
記住我
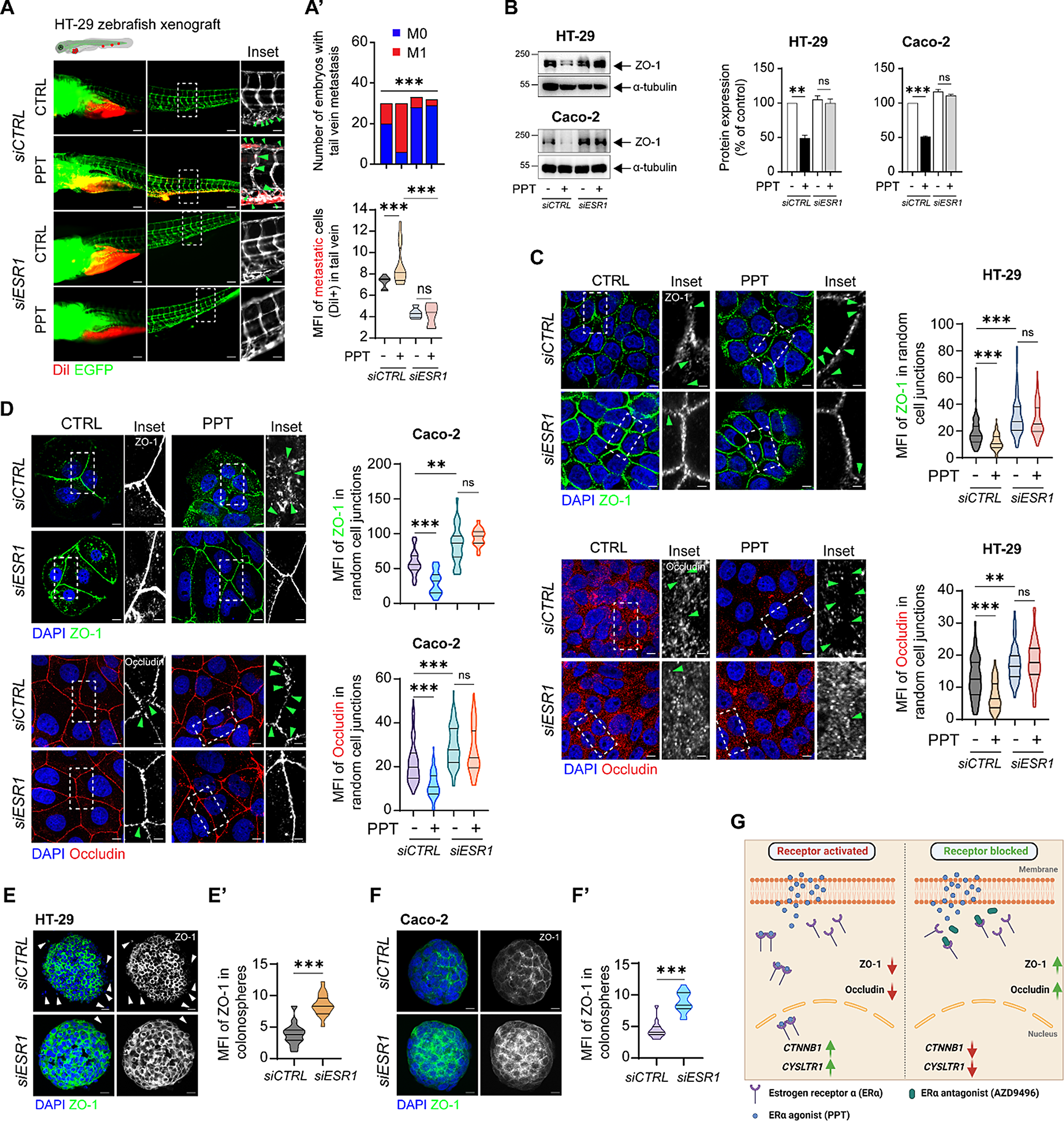



A better understanding of glyco-gene expression changes may facilitate the exploration of association between glycosylation and HBV-HCC. The differences in glyco-gene expression and glycosylation in HBV-HCC were investigated using an integrated strategy with a combination of transcriptomics, glycomics and glycoproteomics (Fig. 1A). First, GSE135631 and GSE94660 gene expression profiles, including 15 and 21 pairs of HBV-HCC and adjacent tissues respectively, were retrieved from the GEO database. A total of 7502 and 7839 genes were differentially expressed in GSE135631 and GSE94660, as shown in Fig. 1B, in which top 100 differentially expressed glyco-genes (DEGGs) were highlighted (Fig. 1B, Table S4). Partial Least Squares Discrimination Analysis (PLS-DA) of these DEGGs exhibited a clear separation of HBV-HCC from adjacent tissues (Fig. 1C). The expression patterns of all DEGGs across HBV-HCC and adjacent tissues from the combined two GSE study were shown as a heatmap (Fig. 1D). Next, functional enrichment analysis revealed that the main molecular function categories were carbohydrate binding, transferring glycosyl and hexosyl group activity (Fig. 1E). Protein–protein interaction (PPI) network of these DEGGs was constructed in an attempt to explicit the molecular mechanisms in the progression of HBV-HCC. Clustering analysis revealed ten major modules, in which protein glycosylation, GPI anchor biosynthetic process and glycosylation processes were mostly enriched (Fig. 1F; S1). Hub genes with a high degree of connectivity in the PPI network are significantly enriched in the process of fucosylation (FUT4, FUT6, FUT2 and FUT1), GPI anchor biosynthesis (PIGM, PIGV, PIGT and GPAA1), galactosylation (B4GALT3) and galNAcylation (B3GNT3).
Fig. 1
Differentially expressed glycan-related genes in HBV-related HCC of GSE135631 and GSE94660. A Workflow of the present study. B Volcano plot of expression patterns of identified genes in GSE135631 and GSE94660. Red dots: up-regulated genes. Green dots: down-regulated genes. Highlighted dots: DEGGs. The q value (log10) is plotted against the log10 (FC: HBV-HCC tissues vs. adjacent tissues) using the cut-offs of fold change > 1.5 or < 0.67 and p value < 0.05. C PLS-DA plot of DEGGs in GSE135631 and GSE94660. D Heatmap showing the expression pattern of DEGGs in HBV-HCC and adjacent tissues of GSE135631 and GSE94660. E Functional enrichment analysis of DEGGs. F The PPI network of DEGGs performed by the STRING database and cytoscape tools. The red colour intensity was proportional to the degree of connectivity. G The mRNA expression of 13 FUTs, PIGV, PIGT, PIGM and B4GALT3 genes in 3 pairs of HBV-HCC and adjacent tissues determined by RT-qPCR
Furthermore, FUTs family expression at mRNA levels were determined by RT-qPCR in HBV-HCC and adjacent tissues (Fig. 1G), revealed an elevated level of FUT1, FUT2, FUT4, FUT5, FUT8, FUT10, PIGT, PIGM and B4GALT3, as well as a reduced level of FUT3, FUT6, FUT7, FUT9, FUT11, FUT12 and PIGV in HBV-HCC tissues. Although HCC and adjacent tissue samples in transcriptomic analysis can not represent the whole HCC progression, these data provide clues in understanding dysregulated glycosylation especially fucosylation in HBV-HCC.
N-glycan profiles of normal, hepatitis, cirrhosis and HCC serum samplesIn our studies, N-glycans in HC, CHB, LC and HBV-HCC serum samples (Tab. S1) were profiled by MALDI-TOF/TOF–MS to reveal abnormal N-glycosylation. Representative MS spectra of N-glycans with signal-to-noise ratios > 5 were displayed and annotated (Fig. 2; Table S5). A total of 36 distinct m/z N-glycan structures were identified, with 31, 32, 25 and 25 N-glycans present in HC, CHB, LC and HBV-HCC samples, respectively. There were 21 N-glycans found in all groups but with different intensities.
Fig. 2
MALDI-TOF/TOF–MS spectra of N-glycans in HC, CHB, LC and HBV-HCC serum samples. Peaks of MALDI-TOF/TOF–MS spectra (signal-to-noise ratio > 5) were selected for relative intensity analysis in HC (A), CHB (B), LC (C) and HBV-HCC (D) samples. Detailed structures were annotated with GlycoWorkbench software. Proposed structures are indicated by m/z value
The expression pattern of identified N-glycans in HC, CHB, LC and HBV-HCC were exhibited in Fig. 3A. Hierarchical clustering revealed that LC/HBV-HCC were clearly separated from HC and CHB, however, LC/HBV-HCC could not be separated from each other. Furthermore, quantitative comparison analysis revealed that 25, 24, and 22 N-glycan structures were differentially expressed in CHB, LC and HBV-HCC versus HC. Of these N-glycans, 11 N-glycans were concertedly down-regulated, and 7 N-glycans were concertedly up-regulated in the progression of HBV-HCC. Notably, All up-regulated N-glycans were fucosylated (Fig. 3A; Table S5).
Fig. 3
N-glycan levels in HC, CHB, LC and HBV-HCC serum samples. A Expression pattern and dysregulation of N-glycans identified in HC, CHB, LC and HBV-HCC. B Relative abundances of high mannose, complex and hybrid N-glycans. The relative abundance is calculated by adding the relative abundances of a given type of N-glycan. C Relative abundances of bi-, mono- and afucosylated N-glycans. D Relative abundances of bi-, mono- and asialylated N-glycans. E Relative abundances of N-glycans with mono-, bi- and tri/tetra-antennary structures
N-glycans were classified into three types, including high mannose, complex and hybrid. Relative abundance of hybrid and complex N-glycans were increased in CHB, LC and HBV-HCC versus HC, while which of high mannose N-glycans were decreased (Fig. 3B). Consistent with elevated FUTs expression at mRNA levels, relative abundance of total fucosylation were up-regulated, of which mono-fucosylation levels were increased, while bi-fucosylation levels were decreased (Fig. 3C). Other terminal modification sialylation levels were increased, followed by a decrease in the progression of HBV-HCC (Fig. 3D). Significantly higher levels of bi-antennary structures were found in CHB, LC and HBV-HCC versus HC, while mono-antennary structures was progressively reduced (Fig. 3E). In combination of transcriptomic and glycomic analysis revealed that fucosylation levels were up-regulated in the progression of HBV-HCC.
Site-specific glycoproteomic profiling in HBV-HCC serum samplesTo decode the biological function of specific N-glycosylation especially fucosylation in the progression of HBV-HCC, intact glycoproteomic analysis of HC, CHB, LC and HBV-HCC serum samples (Table S2) were performed. A total of 1114 glycopeptides were identified (Fig. 4A; Table S6), of which, 1019, 1011, 999 and 982 glycopeptides, including 864 in common were found in HC, CHB, LC and HBV-HCC serum samples, respectively. The identified glycopeptides represented 129 glycosites were modified with 102 glycan structures. These N-glycans contained 8 high mannose, 13 hybrid, 77 complex and 4 paucimannose subtypes (Fig. 4B).
Fig. 4
Quantitative glycoprotemic analysis of HC, CHB, LC and HBV-HCC serum samples. A Venn diagram of intact glycopeptides identified from HC, CHB, LC and HBV-HCC samples. B Distribution of glycan subtypes from intact glycopeptides identified from HC, CHB, LC and HBV-HCC samples. C Volcano plots of expression patterns of identified glycopeptides. Red dots: up-regulated glycopeptides. Green dots: down-regulated glycopeptides. The value q (-log10) is plotted against the log10 (FC: disease group vs. normal group). D Classification of differentially expressed intact glycopeptides based on their attached glycan structures. The numbers indicate the unique glycopeptides modified by the corresponding glycans
Quantitative comparison analysis revealed that 116, 197 and 192 glycopeptides were differentially expressed in HC compared to CHB, LC and HCC, respectively, using the cutoff of fold change > 1.5 or < 0.67 and p value < 0.05 (Fig. 4C). These 288 differentially expressed glycopeptides (DEGPeps) from 73 glycoproteins were mainly decorated with complex type N-glycans, of which sialylated N-glycans accounted for the largest proportion, followed by fucosylated N-glycans and bi-antennary structures (Fig. 4D).
To understand the association between glycoprotein trajectories and the progression of HBV-HCC, hierarchical clustering for all DEGPeps was performed, with which four cluster (I-IV) were generated (Fig. 5A&B; Table S7). Glycopeptides in “cluster I” were continually increased in the progression of HBV-HCC, encompassing 27 proteins. These DEGPeps were significantly enriched in endopeptidase inhibitor activity, acute-phase response, blood microparticle and complement and coagulation cascades by functional enrichment analysis (Fig. 5C). Glycopeptides in “cluster II” were significantly decreased, followed by an increase (39 proteins), and mainly involved in the peptidase regulator activity, complement activation, blood microparticle and complement and coagulation cascades. Glycopeptides in “cluster III” were slightly decreased, followed by an increase (25 proteins), and enriched in regulation of humoral immune response, blood microparticle and complement and coagulation cascades. Glycopeptides in “cluster IV” (31 proteins) were decreased and primarily connected with peptidase regulator activity, acute-phase response, blood microparticle, complement and coagulation cascades. (Fig. S2).
Fig. 5
Hierarchical clustering analysis of differentially expressed glycopeptides identified in HC, CHB, LC and HBV-HCC. A Mfuzz clustering of differentially expressed glycopeptides. B Expression pattern of glycopeptides from cluster I, II, III and IV. C Functional enrichment analysis of differentially expressed glycopeptides from cluster I. D Heatmap of fucosylated N-glycans on intact glycopeptides from cluster I. PSMs of the intact glycopeptides, comprising of different glycans (bottom) and their glycosite locations in different glycoproteins (right) are exhibited in the heat map
Taking into account the escalating malignancy of HBV-HCC progression, 49 glycopeptides with fucosylation attached in “cluster I” were screened out, which were consisted of 23 fucosylated N-glycans and 25 peptides from 21 protiens (Fig. 5D). These fucosylated N-glycans contained three to seven HexNAc, three to eight hexose, up to four fucose, and two sialic acid. Among 21 identified glycoproteins, 16 contained 1 glycosite, 3 contained 2 glycosites, and 1 contained 3 glycosites. Notably, among these glycopeptides, IgA1-340-N5H5F1S2 was identified with highest PSM score.
Site-specific fucosylation on IgA1 and IgG2We further mapped N-glycan structures on each glycosite of DEGPeps from IgA1 and IgG2 based on our glycoproteomics data. A total of 30 and 5 unique intact glycopeptides were identified from IgA1 and IgG2, which were comprised 3 glycosites (144N#LT and 340N#VS for IgA1, 176N#ST for IgG2) and 31 glycans (Fig. 6A). Among these glycopeptides, fucosylation was accounted for 46% of all glycans on glycopeptides, and the majority of fucosylated intact glycopeptides were up-regulated in the progression of HBV-HCC.
Fig. 6
Site-specific glycan profiling of IGHA1 and IGHG2 identified in HC, CHB, LC and HBV-HCC. Heatmap showing all the identified glycans at the glycosite Asn-144, Asn-340 of IGHA1, and Asn-176 of IGHG2
Among these DEGPeps, 7 and 3 site-specific fucosylated glycopeptides from IgA1 and IgG2, showed a directionally concerted up-regulation in the HC-CHB-LC-HCC progression (Fig. 7A). To evaluate the diagnostic value of the aberrant fucosylated Igs in clinical practice, lectin-based ELISA assay with Concanavalin A (ConA), Lens Culinaris Agglutinin (LCA) and Aleuria Aurantia Lectin (AAL) was performed, which respectively recognize high mannose type N-glycans, α1,6-fucose, and α1,2/α1,3/α1,4/α1,6-fucose. The results revealed that total levels of IgG2 and IgA1 were not significantly changed, and different glycoform on Igs exhibited different expression patterns in the progression of HBV-HCC (Fig. 7B). AAL-reactive IgA1 and IgG2 were significantly elevated in HBV-HCC compared to HC, CHB and LC. LCA-reactive IgA1 was increased in HBV-HCC when compared to HC and decreased compared to CHB and LC, and LCA-reactive IgG2 was gradually increased and followed by a decrease in the progression of HBV-HCC. ConA-reactive IgA1 were increased in CHB, LC and HBV-HCC compared to HC, and ConA-reactive IgG2 showed lowest levels in HBV-HCC group. The different performance of these different glycoform on Igs indicated that glycosylation on Igs were specifically altered in the progression of HBV-HCC.
Fig. 7
Differentially expressed intact glycopeptides of IGHA1 and IGHG2. A Box plots of 10 site-specific fucosylated glycopeptides from IgA1 and IgG2 showed a directionally concerted up-regulation in the HC-CHB-LC-HCC progression. B Relative expression of IgA1/ IgG2 and ConA-/ LCA-/ AAL-reactive IgA1/ IgG2 in the HC, CHB, LC and HBV-HCC serum determined by ELISA
In summary, combined results of glycoproteomic and ELISA analysis revealed that IgA1 and IgG2 are highly fucosylated and AAL-reactive fucosylated IgA1 and IgG2 were up-regulated in HBV-HCC, implying their potential diagnostic value.
Proteomic analysis of HBV-HCC serum samplesTo comprehend the potential of the glycoproteins analysis and conclusions, proteomics of these samples have been performed, revealing 78, 163, and 186 differentially expressed proteins in HC compared to CHB, LC and HCC, respectively (Fig. 8A, Table S8). Among these differentially expressed proteins, 21 proteins, including AFP and diverse Igs, were up-regulated (p < 0.05) in patients with liver disease (CHB, LC, or HBV-HCC) compared to HC (Fig. 8B), that were mainly enriched in the cellular components of immunoglobulin complex, blood microparticle and cell periphery, the biological process of adaptive immune response, and molecular function of antigen binding (Fig. 8C). Notably, IgA1 did not show significant differences, while IgG2 was significantly up-regulated in patients with liver disease compared to HC (Fig. 8B). Peptides corresponding to the differentially expressed glycopeptides of Igs were investigated. The non-glycosylated peptides from IgA1 (amino acids 332–353; termed as IgA1[332–353]) did not show significant differences in the progression of HBV-HCC, while IgA1[127–153] and IgG2[172–180] were only detected in HBV-HCC (Fig. 8D), suggesting that fucosylated peptides from IgA1 and IgG2, especially IgA1[332–353], were specially up-regulated in the progression of HBV-HCC.
Fig. 8
Glycosylation site occupancy. A Volcano plots of expression patterns of identified proteins. Red dots: up-regulated proteins. Green dots: down-regulated proteins. The value q (-log10) is plotted against the log2 (FC: disease group vs. normal group). B Lists of concertedly up-regulated proteins in patients with liver disease (CHB, LC, or HBV-HCC) compared to HC. C GO enrichment analysis of the concertedly up-regulated proteins in HBV-HCC. D Levels of peptides from IgA1 and IgG2, including IgA1[127–153], IgA1[332–353] and IgG2[168–180] in HC, CHB, LC and HBV-HCC. E Average glycosylation site occupancy in HC, CHB, LC and HBV-HCC. Glycosylation site occupancy was calculated by dividing the abundance of a given type of glycoform by the total corresponding peptide (glycopeptide and non-glycopeptide) abundance. F Glycosylation site occupancy of IgA1[127–153], IgA1[332–353] and IgG2[168–180]. G Fucosylation site occupancy of IgA1[127–153], IgA1[332–353] and IgG2[168–180]
Furthermore, we correlated the abundance of glycopeptides with total corresponding peptides levels to determine the glycosylation site occupancy (Table S9). We found that the average site occupancy did not significantly changed (Fig. 8E). The glycosylation site occupancy of IgA1[127–153] and IgG2[172–180] exhibited no significant changes in the progression of HBV-HCC, and which of IgA1[332–353] was decreased in CHB compared to HC (Fig. 8F). Specifically, N-glycans on IgG2[172–180] were mainly fucosylated, and fucosylation site occupancy of IgA1[332–353] was elevated in LC and HCC compared to HC (Fig. 8G).
Collectively, When the IgA1 remains unchanged at protein levels, the fucosylation site occupancy of IgA1[332–353] increases, leading to a rise in IgA1[332–352]-N5H5F1S2 or -N6H3F1S2 and consequently resulting in elevated levels of AAL-reactive IgA1 in HBV-HCC. When fucosylation site occupancy of IgG2[168–180] remains unchanged, the IgG2 exhibited a upregulation at protein levels, contributing to a rise in IgG2[168–180]-N4H5F2 and consequently leading to a elevated level of LCA- and AAL-reactive IgG2 in HBV-HCC.
留言 (0)