Males of several insect species offer glandular secretions as nuptial gifts during mating (Gwynne, 2008, Lewis and South, 2012). These gifts are costly to produce (Voigt et al., 2005, Liu et al., 2015, Barbosa et al., 2018, McMahon et al., 2021). However, as a result of gift consumption, females can increase fecundity, and males can avoid sperm competition (Fedorka and Mousseau, 2002, Gwynne, 2008).
In many katydids and cricket species, the nuptial gift takes the form of a gelatinous mass; the spermatophylax, produced by the male’s accessory glands. The spermatophylax is attached to an ampulla, which contains the ejaculate and sperm. The spermatophylax and the ampulla together comprise the spermatophore. During copulation, the ampulla attaches to the female’s genitalia. Following copulation, the female consumes the spermatophylax while sperm migrates to her genital tract (Gwynne, 2001). The spermatophylax mainly consists of water, carbohydrates, proteins, and lipids (Heller et al., 1998, Heller et al., 2000, Marchini et al., 2009). As a result of its consumption, females reduce or lose their sexual receptivity and increase their lifespan and/or fecundity (Gwynne, 2008).
Access to the compounds contained in the spermatophore is limited by their availability in the environment and the ability of the males to gain access to them (Scharf, 2016). Therefore, allocating resources to the gifts may entail a lower apportionment of them to other functions (Lehmann & Lehmann, 2009), generating a trade-off between somatic and reproductive functions. Moreover, due to the constraints associated with resource allocation to the spermatophore, sexual and natural selection are expected to favor an optimal investment of the resources channeled by males in matings. Thus, males should strategically allocate ejaculates according to the perceived quality of the female. Males can increase their reproductive success by mating with virgin, young, and/or large females (Wedell, 1992, Simmons and Kvarnemo, 1997, Lehmann and Lehmann, 2009, Jarrige et al., 2013).
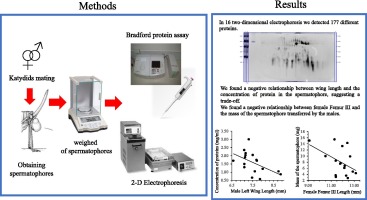
Despite their effect on male and female fitness, little is known of the compounds associated with katydid spermatophores (Heller et al., 1998, Simmons et al., 2013; Lehman et al., 2018), though these compounds may be similar to those found in other groups of insects (e.g., see reviews in Cordero, 1998, Gwynne, 2008). Our study explored the relationship between the mass of the spermatophore, concentration and the mass of proteins in the spermatophores produced by males of Conocephalus ictus (Orthoptera: Tettigoniidae) and the lengths of their femur III and forewing. Femur III was used to indicate general body size, whereas male forewing was considered an indicator of male investment in an energetically costly trait.
The production and maintenance of wings require considerable resources (see Harshman & Zera, 2007). Thus, flight can limit the assignation of resources to spermatophore production (Lewis & South, 2012). In the context of this study, we expected a negative relationship between male forewing and spermatophore parameters, that we did not expect in the Femur III. Also, since large individuals may invest more resources to spermatophore production (Wedell, 1993b, McCartney et al., 2008), we expected a positive relationship between male femur III length and the mass of spermatophore, the concentration and the mass of the proteins and its diversity. In addition to the relationship between male traits and spermatophore properties, we also explored the relationship between the spermatophore properties and the size of the females that received the nuptial gifts. Given that, in general, female body size is positively correlated with fecundity in insects (Honêk, 1993), we expected males to transfer more and diverse resources to large females due increase in potential offspring.
Conocephalus ictus is widely distributed throughout Mexico and Central America (Rehn, 1903, Naskrecki, 2000). In both males and females, there is a continual variation between winged and brachypterous individuals. The species is present all year in Los Tuxtlas, Veracruz, eastern Mexico. Females become adults after 75 d, whereas males become adults after 70 d (RCC personal observation). In the field, females are attracted phonetically. Mating occurs both during the day and at night, and males continue to sing during the process. When a female is receptive, she lays her abdomen on the abdomen of the male, who then transfers a spermatophore that adheres to the female’s genitalia. After mating, the female bends over her abdomen and consumes the spermatophore. Under laboratory conditions, oviposition began after both the spermatophylax and the ampulla were consumed. Consumption of the spermatophore increases the lifespan and fecundity of females, yet their receptivity turns off after mating (Ortíz-Jiménez & Cueva del Castillo, 2015).




留言 (0)