, the knockdown of aspm203 in the MUT group resulted in the loss of the ability to protect DVL2 from degradation (Fig. 7. H). These results suggested that ASPM203 bound to DVL2 and reduced its degradation, thereby enhancing the Wnt signaling pathway.
Fig. 7
ASPM203 interacted with DVL2 to facilitate Wnt/β-catenin signaling. A The diagram illustrates the role of ASPM in regulating the Wnt signaling pathway, as reported in previous studies. B Co-immunoprecipitation was performed to verify the physical interaction between ASPM203 and dishevelled-2 (DVL2), a key mediator of Wnt/β-catenin signaling. C, D Using RT-PCR and western blot to determine the different levels of Wnt/β-catenin signaling associated genes in the NC, WT, and MUT groups. E, F Using RT-PCR and western blot to determine the different levels of Wnt/β-catenin signaling associated genes with knocking down of ASPM203 in the NC group. G, H Expression and ubiquitylation of DVL2 were detected in four groups, including the NC, WT, MUT, and ASPM203-knockdown groups
RBM10C761Y -modulated ASPM203 promoted CCA progression in a Wnt/β-catenin signaling dependent mannerTo explore how RBM10C761Y modulated ASPM203 and enhanced CCA development in a β-catenin signaling dependent manner, we investigated the effect of ASPM203 silencing and DVL2 upregulation on CCA cell proliferation in two cell lines. In the MUT group, ASPM203 silencing significantly reduced cell proliferation, while in the DVL2 upregulation restored it, as shown by the EdU, CCk8, and colony formation assay (Fig. 8. A, B & C). In addition, we examined the effect of ASPM203 silencing and DVL2 upregulation on CCA cell migration in the MUT group: ASPM203 silencing abolished it, while DVL2 upregulation induced it, as evidenced by the wound healing and transwell assay (Fig. 8. D & E). The effect of DVL2 overexpression on β-catenin protein level was investigated in the ASPM203-silenced group: DVL2 overexpression could recover β-catenin protein level, as shown by the Western blot analysis (Fig. 8. F). The above results indicated that the C761Y mutation reduced the binding ability of RBM10 and SRSF2, leading to a decrease in ASPM exon18 ES event, an increase in ASPM203 expression, an enhancement of DVL2 stability, an upregulation of Wnt pathway, and a promotion of cholangiocarcinoma progression (Fig. 8. G).
Fig. 8
ASPM203 promoted CCA progression in a Wnt/β-catenin signaling dependent manner. A, B, C The alterations of CCA cell lines proliferation were evaluated by EdU, CCK-8, and colony formation assay in MUT, ASPM203-knockdown, and ASPM203-knockdown with DVL2-overexpression groups. D, E The alterations of CCA cell line migration were assessed by wound healing and transwell assay in MUT, ASPM203-knockdown, and ASPM203-knockdown with DVL2-overexpression groups. F The efficiency of ASPM203 knockdown and DVL2 overexpression were evaluated by western blotting. G Schematic representation of a model that RBM10 C761Y mutation induced oncogenic ASPM isoforms and regulated Wnt/β-catenin signaling in CCA
留言 (0)