Membrane atg8ylation is a process of modification of diverse cellular membranes by ubiquitin-like mammalian ATG8 proteins (mATG8s) set in motion in response to membrane damage, stress, and remodeling signals (Fig. 1) [1]. This process is analogous to protein ubiquitylation, with, as stated elsewhere [1], “atg8ylation being to membranes what ubiquitylation is to proteins”. The principal components carrying out atg8ylation and ubiquitylation, mATG8s and ubiquitin, are closely related (Fig. 2), with mATG8s having additional features [2] that give them a preference for membranes as substrates. The conjugation cascades that activate mATG8s or ubiquitin and result in membrane atg8ylation or protein ubiquitylation are similar and involve activation with ATP and transfer of mATG8s or ubiquitin via E1, E2 and E3 ligases to their respective targets, membranes or ubiquitin [3]. The components of the conjugation cascade leading to membrane atg8ylation have additional properties allowing them to specialize for membranes 4, 5, 6, 7, 8, 9, 10. The result are two cascades with ATG12–ATG5 and mATG8–phosphatidylethanolamine (PE) conjugates as end products. The ATG12–ATG5 protein-protein conjugate associates with additional proteins to form E3 ligase complexes 8, 9, 10, 11 to marshal the formation of protein-lipid conjugates, culminating in atg8ylation of targeted membrane domains.
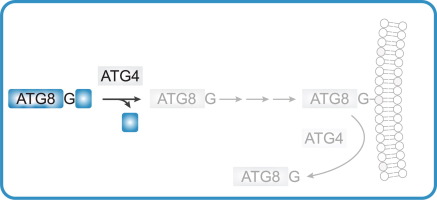
The atg8ylation cascade parallels ubiquitylation-like steps (Fig. 1A and B). There are seven principal mATG8s: LC3A, LC3B, LC3B2, LC3C, GABARAP, GABARAPL1 and GABARAPL2 12, 13, 14. The yeast’s single Atg8 has served an important role in delineating what can be viewed as a posttranslational modification with the lipid phosphatidylethanolamine (PE) 15, 16, which can also occur with another aminophospholipid, phosphatidylserine (PS) [17]. After proteolysis with ATG4s, the exposed C-terminial glycins of mATG8s are activated by ATP, and mATG8s transferred to ATG7 (E1) to form an mATG8-ATG7 conjugate, then transferred to ATG3 (E2) to form an mATG8-ATG3 conjugate, and finally covalently linked to PE in membranes guided by the E3 enzymes. Until very recently 8, 9, 10, the sole known E3 enzyme was the ATG12–ATG5 conjugate associated with ATG16L1 [11] before a new E3 ligase consisting of TECPR1 in a complex with ATG12–ATG5 conjugate has been characterized 8, 9, 10 (Fig. 1C), (Fig. 1C). The ATG12–ATG5 component of the atg8ylation E3 ligases is generated in its own conjugation cascade [11] (Fig. 1C). ATG12 is also a ubiquitin-like molecule. After activation by ATP It is conjugated to ATG7 (E1), then transferred to ATG10 (E2) and ultimately to ATG5. The resulting ATG12–ATG5 conjugate then noncovalently associates with ATG16L1 [11] or TECPR1 8, 9, 10, to form E3 ligases catalyzing atg8ylation of diverse membranes including autophagosomal and other membranous organelles [1]. ATG12–ATG5 containing E3 ligase activates its substrate mATG8-ATG3 by presenting the mATG8-G-C-ATG3 intermediate’s thioester bond for transfer of the mATG8 to the membrane via an amide bond with the PE headgroup [11].
The specialization of atg8ylation for membranes is supported by several mATG8s’ features and mechanisms. First, there are two additional (absent in ubiquitin) α-helices at the N-terminus of mATG8s with hidden affinities for membranes that become actualized during atg8ylation [2]. Second, the intrinsic membrane affinities of the atg8ylation cascade components ATG3 4, 5 and ATG16L1 6, 7, and as recently demonstrated for TECPR1 8, 9, 10, guide the atg8ylation system toward membranes. Third, as manifested during canonical autophagy, WIPI2, an effector of a stress-signaling phospholipid phosphatidylinositol-3-phosphate (PI3P) binds ATG16L1 18, 19, 20 and helps dock the ATG12–ATG5-ATG16L1 E3 ligase to the PI3P-marked membranes 7, 21. Alternatively, ATG16L1 can be guided to specific membranes by H+ V-ATPse [22]. In other cases, sphingomyelin presented on the cytofacial side of perturbed/damaged endolysosomal membranes recruits TECPR1 to guide atg8ylation of non-autophagic membranes 8, 9, 10.
The separation between membrane atg8ylation and protein ubiquitylation has exceptions, such as ubiquitylation of membranes and atg8ylation of proteins 23, 24, 25, 26, 27, 28. Thus, the specialization is not absolute, as often observed in many biological systems.
Diverse presentations of membrane atg8ylation guided by E3 ligases
In mammalian cells, membrane atg8ylation has been historically referred to as ‘LC3 lipidation’ and LC3 puncta formation 11, 29 and in the past considered as being synonymous with autophagy [30]. It has, however, become evident that ‘LC3 lipidation’ occurs on various endomembranes other than autophagosomes [31]. The membranes covalently modified by mATG8s, typically reported as LC3 lipidation include (Fig. 3): canonical autophagosomes [30], conventional phagosomes containing pathogens or microbial products 10, 22, 32, 33, 34, stressed or signaling endosomal compartments 33, 35, 36, 37, 38, lysosomes 9, 26, 39, 40, 41, 42, exocytic vesicles including exosomes 43, 44, ER during its ESCRT-dependent lysosomal degradation [45], and notably lipid droplets [46] which are delimited only by a phospholipid hemilayer.
The growing plethora of “non-canonical” phenomena (Fig. 3A,B) with no autophagic functions but engaging mATG8s (Fig. 3C) 1, 31 has led to a variety of terms and acronyms such as LAP (LC3-associated phagocytosis) [32], LANDO (LC3-associated endocytosis) [47], LC3-associated micropinocytosis (LAM) [36], SMAC (single membrane ATG8 conjugation) 48, 49 or CASM (conjugation of ATG8 to single membranes) 42, 50, and ‘V-ATPase-ATG16L1 axis xenophagy’ [22] that later on received the acronym VAIL (V-ATPase-ATG16L1-induced LC3 lipidation) 35, 37, 38.
There have been conceptual proposals to classify membrane atg8ylation based on single or double membranes as their substrates, reflected in many of the acronyms, as a main distinction between canonical autophagy and other non-canonical processes. Whereas autophagic organelles eventually mature into double membrane autophagosomes, prior to their closure they begin as single membranes (phagophores; Fig. 3B) and atg8ylation of these membranes takes place on the single membranes of the phagophores. Of note, the delimiting border of lipid droplets modified by LC3B is not even a conventional lipid bilayer but is a hemilayer of phospholipids surrounding cores made of neutral lipids (Fig. 3B) [46]. Thus, the membranes that are substrates for atg8ylation are not limited to double or single bileyers, and the minimal substrate is a phospholipid monolayer/hemilayer containing PE or PS.






留言 (0)